
Juan Yan
Department of Pharmacology, Hebei Medical University, Shijiazhuang, People`s Republic of China
Maodong Zheng
Department of Pharmacy, The First Affiliated Hospital of Hebei North University, Zhang Jiakou, People`s Republic of China
Danshen Zhang
Hebei University of Science and Technology, Shijiazhuang, People`s Republic of China
International Journal of Pharmacology
Year: 2014 | Volume: 10 | Issue: 1 | Page No.: 55-68







ABSTRACT
Previous studies have indicated that Chrysophanol Liposomes (Chr-lip) have demonstrated potent neuroprotective effects against Cerebral Ischemia Reperfusion (CIR) injury but supporting evidence at molecular mechanisms is lacking. Two hundred healths Kunming male mice were randomly divided into the sham group, model group and Chr-lip treatment (10.0, 5.0 and 0.5 mg kg-1) groups. Chr-lip treatment groups intraperitoneally injected with Chr-lip for three successive days, then subjected to brain ischemia induced by MCAO. After reperfusion 24 h, neurological deficits, brain water content, neuronal ultrastructure, histopathological changes, oxidative stress-related biochemical parameters, neuronal apoptosis and apoptosis-related proteins were assessed by the methods of HE stainning, Hoechst33258 stainning, kits, western blot and real-time quantitative PCR. Chr-lip significantly improved neurological deficits (p<0.05), neuronal ultrastructure and histological deficits, reduced brain water content (p<0.05) and decreased apoptotic cells (p<0.05) after cerebral ischemia. Chr-lip also markedly reduced the level of malondialdehyde (MDA) and enhanced the activities of antioxidant enzymes superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) in CIR mice (p<0.05). Further investigation demonstrated that Chr-lip remarkably promoted the expression of Bcl-2 and inhibited the expression of Bax, Cytochrome C (Cyt-C) and Caspase3 at protein and mRNA levels (p<0.05), respectively. These findings indicate that Chr-lip has protective effects against CIR injury in mice and the neuroprotective effects may be attributed to attenuating oxidative stress and inhibiting neuronal apoptosis.
PDF Abstract XML References Citation
Received: December 07, 2013;
Accepted: March 17, 2014;
Published: May 13, 2014
How to cite this article
Juan Yan, Maodong Zheng and Danshen Zhang, 2014. Chrysophanol Liposome Preconditioning Protects against Cerebral Ischemia-reperfusion
Injury by Inhibiting Oxidative Stress and Apoptosis in Mice. International Journal of Pharmacology, 10: 55-68.
DOI: 10.3923/ijp.2014.55.68
URL: https://scialert.net/abstract/?doi=ijp.2014.55.68
DOI: 10.3923/ijp.2014.55.68
URL: https://scialert.net/abstract/?doi=ijp.2014.55.68
INTRODUCTION
Cerebral ischemia is one of the leading causes of disability and human death across the world and its incidence is speculated to rise with the increase in the number of the aging population (Wang et al., 2012a). There are several pathological mechanisms in the process of cerebral ischemic such as excessive release of excitatory amino acids (Wang et al., 2013a), loss of ionic homeostasis (Shah and Abbruscato, 2013), energy failure (Ten and Starkov, 2012), inflammatory response (Liu et al., 2014), increased oxidative stress (Yan et al., 2014) and apoptosis (Guan et al., 2013). These mechanisms eventually cause irreversible damage of the brain tissue. So recent years, chemical drugs such as calcium ion antagonist and radical scavengers as well as neuroprotective agents have been used for the treatment of CIR injury. However, side effects such as resistance to drugs, cerebral hemorrhage and gastrointestinal irritation may exceed the clinical benefits for long-term therapy (Chen et al., 2014). Furthermore, chemical drugs are generally difficult to achieve significant therapeutic results. Fortunately, clinical applications and experimental reports of traditional Chinese medicines against CIR injury have been ascendant (Qi et al., 2009; Zhao et al., 2010; He et al., 2012).
Chrysophanol (Chry), an important anthraquinone compound, come from the dry roots of Rheum palmatum. Whose chemical structure is made of anthraquinone rings (Fig. 1) and it can scavenge O2¯ and diphenylpicrylhydrazyl (DPPH) free radicals (Iizuka et al., 2004). Numerous studies have demonstrated that Chry possesses anti-allergic (Kim et al., 2000), antifungal (Agarwal et al., 2000), anti-tumor (Shi et al., 2001) and anti-apoptotic properties (Chen et al., 2002). Our previous studies demonstrated that Chr-lip had the obvious bioavailability and could improve passive avoidance learning and memory after CIR in mice than Chry monomer (Li and Zhang, 2011).
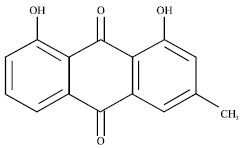 | |
| Fig. 1: | Chemical structure of chrysophanol |
In addition, Chr-lip has the character of brain-targeting. However, the protective effect of Chr-lip on CIR and its mechanisms have not been well elucidated. Oxidative stress has been considered as a key harmful factor in CIR injury and may ultimately result in the initiation of pathways that lead to apoptotic cell death (Manzanero et al., 2013). Apoptosis is one of the major pathways that lead to cell death after CIR injury (Broughton et al., 2009). Considering the facts mentioned above and the role of oxidative stress and apoptosis in the pathophysiology of CIR injury, the present study was designed to evaluate the neuroprotective effects of Chr-lip, as well as the underlying mechanisms by focusing on oxidative stress and neuronal apoptosis.
MATERIALS AND METHODS
Chemicals and reagents: Rheum palmatum was obtained from Zhangheng traditional Chinese medicine CO. (Zhangjiakou, China, Batch No. 120401), were identified by Prof. Shu-lan Ma, College of Pharmacy, Hebei North University, Zhangjiakou, China. Chrysophanol obtained from the dried roots was used in this study. The reagent kits of malondialdehyde (MDA), glutathione peroxidase (GSH-Px) and superoxide dismutase (SOD), Hoechst33258 were the products of Nanjing Jiancheng Bioengineering Institute (Nanjing, China). HPLC grade methanol was obtained from Beckman Coulter, Inc (Brea, CA, USA). We purchased primary antibodies against β-actin, Bax, Bcl-2, Cyt-C and Caspase3 (Beijing Bioss co., LTD Beijing, China). The reagent kits of first strand TIANScript-cDNA, SuperReal PreMix (SYBR-Green) were the products of Tiangen Biotech Co., LTD (Beijing, China). Primers of Bax, Bcl-2, Cyt-C, Caspase3 and GAPDH were designed by Beijing Dingguo Changsheng Biotechnology Co., LTD (Beijing, China). All other chemicals and reagents were of analytical grade.
Animals: Adult male Kunming mice, 28-30 g, were housed in a room with temperature of 22°C, relative humidity of 55% and a 12 h light/12 h dark cycle. All animal experiments and care were performed in accordance with the procedure approved by the Animal Ethics Committee of the Hebei North University (Chinese License Key Number: SCXK (Ji) 2004-0001). This study was approved by the Local Ethics Committee in May 2012.
