
Yousef Altaher
Faculty of Veterinary Medicine and Animal Resources, King Faisal University, Alhofuf, Alahsa, Saudi Arabia
Masayuki Nakanishi
Faculty of Pharmaceutical Sciences, Matsuyama University, Bunkyo-Cho, Ehime, Matsuyama, Japan
Mahmoud Kandeel
Department of Physiology, Biochemistry and Pharmacology, Faculty of Veterinary Medicine and Animal Resources, King Faisal University, Alhofuf, Alahsa, Saudi Arabia
International Journal of Pharmacology
Year: 2015 | Volume: 11 | Issue: 3 | Page No.: 243-247







ABSTRACT
While camels rapidly evoluted several genes to adapt their environment, contemporary studies recorded that camels are sensitive to toxicity with some drugs as monensin, narasin and maduramicin ionophores compounds, which are extensively used in veterinary medicine. Recently, camel genome was resolved, which will allow for discovery of secrets of camel life. To this end, it is not known whether camel cytochrome enzymes were specially developed or adapted to camel environment or it has been associated with increasing the susceptibility of camels to toxicity with drugs. The objectives of this work are annotation of camel genome and retrieval of camel for cytochrome P450 1A2 (CYP 1A2) for phylogenetic, molecular modeling studies, docking studies and determination of its drug binding power. Results indicated poor efficiency of camel CYP 1A2 in binding with drugs. Furthermore, despite the high evolution of many camel genes to adapt camel environment, CYP 1A2 was found to be phylogenetically retarded gene showing lower rate of evolution compared with subset of data from 100 different organisms. These results suggests the potential toxicity of camels with drugs undergoing oxidation by CYP 1A2.
PDF Abstract XML References Citation
Received: October 06, 2014;
Accepted: January 13, 2015;
Published: March 04, 2015
How to cite this article
Yousef Altaher, Masayuki Nakanishi and Mahmoud Kandeel, 2015. Annotation of Camel Genome for Estimation of Drug Binding Power, Evolution and Adaption of Cytochrome P450 1a2. International Journal of Pharmacology, 11: 243-247.
DOI: 10.3923/ijp.2015.243.247
URL: https://scialert.net/abstract/?doi=ijp.2015.243.247
DOI: 10.3923/ijp.2015.243.247
URL: https://scialert.net/abstract/?doi=ijp.2015.243.247
INTRODUCTION
Camels are raised in arid and semiarid environment. For this reason, camels have especially evoluted some genes for accustoming its surrounding environment. For example, camels have unique water and heat regulation metabolic pathways that allowed them to live in the desert environment. This adaptation is accompanied by rapid evolution of water and regulation mechanisms (Wu et al., 2014). As a form of adaptation, the camel mitochondrial proteins showed higher evolution rate in camelids to allow for living in different environments (Di Rocco et al., 2006, 2009).
Camels are sensitive to toxicity with some drugs which have wide safety margin in other animals (Ali, 1988; Mousa and Elsheikh, 1992; El Bahri et al., 1999; Alquarawi and Ali, 2000). For instance, ionophores compounds as monensin, narasin and maduramicin are extensively applied in veterinary medicine for their antimicrobial, antiprotozoal and growth promoting effect (Miller et al., 1990; Mousa and Elsheikh, 1992; Abu Damir et al., 2013). Camels are very sensitive to toxicity with ionophores compared with other animals as poultry, sheep and cattle. A camel can die after administration of low dose of 0.6 mg kg-1. In contrast, poultry can withstand at least 30 folds higher of such dose. Similarly, diminazine aceturate is a common drug for treating blood parasites in animals is not recommended for camels due to serious toxicity, which is well tolerated in other species (Homeida et al., 1981).