Separation and purification of chrysophanol by PHPLC: Degreased anthraquinone glycosides were hydrolyzed with dilute sulfuric acid in rhubarb. We extracted aglycone by hot chloroform. Impurity was removed from extracting solution accordingly using different alkaline solution such as rheinic acid and archen. Chry was precipitated through sodium hydroxide with hydrochloric acid. The precipitate was dried in vacuo. PHPLC (PHPLC, Agilent, USA and California) method was established for purifying the substances of Chry. The column was ZORBAX SB-C18: (21.2x250 mm, 7 μm) with a mixture of methanol-0.01% phosphoric acid (85:15) solution as the mobile phase, at a flow rate of 20 mL min-1. The column temperature was at 28°C. The detection wavelength was 254 nm. Fractions were collected based on the peak, the threshold being Min: 2.2. Both Nuclear Magnetic Resonance (NMR) and HPLC were used for the structure and quantitative analysis of Chry. HPLC (Agilent, USA, California) method was established for quantification of Chry. The column was Hypersil ODS2 (4.6x150 mm, 5.0 μm), the column temperature was at 35°C, the injection volume was 20 μL, the mobile phase was methanol-0.1% phosphoric acid (85:15), the flow rate was 1 mL min-1, the detection wavelength was 254 nm.
Preparations of Chr-lip: According to the previous study (Wang et al., 2011) the optimized preparation conditions were as follows: lecithin (30.0 mg), cholesterol (10.0 mg) and Chry (3.0 mg), vitamin E (10.0 mg), PEG2000, dehydrated alcohol (10 mL), Tris damping fluid (10 mL), hydration time was 50 min and hydration temperature was 45°C. The organic solvent was completely evaporated and shaken for 2 h at 40°C. Finally, we obtained high, middle and low-doses of Chr-lip (10.0, 5.0, 0.5 mg kg-1). Chr-lip permeability was 2.8 stored for 15 days at 4°C.
Surgical procedure: Mice (n = 40 per group) were anesthetized by ip 3.5% chloral hydrate (10 mL kg-1). Body temperature was regulated at 37°C by homoisothermy bench. The model was similar to that described previously (Wang et al., 2012b). After the skin incision, the External Carotid Artery (ECA), the left Common Carotid Artery (CCA) and the Internal Carotid Artery (ICA) were carefully exposed and dissected away from adjacent nerves. Microvascular aneurysm clips were applied to the ICA and the left CCA. A coated filament was introduced into an arteriotomy hole, fed distally into the ICA and advanced 10 mm from the carotid bifurcation. The ICA clamp was removed and focal cerebral ischemia started. After ischemia for 2 h, the filament was gently removed. The collar suture at the base of the ECA stump was tightened. The skin was closed, anesthesia discontinued and the animals were returned to the pre-warmed cages. Sham group underwent the same surgery but not subjected to ischemia-reperfusion.
Experimental groups: Following successful surgery the mice were divided into model group, high, middle and low-dose treated group. In the randomized and controlled animal experiment, mice were divided into the sham, model and Chr-lip treatment (10.0, 5.0, 0.5 mg kg-1) groups. Chr-lip was i.p. once a day for three days before ischemia and once at 2 h before the onset of ischemia. While the sham and model groups were given the same volume of saline (mL 200 g-1). Animals were sacrificed and tissues were removed for analysis after 24 h of reperfusion.
Measurement of neurological deficits: Neurological deficits of each mouse were determined after 24 h of reperfusion according to Clark et al. (1997) by a single experimenter who was blinded to the experimental group.
Assessment of the brain water content: Mice were anesthetized by an intraperitoneal injection of 3.5% chloral hydrate (10 mL kg-1) and decapitated in 24 h after reperfusion. The brains were taken and then separated into left and right hemisphere. According to the previous study (Li et al., 2009a), the brain samples were weighed immediately in order to obtain wet weight. And then, we put those brain samples in an oven at 100°C for 48 h to obtain the dry weight. The brain water content was calculated through:
Observations of cerebral neuronal ultrastructure: After heart reperfusion, brain tissues (1 mm3) were cut and fixed in 2.5% glutaraldehyde in PBS at 4°C for 24 h. The tissues were then dehydrated, permeated, embedded, stained and cut into ultra-thin sections. Ultrastructual observations of the neuronal tissues were made under a transmission electron microscope (H-7650, Hitachi, Tokyo, Japan).
Measurement of histopathological changes: After 24 h of reperfusion, mice were anesthetized and perfused with NS and 4% paraformaldehyde in 0.1 M phosphate buffer solution (PBS, pH 7.4). Brains were taken and further fixed in 4% paraformaldehyde at 4°C for 24 h and then cut into equally spaced blocks. Paraffin-embedded blocks were cut into a series of 5 μm thick slices and stained with Hematoxylin Eosin (HE) and examined under a light microscope (Olympus, Tokyo, Janpan; type CX21, x40).
Determination of oxidative stress indicators: The ischemic hemispheres (n = 6, for each group) were homogenized in ice-cold saline (1: 9). According to the previous study (Wang et al., 2013b), the homogenate was centrifuged at 3,000 g and 4°C for 15 min. Afterwards the supernatant was used to determine the content of MDA, activities of SOD and GSH-Px in accordance with the manufacturer's instructions (Jiancheng Institute of Bioengineering, Nanjing, China).
Hoechst33258 staining: Mice were anesthetized by an intraperitoneal injection of 3.5% chloral hydrate and then quickly perfused with normal saline and 4% paraformaldehyde in PBS. The brain was removed, fixed in 4% paraformaldehyde for 24 h, immersed overnight in 70% ethanol, dehydrated and paraffin embedded. After sectioning, tissues were deparaffinized with xylene two times for 20 min each time, dehydrated with an ethanol gradient and then rinsed with PBS. The sections were incubated with 25 mM Hoechst 33258 (Beyotime Institute of Biotechnology, Beijing, China) for 15 min at 37°C, washed with PBS, mounted onto slides using antifade mounting medium and then examined under a 90i multifunction microscope (Nikon, Tokyo, Japan).
Western blot analysis: Mice were anesthetized by an intraperitoneal injection of 3.5% chloral hydrate and then quickly perfused with normal saline. Brains were immediately removed and stored at -80°C until use for detecting protein expression of Bax, Bcl-2, Cyt-C and Caspase3. Protein concentrations were determined using a BCA kit. Samples loaded at equivalent amounts of total protein (25 μg) were analyzed by 12% SDS-PAGE. After separation, proteins were subsequently transferred to a polyvinylidene difluoride (PVDF) membrane by a semi-dry transfer method which was then incubated with 5% skim milk for 1 h at 37°C. Afterward, the PVDF membrane was incubated with the following primary antibodies: Rabbit polyclonal anti-Bax (1:200), rabbit polyclonal anti-Bcl-2 (1:200), rabbit polyclonal anti-Cyt-C (1:200), rabbit polyclonal anti-caspase3 (1:200) or rabbit polyclonal anti-β-actin (1:200) antibody (Beijing Biosynthesis Biotechnology, Beijing, China) overnight at 4°C. After washing with TBST, the PVDF membrane was incubated with the appropriate secondary antibody (goat anti-rabbit IgG/HRP 1:1500) (Beijing Biosynthesis Biotechnology) for 1 h at 37°C and developed using a DAB kit. The images were quantified using Quantity One software and β-actin served as the internal standard.
qRT-PCR analysis: Mice were anesthezed by an intraperitoneal injection of 3.5% chloral hydrate and then quickly perfused with normal saline. Brains were immediately removed and frozen in liquid nitrogen and stored at -80°C. Total RNA was extracted using Trizol (Tiangen Biotech Beijing Co., Beijing, China) and reverse transcribed into cDNA using the TIANScript RT Kit (Tiangen Biotech Beijing Co., Beijing, China) according to the manufacturer’s instructions. The newly synthesized cDNA was amplified using the SuperReal PreMix (SYBR Green) PCR kit (Tiangen Biotech Beijing Co., Beijing, China) and detected with the 7300 real time PCR System (ABI, Carlsbad, CA, USA). Primer sequences and PCR product lengths for various transcripts were as follows: Bax, forward 5’-TTTTTGCTACAGGGTTTCATCCAGG-3’, reverse 5’-CCATATTGCTGTCCAGTTCATCTCC-3’, 151 bp; Bcl-2, forward 5’-ACCCTCCTGATTTTTCCTCCACC TA-3’, reverse 5’-AATACATAAGGCAACCACACCAT CG-3’), 119 bp; Cyt-C (forward 5’-ATAGAACCAAGAA GGA GATTGACCA-3’, reverse 5’-TACAGACACCTA TCAGAATAACCCA-3’), 120 bp; Caspase3 (forward 5’-GACTTCTCTGACTATCGTCGTGCTG-3’, reverse 5’-CGCATAAGCAACACCCACAGTAGTA-3’), 126 bp; GAPDH forward 5’-ATGACATCAAGAAGGTGGTGAAG CA-3’, reverse 5’-GAAGAGTGGGAGTTGCTGTTGAAG T-3’), 112 bp. Relative quantification of transcript abundance was carried out using the following equation:
F = 2-[Ct(target, test)-Ct(ref, test)]- [Ct(target, calibrator)-Ct(ref, calibrator)]
Statistical analysis: Statistical analyses were performed using SPSS 10.0 (SPSS, Chicoga, IL, USA). Statistical differences between groups were analyzed using one-way analysis of variance (ANOVA) and Least Significant Differences (LSD) test. Results are expressed as Means±SD. The level of significance was p<0.05.
RESULTS
Purity of Chry and entrapment efficiency in Chr-lip: The chemical formula of the product was determined to be C15H10O4. 1H-NMR (DMSO-d6): δ: 12.17(1H, s, -OH), 12.07(1H, s, -OH), 7.15(1H, s, C2-H), 7.70 (1H, s, C4-H), 7.87 (1H, d, C5-H), 7.72(1H, dd, C6-H), 7.32 (1H, dd, C7-H), 2.51 (3H, s, Ar-CH3). The purity of Chry was 98.9%; the entrapment efficiency of Chr-lip was 88.5% (Fig. 2).
Neurological scores and the water content of brain: We found that the neurological scores in sham group were (1.17±0.76) and the neurological scores in model group were increased to (27.33±1.53). Pretreatment with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) significantly reduced the neurological deficits (p<0.01, p<0.01, p<0.05) (Fig. 3a).
As shown in Fig. 3b, was significantly increased in model group compared with sham group (75.05±3.121% vs. 58.67±4.04%, p<0.01). Pretreatment with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) significantly reduced the branin water content (p<0.01, p<0.01, p<0.05).
Effects of chr-lip on neuronal ultrastructural changes: As shown in Fig. 4, in the sham group, chromosomes were distributed uniformly and the nuclear membrane was clearly seen. The structures of the abundant mitochondria and the Golgi apparatus were distinct. In the model group, the cell nucleus was swollen and the nuclear membrane was disrupted or disintegrated. The chromatin was marginated and vacuoles were observed in the cytoplasm. Mitochondrial cristae and membranes had mostly disappeared while degranulation of the rough endoplasmic reticulum and apoptotic bodies were apparent. In the treatment groups, the chromosome distribution was relatively uniform and the nuclear membrane was clear. The mitochondria were reduced to some extent, but swelling of the rough endoplasmic reticulum was mild.
Effect on histopathology in mice with MCAO-induced CIR injury: As seen in Fig. 5a, the sham group showed normal neurons with no pathological change. On the other hand, in the model group, neuronal loss was severe and dying neurons showed shrunken cytoplasm and pyknotic nuclei and cellular edema (Fig. 3b). Chr-lip markedly reduced these ischemia/reperfusion induced histopathological changes (Fig. 5c-e).
Effects of chr-lip on MDA content and antioxidant enzymes activities: As shownin Table 1, the MDA level of model group rats was enhanced to 6.22±1.22 nmol mg-1 pro-1 compared with that of sham group 0.92±0.55 nmol mg-1 pro-1.