The camel genome now is already available for public use (Wu et al., 2014). This study is directed toward the use of the recently announced camel genome sequences for discovering the genetic bases of camel susceptibility to ionophores toxicity. Furthermore, the rate of evolution of camel cytochrome P450 A2 will be investigated and compared with other species. Cytochrome P450 1A2 is a major enzyme in drug and xenobiotics metabolism. In order, to compare the drug binding power with other well characterized enzymes e.g., human CYP 1A2, docking studies will be conducted to determine the binding affinity for monensin, narasin and maduramicin. In this study, we show the slower evolution and lower drug binding capacity and unstable complexes with narasin and maduramicin of camel CYP 1A compared with the human enzymes were showed. This might suggest the reason beyond camel toxicity with these compounds.
MATERIALS AND METHODS
Collection of camel genome data: The Arabian camel genome database at King AbdulAziz City of Science and Technology (http://www.camel.kacst.edu.sa “Last Time Access on This Date 2015-01-06”) and the genome databases at the National Institute of Heath were used to retrieve the genes and proteins of camel. The data from KACST included a set of expression sequence tags (EST library).
Handling of camel genes and retrieval of camel CYP A1 sequence: Human CYP1A2 sequence was used to BLAST the retrieved camel sequences. The highest contig hits were retrieved. The retrieved contig was then used to BLAST gene databases at NCBI to get the highest similarity hits. Handling of genomic data, BLAST search and sequence alignment were performed by CLC genomics workbench.
Analysis of gene evolution rate: The evolution rate was assessed by BEAST programs (Drummond and Rambaut, 2007; Drummond et al., 2012). Geneious 7.1.7 software package was used to build alignment in NEXUS format. The BEAUti software was used to convert NEXUS file to xml format for Bayesian evolutionary analysis. Substitution rate was set for 1, gamma category count was set to 4 and the JC69 was used as a substitution model. Timing data were not provided and the branch lengths represented the substitutions per year with an assumed average of 1. Relaxed clock log normal was set for clock model at clock rate of 1 and uniform birth rate. Marcov Chain Monte Carlo (MCMC) mathematical model is used by BEAST, the length of chain was set to 1000000 and the log file is saved for tree annotation by TreeAnnotator software. The produced tree is visualized and annotated by FigTree software. Examination of MCMC results were viewed Tracer program.
Analysis of binding strength of camel CYP1 with drugs: A structure model of camel CYP1A2 was built by using SWISS model web server. The obtained structure model was checked with verify 3D program and exported.
 | |
| Fig. 1: | Structure of the compounds used in this study |
The structure of monensin, narasin and maduramicin were obtained from pubchem database (Fig. 1). Docking studies were done by Molegro virtual docker. Comparative drug binding was with human CYP1 PDB ID 2HI4.
RESULTS
Evolution rate of camel CYP evolution: After BLAST search of gene databases with camel CYP the top 100 high similarity sequences and built phylogenetic tree were retrieved (Fig. 2). The average gene evolution rate among all organisms was 0.94. human CYP showed more acceleration by showing 1.18 evolution rate (Table 1). In contrast camel CYP showed slower rate compared with human and also was lower than the mean rate in the tested organisms.
| Table 1: | Parameters of rate obtained by BEAST analysis and Tracer program for CYP1A |
The binding of monensin, narasin and maduramicin with human and camel CYP1A2: The comparison of monensin, narasin and maduramicin binding properties of camel and human CYP1 are shown in Table 2 and Fig. 3. The molecular model retrieved from SWISS model server is represented in Fig. 4. During setting up of docking run, docking was set to accurate or slow mode to get the maximum efficiency of results. Ten poses for every drug were retrieved and examined in the their active binding site for any stearic problems. The ligand efficiency L1 and L2 as a parameter for comparison was used.