| Table 1: | Effects of Chr-lip on MDA content and SOD, GSH-Px activity in mice brain after MCAO (x±s, n = 6) |
 | |
| #p<0.01 vs. sham group; *p<0.01, **p<0.05 vs. model group | |
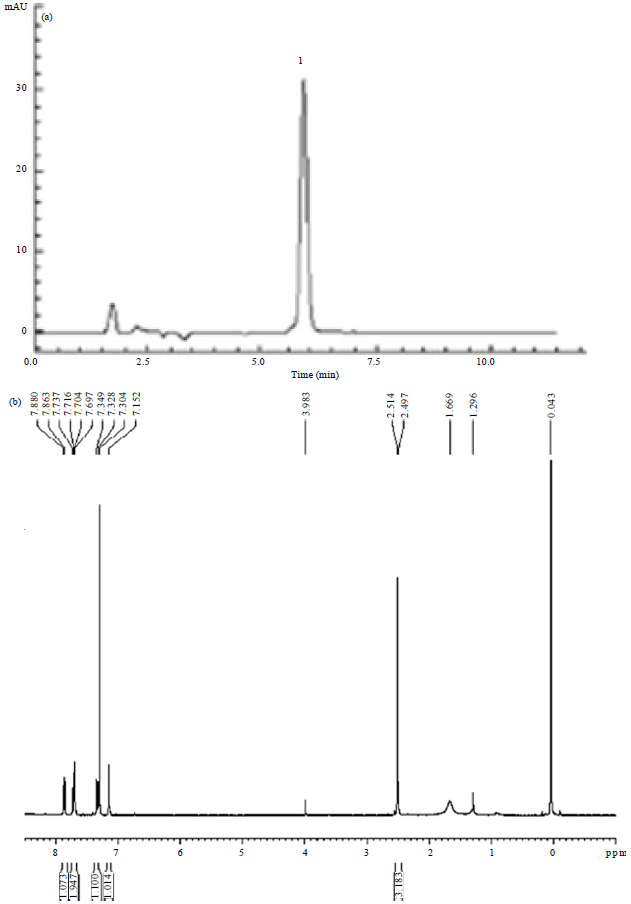 | |
| Fig. 2(a-b): | (a) HPLC Chromatograms and (b) 11-NMR of chrysophanol: 1 chrysophanol |
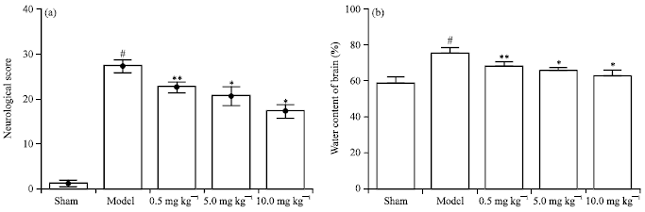 | |
| Fig. 3(a-b): | (a) Chr-lip preconditioning protected neurological function (n = 5-7) and (b) Reduced brain edema (n = 5 per group), following CIR. #p<0.01 compared with sham group, *p<0.01, **p<0.05 compared with model group |
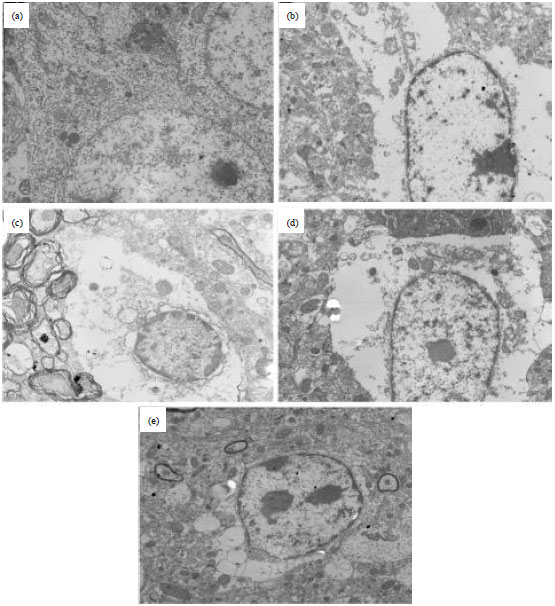 | |
| Fig. 4(a-e): | Transmission electron microscopic images of cerebral neuronal ultrastructure after 24 h of reperfusion (12,000x), (a) Sham group, (b) Model group, (c) Chr-lip-treated group (0.5 mg kg-1), (d) Chr-lip-treated group (5.0 mg kg-1) and (e) Chr-lip-treated group (10.0 mg kg-1) |
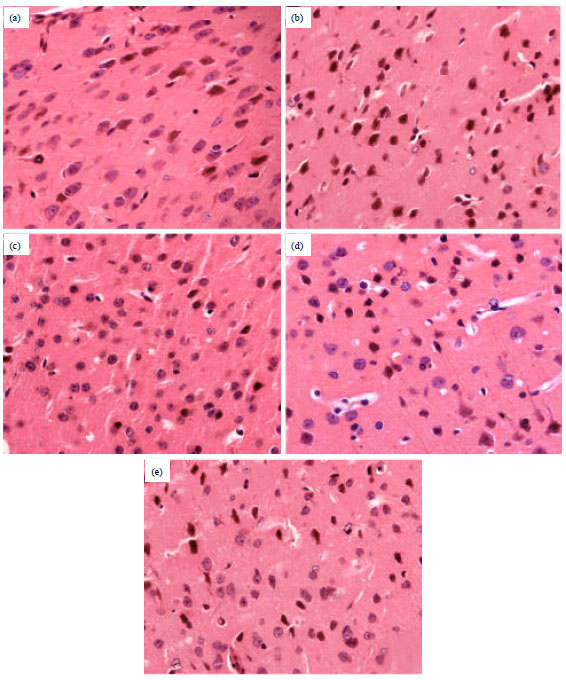 | |
| Fig. 5(a-e): | Representative coronal sections stained with Hematoxylin Eosin (HE) after 24 h of reperfusion (x400 magnifications), (a) Sham group, (b) Model group, (c) Chr-lip-treated group (0.5 mg kg-1), (d) Chr-lip-treated group (5.0 mg kg-1) and (e) Chr-lip -treated group (10.0 mg kg-1) |
The treatment with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) could diminish the MDA content significantly (p<0.01, p<0.01, p<0.05).
Lower levels of SOD and GSH-Px were detected in model group (102.80±6.97 and 25.33±1.52 U mg-1 pro-1) compared with those of sham group (258.91±15.92 and 44.67±2.51 U mg-1 pro-1) (p<0.01). Treatment with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) could up-regulate the levels of SOD obviously (p<0.01 vs. model group), meanwhile, increase the GSH-Px activity (p<0.01, p<0.01, p<0.05).
Effects of chr-lip on neuronal apoptosis: Neuronal injury in the ischemic hemispheres was analyzed by Hoechst33258 staining. Apoptotic cells were sparsely detected in the sham group; while the apoptotic rate was significantly increased in the model group compared with the sham group (p< 0.01).
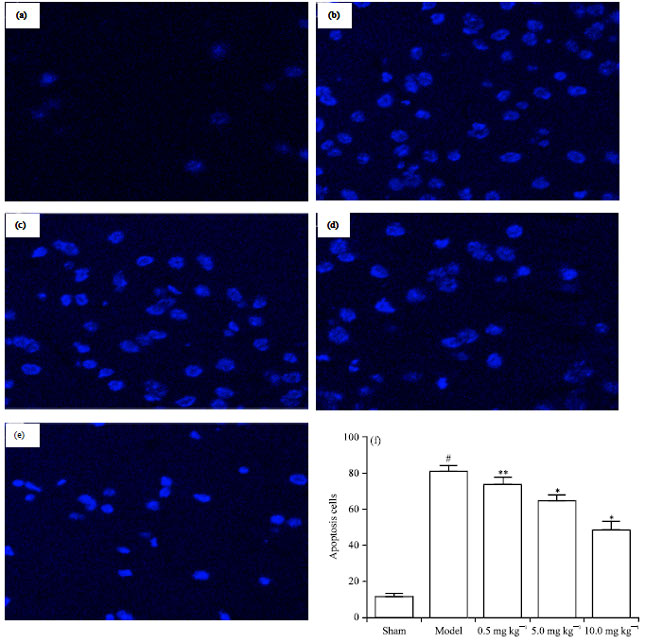 | |
| Fig. 6(a-f): | Chr-lip protect neuronal cell against cerebral ischemia-induced apoptosis (magnification, 400x), (a) Sham group, (b) Model group, (c) Chr-lip-treated group (0.5 mg kg-1), (d) Chr-lip -treated group (5.0 mg kg-1) and (e) Chr-lip-treated group (10.0 mg kg-1) |
The treatment with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) effectively attenuated the neuronal apoptosis caused by CIR injury, as indicated by significant reduction of apoptotic rate (p<0.01, p<0.01, p<0.05) (Fig. 6).
Western blot analysis: As seen in Fig. 7, the expression of Bax was significantly increased in model group compared with the sham group from 0.32±0.08 to 1.22±0.06 (p<0.01). In the treatment groups with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) group, could decrease the overexpression of Bax levels to 0.60±0.05, 0.82±0.05, 1.02±0.03 (p<0.01) compared with the model group. In the treatment groups with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) group, increased the Bcl-2 levels to 1.09±0.06, 0.93±0.03, 0.72±0.04 (p<0.01, p<0.01, p<0.05) compared with the model group. There was a significant exaltation of Cyt-C in model group which the relative ratio of Cyt-C/actin increased to 1.90±0.07 (p<0.01) compared with sham group (0.22±0.01).
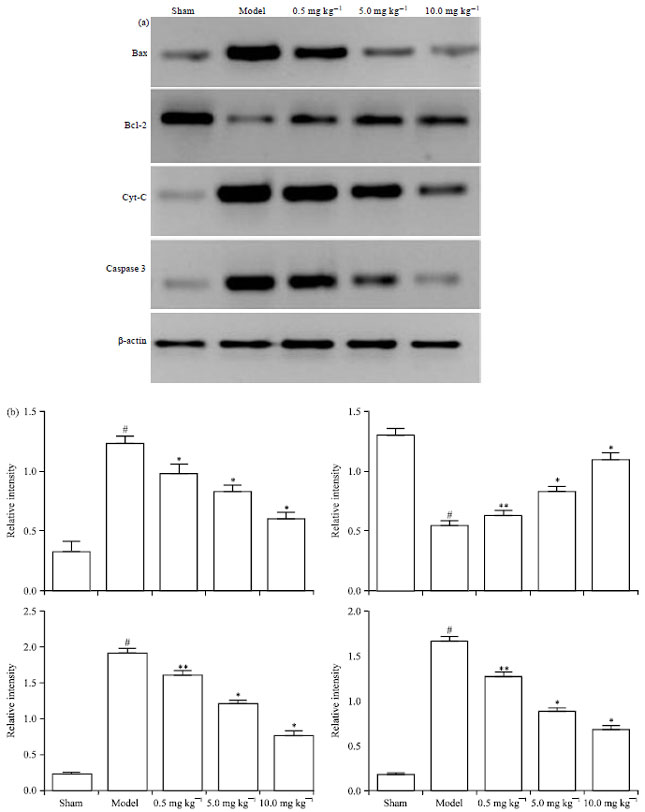 | |
| Fig. 7(a-b): | Western blot analyses of protein levels of Bax, Bcl-2, Cyt-C and Caspase3 (a) Western blot bands of Bax, Bcl-2, Cyt-C and Caspase3 expression in the ischemic hemispheres at 24 h after reperfusion, (b) Effect of Chr-lip (10.0, 5.0 or 0.5 mg kg-1) on the Bax, Bcl-2, Cyt-C and Caspase3 expression in ischemic hemispheres cortex at 24 h after reperfusion. #p<0.01 compared with sham group; *p<0.01, **p<0.05 compared with model group |
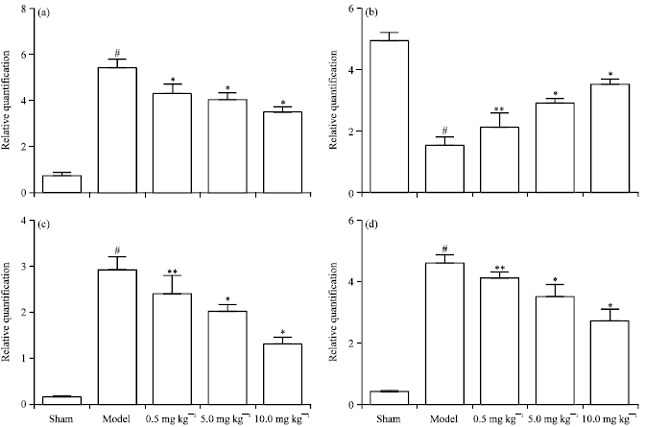 | |
| Fig. 8(a-d): | Effects of Chr-lip on the mRNA expressions of (a) Bax, (b) Bcl-2, (c) Cyt-C and (d) Caspase3 in mice. #p<0.01 compared with sham group; *p<0.01, **p<0.05 compared with model group |
In the treatment groups with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) group, expression of Cyt-C was decreased to 0.74±0.06, 1.37±0.04, 1.76±0.05 (p<0.01, p<0.01, p<0.05). There was a significant exaltation of Caspase3 in model group which the relative ratio of Caspase3/actin increased to 1.66±0.07 (p<0.01) compared with sham group (0.17±0.01). In the treatment groups with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) group, expression of Caspase3 was decreased to 0.69±0.04, 1.10±0.03, 1.48±0.05 (p<0.01, p<0.01, p<0.05).
Quantitative real-time PCR (qRT-PCR) analysis: There existed an excellent linear correlation (r = 0.993 for Bax, r = 0.995 for Bcl-2, r = 0.990 for Cyt-C, r = 0.996 for caspase-3 and r = 0.999 for GAPDH) between the Ct value and the logarithm of the DNA copy number. Figure 8 showed that the mRNA expression of Bcl-2 in the ischemic hemispheres was markedly decreased from 4.90±0.30 to 1.53±0.25 (p<0.01, n = 6) than that of sham group. Furthermore, administration of Chr-lip (10.0, 5.0 or 0.5 mg kg-1) increased the mRNA expression of Bcl-2 to 3.67±0.15, 2.73±0.15, 2.17±0.49 (p<0.01, p<0.01, p<0.05). In addition, caspase-3 mRNA in the ischemic hemispheres of model group was expressed much higher from 5.80±0.26 to 0.39±0.020 (p<0.01, n = 6) compared with the sham group. However, treatment with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) to the ischemic mice significantly decreased the mRNA expression of caspase-3 to 3.20±0.40, 4.00±0.43, 5.16±0.20 (p<0.01, p<0.01, p<0.05, n = 6), respectively, vs. the model group. Bax mRNA in the ischemic hemispheres of model group was expressed much higher from 6.90±0.36 to 0.70±0.15 (p<0.01, n = 6) compared with the sham group. However, treatment with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) to the ischemic mice significantly decreased the mRNA expression of Bax to 3.50±0.20, 4.36±0.32, 5.90±0.40 (p<0.01, n = 6), respectively, vs. the model group. Cyt-C mRNA in the ischemic hemispheres of model group was expressed much higher from 3.40±0.30 to 0.15±0.26 (p<0.01, n = 6) compared with the sham group. However, treatment with Chr-lip (10.0, 5.0 or 0.5 mg kg-1) to the ischemic mice significantly decreased to 1.43±0.15, 2.33±0.15 and 2.96±0.40 (p<0.01, p<0.01, p<0.05, n = 6), respectively, vs. the model group.