| Table 2: | Results of docking studies* |
 | |
| *L stands for ligand efficiency | |
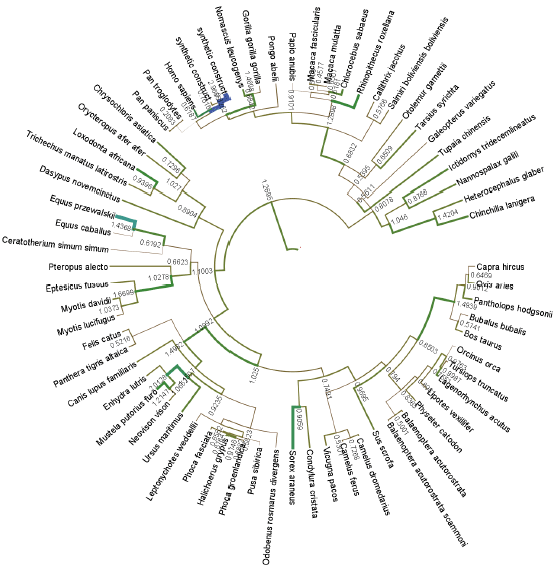 | |
| Fig. 2: | Circular cladogram showing evolution pattern of CYP1A2s |
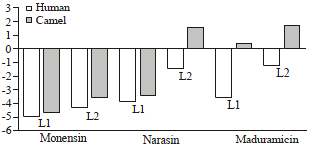 | |
| Fig. 3: | Comparison of human and camel docking parameters |
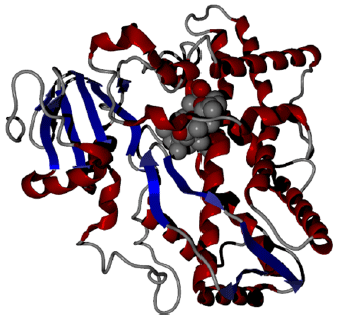 | |
| Fig. 4: | Camel CYP1A2 model built by SWISS Model server obtained my Molegro Virtual Docker |
L1 is the docking score divided by the number of atoms in molecule. L2 is a rerank for L1 after consideration of stearic forces. Therefore, L1 and L2 parameters are more accurate for comparing two different drugs across two different molecular targets. Furthermore, L2 is more valuable in comparison as it is a reranking algorithm for L1.
The data show that all compounds are dockable to the active site of human and camel CYP1A2 (due to the negative values of L1 and L2) except narasin and maduramicin, which is expected to be poor substrate to camel CYP1 due to positive value of L2. The data also indicates weaker binding to the camel enzymes compared with human CYP1A. This evidenced by lower negative values of L1 and L1.
DISCUSSION
Camels were domesticated to be reared for human usage about 3000-6000 years ago (Burger and Palmieri, 2014). Investigation of gene evolution by molecular modeling studies lead to understanding the physiology, disease development and treatment opportunities (Kandeel and Kitade, 2013a, b; Kandeel et al., 2014). This study showed lower evolution rate of camel CYP. Furthermore, docking studies showed lower binding potency of camel CYP with two toxic compounds with camels, monensin, narasin and maduramicin. The human CYP was more effective in binding to these compounds.
Narasin and maduramicin are expected to form unstable complexes with camel enzyme due to the positive value of L2. Thus, the camel enzyme binds monensin with lower potency than human and expected to do not bind with narasin and maduramicin.
Although camels rapidly evoluted some genes to help it to adapt the harsh environment, the rate of CYP evolution was lower than other animals. It was may interpreted that this is a form of adaptation. The stress of xenobiotics and toxicity might be lower in desert environment, that does not put metabolizing enzymes under selection pressure. In addition, in the advent of new development of drugs for veterinary and other uses, together with modernization of camel habitat would put camels to be susceptible to toxicity more than other species. This is due to poor development of their CYP enzymes.
CONCLUSION
Camel genomics data were used to conclude the genetic bases of camel susceptibility to ionophores anticoccidials. Camels showed lower evolution rate of cytochrome P450 1A2 enzyme with lower and unstable binding with drugs. Therefore, care should be taken in using drugs undergoing metabolism by oxidation in camels due to their potential toxicity.
ACKNOWLEDGMENTS
This work is supported by grants for projects of undergraduate students to MK and YA from the deanship of scientific research, King Faisal University, Hofuf, Alahsa, Saudi Arabia.