DISCUSSION
Chry has multiple pharmacological effects, including anti-aging, anti-inflammatory and anti-cancer activities. It has demonstrated the ability to inhibit growth of some human cancer cells, such as by inducing cell death (necrosis) in human lung cancer A549 cells and necrosis in J5 cells (Kim et al., 2010; Lu et al., 2010; Ni et al., 2012a, b). However, we found that Chr-lip had the obvious bioavailability and could improve passive avoidance learning and memory after CIR in mice (Li and Zhang, 2011). As the new formulation, it has the potent ablity to provide an opportunity for management neurological abnormalities in dementia conditions. In the present study, we showed abundance evidences that Chr-lip exhibited neuroprotective activity against CIR damage in a middle cerebral artery occlusion group. The neurological deficits, brain water content, neuronal ultrastructure and histopathological characterization reveal the neuronal necrosis and degeneration (Hritcu et al., 2012). Neurological deficits were found to be followed by neuronal degeneration and necrosis. The present study showed that pretreatment of Chr-lip improved the neurological deficit and neuronal degeneration. The data also revealed that pretreatment with Chr-lip could reduce brain water content, improving neurological, neuronal ultrastructure and histological deficits in CIR mice. These observations indicated that Chr-lip could efficiently prevent the brain injury induced by CIR.
CIR leads to diverse structural changes in neural cells. In the experiment, we observed the nuclear membrane was disrupted or disintegrated, swelling mitochondrial, decreased rough endoplasmic reticulum, disaggregation of polyribosomes and Golgi apparatus in postischemic ischemic hemisphere neurons which is consistent with the studies before (Li et al., 2012). Mitochondrial swelling is one of the initial postischemic changes. In mild ischemia, swollen mitochondria recover their normal shape soon. But, in severe ischemia, in neurons a large number of organelles show condensation, increased matrix density and deposits of an electron-dense material followed by the fracture of mitochondria (Petito and Pulsinelli, 1984). In the experiment, we proved that the cell nucleus was swollen and the nuclear membrane was disrupted or disintegrated, the chromatin was marginated and vacuoles were observed in the cytoplasm, mitochondrial cristae and membranes had mostly disappeared while degranulation of the rough endoplasmic reticulum and apoptotic bodies were apparent in model group. However, we also demonstrated slightly shrunken nucleus and complete nuclear membrane in the Chr-lip pretreatment group compared with the significantly aggregated chromatin toward the nuclear membrane in the model group.
During the period of CIR (Wang et al., 2013a), multiple pernicious processes including free radicals accumulation, inactivation of detoxification systems, overproduction of oxidants and consumption of antioxidants have been reported in many researchs (Chao et al., 2013; Yun et al., 2013). These changes disorder the normal antioxidative defense system in brain tissue (Chan, 2001). Then free radical will attack biofilm structure which further affect the stability of cell membrane, finally changes in the structure and function of the cell, aggravate cell injury and brain dysfunction. As the final product of lipid peroxidase, MDA is highly reactive and responsible for cytotoxic effects and neuronal death; its content can reflect the extent of lipid peroxidation and indirect react cell damage degree (Patockova et al., 2003). In the normal state of human body, endogenous antioxidant enzymes such as SOD, GSH-Px and GSH play important roles in the maintenance of redox homeostasis in the tissue (Wang et al., 2013b). Therefore, the body's ability to remove oxygen free radicals can use the activity of GSH-Px, SOD. In the present experiment, the findings have revealed, there was a significant decrease in the SOD and GSH-Px levels, with increase in the levels of MDA, in the brain homogenate of model group compared to sham group. In contrary, the elevated level of MDA was markedly decreased by treatment with Chr-lip, indicating that the neuroprotection conferred by Chr-lip may be attributed to attenuating lipid peroxidation following transient global cerebral ischemia. In addition, Chr-lip was found to be effective in stimulating the activities of SOD and GSH-Px. We demonstrated that when pretreatment with Chr-lip, MDA levels were obviously decreased and activities of SOD and GSH-Px were significantly increased which indicated that Chr-lip has a strong anti-oxidative effect on CIR injury.
Previous studies have demonstrated that apoptosis plays an important role in neuronal death following CIR (Li et al., 2009b). As we all known, The Bcl-2 families of proto-oncogenes encodes specific proteins such as Bax and Bcl-2 that critically regulate apoptosis (Abas et al., 2010). The anti-apoptotic effect of Bcl-2 occurs by prevention of Cyt-C release into the cytoplasm (Martinou and Youle, 2011). Activated Bax, the pro-apoptotic protein, promotes cell death, unless it is bound by either Bcl-2 or Bcl-XL (Kim et al., 2011). Compared with the sham group, the Bcl-2 expression was markedly decreased and Bax expression was markedly increased in the model group as reported in previous studies (Zhou et al., 2010; Chen et al., 2013). Xi et al. (2011) reported that propofol improved neurobehavioral outcome of CIR rats by the increasing expression of Bcl-2 while decreasing expression of Bax with CIR-induced rats at 24 h after ischemia. Wang et al. (2012a) demonstrated that esculetin exerted its anti-apoptotic activity through up-regulating the expression of Bcl-2 and down-regulating the expression of Bax, two apoptosis-related proteins. The current results indicate that treatment with Chr-lip after reperfusion significantly up-regulated the expression of Bcl-2 and down-regulated that of Bax in comparison to that observed in the CIR mice, suggesting that Chr-lip inhibited cerebral apoptosis after CIR and offered ideal therapeutic approach to CIR injury. However, caspase is a kind of protease family leading to apoptotic disintegration and executes the critical process of apoptosis in the apoptotic pathway (Degterev et al., 2003; Sung et al., 2012). Caspase3 is an established member of the caspase family known to be involved in the final execution phase of apoptosis and is one of the common down stream effect or sinvaried death receptor-mediated apoptosis pathway (Harrison et al., 2000; Teschendorf et al., 2008). A large number of evidences have testified that the Caspase3 has been up-regulated after CIR. But, previous studies (Torii et al., 2008) suggest that Caspase3 activation depends on Cyt-C release from the mitochondria via regulation by Bcl-2 and Bax, i.e., increased Bax expression promotes Cyt-C release and Bcl-2 plays an indirect role on Cyt-C release. Neohesperidin (Wang and Cui, 2013) inhibited MCAO-induced upregulation of Bax, Cyt-C and Caspase3, as well as the downregulation of Bcl-2. After leonurine treatment (Qi et al., 2009), there showed a marked decrease in the expression of Bax and an increase of Bcl-2. The attenuation of mitochondrial membrane swelling and content of Cyt-C in mitochondria isolated from ischemic cortex could also be observed in leonurine treated group. Besides, C.oil (Dohare et al., 2008) suppressed the rasied Bax protein level and assisted mitochondrial translocation and activation of Bcl-2 by changed mitochondrial membrane potential. It also inhibits the cytosolic release of Cyt-C, inhibits the activation of Caspase3 ultimately inhibiting apoptosis. In the present study, western blot and qRT-PCR analysis showed that a marked decrease in the expression of Bax and an increase of Bcl-2 in the treatment groups with Chr-lip; moreover, Chr-lip treatment decreased Cyt-C and Caspase3 expression. This suggested that suggesting that Chr-lip inhibited cerebral apoptosis after CIR and offered ideal therapeutic approach to ischemia cerebral injury.
CONCLUSION
In conclusion, we demonstrated that Chr-lip protected against CIR injury by reducing brain water content, improving neurological, neuronal ultrastructure and histological deficits and these beneficial effects were associated with inhibition of oxidative stress or neuronal apoptosis-related pathways, such as elevation of SOD and GSH-Px activities, reduction of the MDA content and the number of apoptotic cells, promotion of Bcl-2 expression, inhibition of Bax expression and Cyt-C release and suppression of Caspase3 activation. So, the protective mechanisms of Chr-lip against CIR injury might be involved to its anti-oxidant activities and anti-apoptotic.
ACKNOWLEDGMENTS
This study was supported by major projects of Hebei North University (ZD201310) and Hebei Province Science Foundation for Youths (H2014405031).
REFERENCES
- Abas, F., T. Alkan, B. Goren, O. Taskapilioglu, E. Sarandol and S. Tolunay, 2010. Neuroprotective effects of postconditioning on lipid peroxidation and apoptosis after focal cerebral ischemia/reperfusion injury in rats. Turk. Nneurosurg., 20: 1-8.
PubMed - Agarwal, S.K., S.S. Singh, S. Verma and S. Kumar, 2000. Antifungal activity of anthraquinone derivatives from Rheum emodi. J. Ethnopharmacol., 72: 43-46.
CrossRefPubMedDirect Link - Broughton, B.R.S., D.C. Reutens and C.G. Sobey, 2009. Apoptotic mechanisms after cerebral ischemia. Stroke, 40: e331-e339.
CrossRefPubMedDirect Link - Chan, P.H., 2001. Reactive oxygen radicals in signaling and damage in the ischemic brain. J. Cerebral Blood Flow Metab., 21: 2-14.
CrossRef - Chao, X.D., Y.H. Ma, P. Luo, L. Cao and W.B. Lau et al., 2013. Up-regulation of Heme oxygenase-1 attenuates brain damage after cerebral ischemia via simultaneous inhibition of superoxide production and preservation of NO bioavailability. Exp. Neurol., 239: 163-169.
CrossRef - Chen, L., Y. Xiang, L. Kong, X. Zhang, B. Sun, X. Wei and H. Liu, 2013. Hydroxysafflor yellow A protects against cerebral ischemia-reperfusion injury by anti-apoptotic effect through PI3K/Akt/GSK3β pathway in rat. Neurochem. Res., 38: 2268-2275.
CrossRefPubMedDirect Link - Chen, L., Y. Zhao, T. Zhang, X. Dang and R. Xie et al., 2014. Protective effect of Sheng-Nao-Kang decoction on focal cerebral ischemia-reperfusion injury in rats. J. Ethnopharmacol., 151: 228-236.
CrossRefDirect Link - Chen, Y.C., S.C. Shen, W.R. Lee, F.L. Hsu, H.Y. Lin, C.H. Ko and S.W. Tseng, 2002. Emodin induces apoptosis in human promyeloleukemic HL-60 cells accompanied by activation of caspase 3 cascade but independent of reactive oxygen species production. Biochem. Pharmacol., 64: 1713-1724.
CrossRefDirect Link - Clark, W.M., N.S. Lessov, M.P. Dixon and F. Eckenstein, 1997. Monofilament intraluminal middle cerebral artery occlusion in the mouse. Neurol. Res., 19: 641-648.
PubMed - Dohare, P., S. Varma and M. Ray, 2008. Curcuma oil modulates the nitric oxide system response to cerebral ischemia/reperfusion injury. Nitric Oxide, 19: 1-11.
CrossRefDirect Link - Guan, J., H. Li, T. Lv, D. Chen, Y. Yuan and S. Qu, 2013. Bone morphogenetic protein-7 (BMP-7) mediates ischemic preconditioning-induced ischemic tolerance via attenuating apoptosis in rat brain. Biochem. Biophys. Res. Commun., 441: 560-566.
CrossRefDirect Link - Harrison, D.C., A.D. Medhurst, B.C. Bond, C.A. Campbell, R.P. Davis and K.L. Philpott, 2000. The use of quantitative RT-PCR to measure mRNA expression in a rat model of focal ischemia-caspase-3 as a case study. Mol. Brain Res., 75: 143-149.
CrossRefDirect Link - He, Y., H. Wan, Y. Du, X. Bie, T. Zhao, W. Fu and P. Xing, 2012. Protective effect of Danhong injection on cerebral ischemia-reperfusion injury in rats. J. Ethnopharmacol., 144: 387-394.
CrossRefDirect Link - Hritcu, L., O. Cioanca and M. Hancianu, 2012. Effects of lavender oil inhalation on improving scopolamine-induced spatial memory impairment in laboratory rats. Phytomedicine, 19: 529-534.
CrossRefDirect Link - Iizuka, A., O.T. Iijima, K. Kondo, H. Itakura and F. Yoshie et al., 2004. Evaluation of Rhubarb using antioxidative activity as an index of pharmacological usefulness. J. Ethnopharmacol., 91: 89-94.
CrossRefDirect Link - Kim, D.H., E.K Park, E.A Bae and M.J. Han, 2000. Metabolism of rhaponticin and chrysophanol 8-o-beta-D-glucopyranoside from the rhizome of rheum undulatum by human intestinal bacteria and their anti-allergic actions. Biol. Pharm. Bull., 23: 830-833.
PubMed - Kim, H.S., M.S. Park, J.K. Lee, H.J. Kim, J.T. Park and M.C. Lee, 2011. Time point expression of apoptosis regulatory proteins in a photochemically-induced focal cerebral ischemic rat brain. Chonnam Med. J., 47: 144-149.
CrossRef - Kim, S.J., M.C. Kim, B.J. Lee, D.H. Park, S.H. Hong and J.Y. Um, 2010. Anti-Inflammatory activity of chrysophanol through the suppression of NF-κB/Caspase-1 activation in vitro and in vivo. Molecules, 15: 6436-6451.
CrossRefPubMedDirect Link - Li, H., C.Q. Deng, B.Y. Chen, S.P. Zhang, Y. Liang and X.G. Luo, 2009. Total saponins of Panax Notoginseng modulate the expression of caspases and attenuate apoptosis in rats following focal cerebral ischemia-reperfusion. J. Ethnopharmacol., 121: 412-418.
CrossRefDirect Link - Li, J.S., W. Zhang, Z.M. Kang, S.J. Ding and W.W. Liu et al., 2009. Hyperbaric oxygen preconditioning reduces ischemia-reperfusion injury by inhibition of apoptosis via mitochondrial pathway in rat brain. Neuroscience, 159: 1309-1315.