REFERENCES
- Abu Damir, H., M.A. Ali, T. Abbas, E. Omer and A. Al Fihail, 2013. Narasin poisoning in the dromedary camel (Camelus dromedarius). Comp. Clin. Pathol., 22: 305-311.
CrossRefDirect Link - Ali, B.H., 1988. A survey of some drugs commonly used in the camel. Vet. Res. Commun., 12: 67-75.
CrossRefDirect Link - Alquarawi, A.A. and B.H. Ali, 2000. A survey of the literature (1995-1999) on the kinetics of drugs in camels (Camelus dromedarius). Vet. Res. Commun., 24: 245-260.
CrossRefPubMedDirect Link - Burger, P.A. and N. Palmieri, 2014. Estimating the population mutation rate from a de novo assembled bactrian camel genome and cross-species comparison with dromedary ESTs. J. Hered., 105: 839-846.
CrossRefDirect Link - Di Rocco, F., G. Parisi, A. Zambelli and L. Vida-Rioja, 2006. Rapid evolution of cytochrome c oxidase subunit ii in camelids (Tylopoda, Camelidae). J. Bioenerg. Biomembr., 38: 293-297.
CrossRefPubMedDirect Link - Di Rocco, F., A.D. Zambelli and L.B. Vidal Rioja, 2009. Identification of camelid specific residues in mitochondrial ATP synthase subunits. J. Bioenerg. Biomembr., 41: 223-228.
CrossRefDirect Link - Drummond, A.J. and A. Rambaut, 2007. Beast: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol., Vol. 7.
CrossRefDirect Link - Drummond, A.J., M.A. Suchard, D. Xie and A. Rambaut, 2012. Bayesian phylogenetics with beauti and the beast 1.7. Mol. Biol. Evol., 29: 1969-1973.
CrossRefDirect Link - El Bahri, L., O. Souilem, M. Djegham and J. Belguith, 1999. Toxicity and adverse reactions to some drugs in dromedary (Camelus dromedarius). Vet. Hum. Toxicol., 41: 35-38.
PubMed - Homeida, A.M., E.A. El Amin, S.E. Adam and M.M. Mahmoud, 1981. Toxicity of diminazene aceturate (Berenil) to camels. J. Comp. Pathol., 91: 355-360.
CrossRefPubMedDirect Link - Kandeel, M., A. Al-Taher, R. Nakashima, T. Sakaguchi and A. Kandeel et al., 2014. Bioenergetics and gene silencing approaches for unraveling nucleotide recognition by the human EIF2C2/Ago2 PAZ domain. PloS ONE, Vol. 9.
CrossRefDirect Link - Kandeel, M. and Y. Kitade, 2013. Computational analysis of siRNA recognition by the Ago2 PAZ domain and identification of the determinants of RNA-induced gene silencing. PLoS ONE, Vol. 8.
CrossRefDirect Link - Kandeel, M. and Y. Kitade, 2013. In silico molecular docking analysis of the human argonaute 2 paz domain reveals insights into rna interference. J. Comput.-Aided Mol. Des., 27: 605-614.
CrossRefDirect Link - Miller, R.E., W.J. Boever, R.E. Junge, L.P. Thornburg and M.F. Raisbeck, 1990. Acute monensin toxicosis in stone sheep (Ovis dalli stonei), blesbok (Damaliscus dorcus phillipsi) and a bactrian camel (Camelus bactrianus). J. Am. Vet. Med. Assoc., 196: 131-134.
PubMed - Mousa, H.M. and H.A. Elsheikh, 1992. Monensin poisoning in dromedary camels. Deutsche Tierarztlilche Wochenschrift, 99: 464-464.
PubMed - Wu, H., X. Guang, M.B. Al-Fageeh, J. Cao and S. Pan et al., 2014. Camelid genomes reveal evolution and adaptation to desert environments. Nat. Commun., Vol. 5.
CrossRefDirect Link