CrossRefDirect Link - Li, T., N. Wang and M. Zhao, 2012. Neuroprotective effect of phosphocreatine on focal cerebral ischemia-reperfusion injury. J. Biomed. Biotechnol., Vol. 2012.
CrossRef - Liu, Y., T. Nakamura, T. Toyoshima, F. Lu and K. Sumitani et al., 2014. Ameliorative effects of yokukansan on behavioral deficits in a gerbil model of global cerebral ischemia. Brain Res., 1543: 300-307.
CrossRefDirect Link - Manzanero, S., T. Santro and T.V. Arumugam, 2013. Neuronal oxidative stress in acute ischemic stroke: Sources and contribution to cell injury. Neurochem. Int., 62: 712-718.
CrossRefDirect Link - Martinou, J.C. and R.J. Youle, 2011. Mitochondria in apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev. Cell, 21: 92-101.
CrossRefDirect Link - Ni, C.H., P.Y. Chen, H.F. Lu, J.S. Yang and H.Y. Huang et al., 2012. Chrysophanol-induced necrotic-like cell death through an impaired mitochondrial ATP synthesis in Hep3B human liver cancer cells. Arch. Pharm. Res., 35: 887-895.
CrossRefDirect Link - Ni, C.H., C.S. Yu, H.F. Lu, J.S. Yang and H.Y. Huang et al., 2012. Chrysophanol-induced cell death (necrosis) in human lung cancer A549 cells is mediated through increasing reactive oxygen species and decreasing the level of mitochondrial membrane potential. Environ. Toxicol.
CrossRef - Patockova, J., P. Marhol, E. Tumova, M. Krsiak, R. Rokyta, S. Stipek, J. Crkovska and M. Andel, 2003. Oxidative stress in the brain tissue of laboratory mice with acute post insulin hypoglycemia. Physiol. Res., 52: 131-135.
PubMedDirect Link - Petito, C.K. and W.A. Pulsinelli, 1984. Delayed neuronal recovery and neuronal death in rat hippocampus following severe cerebral ischemia: Possible relationship to abnormalities in neuronal processes. J. Cerebral Blood Flow Metab., 4: 194-205.
CrossRef - Qi, J., Z.Y. Hong, H. Xin and Y.Z. Zhu, 2009. Neuroprotective effects of leonurine on ischemia/reperfusion-induced mitochondrial dysfunctions in rat cerebral cortex. Biol. Pharm. Bull., 33: 1958-1964.
PubMed - Shi, Y.Q., T. Fukai, H. Sakagami, J. Kuroda and R. Miyaoka et al., 2001. Cytotoxic and DNA damage-inducing activities of low molecular weight phenols from rhubarb. Anticancer Res., 21: 2847-2853.
PubMed - Sung, J.H., M.O. Kim and P.O. Koh, 2012. Ferulic acid attenuates the focal cerebral ischemic injury-induced decrease in parvalbumin expression. Neurosci. Lett., 516: 146-150.
CrossRefDirect Link - Ten, V.S. and A. Starkov, 2012. Hypoxic-ischemic injury in the developing brain: The role of reactive oxygen species originating in mitochondria. Neurol. Res. Int., Vol. 2012.
CrossRef - Teschendorf, P., S.A. Padosch, F. Spohr, M. Albertsmeier and A. Schneider et al., 2008. Time course of caspase activation in selectively vulnerable brain areas following global cerebral ischemia due to cardiac arrest in rats. Neurosci. Lett., 448: 194-199.
CrossRefDirect Link - Torii, K., K. Nishizawa, A. Kawasaki, Y. Yamashita and M. Katada et al., 2008. Anti-apoptotic action of Wnt5a in dermal fibroblasts is mediated by the PKA signaling pathways. Cell. Signalling, 20: 1256-1266.
CrossRefDirect Link - Wang, C., A. Pei, J. Chen, H. Yu and M.L. Sun et al., 2012. A natural coumarin derivative esculetin offers neuroprotection on cerebral ischemia/reperfusion injury in mice. J. Neurochem., 121: 1007-1013.
CrossRefDirect Link - Wang, D., X. Yuan, T. Liu, L. Liu, Y. Hu, Z. Wang and Q. Zheng, 2012. Neuroprotective activity of lavender oil on transient focal cerebral ischemia in mice. Molecules, 17: 9803-9817.
CrossRefDirect Link - Wang, J.J. and P. Cui, 2013. Neohesperidin attenuates cerebral ischemia-reperfusion injury via inhibiting the apoptotic pathway and activating the Akt/Nrf2/HO-1 pathway. J. Asian Nat. Prod. Res., 15: 1023-1037.
CrossRefDirect Link - Wang, L., Y. Huang, J. Wu, G. Lv, L. Zhou and J. Jia, 2013. Effect of Buyang Huanwu decoction on amino acid content in cerebrospinal fluid of rats during ischemic/reperfusion injury. J. Pharm. Biomed. Anal., 86: 143-150.
CrossRefPubMedDirect Link - Wang, T.F., Z. Lei, Y.X. Li, Y.S. Wang and J. Wang et al., 2013. Oxysophoridine protects against focal cerebral ischemic injury by inhibiting oxidative stress and apoptosis in mice. Neurochem. Res., 38: 2408-2417.
CrossRefPubMedDirect Link - Xi, H.J., T.H. Zhang, T. Tao, C.Y. Song, S.J. Lu, X.G. Cui and Z.Y. Yue, 2011. Propofol improved neurobehavioral outcome of cerebral ischemia-reperfusion rats by regulating Bcl-2 and Bax expression. Brain Res., 1410: 24-32.
CrossRefPubMedDirect Link - Yan, Y., Y. Min, H. Min, C. Chao, Q. Ying and H. Zhi, 2014. N-butanol soluble fraction of the water extract of chinese toon fruit ameliorated focal brain ischemic insult in rats via inhibition of oxidative stress and inflammation. J. Ethnopharmacol., 151: 176-182.
CrossRefPubMedDirect Link - Yun, X., V.D. Maximov, J. Yu, H. Zhu, A.A. Vertegel and M.S. Kindy, 2013. Nanoparticles for targeted delivery of antioxidant enzymes to the brain after cerebral ischemia and reperfusion injury. J. Cereb. Blood Flow Metab., 33: 583-592.
CrossRefPubMedDirect Link - Zhao, J., S. Yu, W. Zheng, G. Feng, G. Luo, L. Wang and Y. Zhao, 2010. Curcumin improves outcomes and attenuates focal cerebral ischemic injury via antiapoptotic mechanisms in rats. Neurochem. Res., 35: 374-379.
CrossRefPubMedDirect Link - Zhou, Z.Y., Y.P. Tang, J. Xiang, P. Wua and H.M. Jin et al., 2010. Neuroprotective effects of water-soluble Ganoderma lucidum polysaccharides on cerebral ischemic injury in rats. J. Ethnopharmacol., 131: 154-164.
CrossRefPubMedDirect Link