
Mohamed Mansour El-Loly
Department of Dairy Sciences, National Research Centre, Dokki, Cairo, Egypt
Mohamed Bahy Mahfouz
Department of Dairy Sciences, National Research Centre, Dokki, Cairo, Egypt
International Journal of Dairy Science
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 79-111







ABSTRACT
Lactoferrin (LF) is an iron-binding glycoprotein member of the Transferrin (TF) family which present in milk, other exocrine secretions and neutrophil granules in mammals, it is highly conserved among human and other mammals' species. First identified in milk as the red milk protein due to its iron content, LF has since attracted much interest over the past fifty years. LF is considered to be an important host defense molecule and has a diverse range of physiological functions such as antimicrobial/antiviral activities, immunomodulatory activity and antioxidant activity. During the past decade, it has become evident that oral administration of LF exerts several beneficial effects on the health of humans and animals, including anti-infective, anticancer and anti-inflammatory effects. This has enlarged the application potential of LF as a food additive. The technology of producing bovine LF on a large scale was established over 20 years ago. This review summarizes our current understanding of the properties of physico-chemical of LF, physiological functions and technological characteristics as well as nutritional and applications relationships that explain the roles of LF in host defense.
PDF Abstract XML References Citation
Received: February 24, 2011;
Accepted: April 12, 2011;
Published: June 03, 2011
How to cite this article
Mohamed Mansour El-Loly and Mohamed Bahy Mahfouz, 2011. Lactoferrin in Relation to Biological Functions and Applications: A Review. International Journal of Dairy Science, 6: 79-111.
DOI: 10.3923/ijds.2011.79.111
URL: https://scialert.net/abstract/?doi=ijds.2011.79.111
DOI: 10.3923/ijds.2011.79.111
URL: https://scialert.net/abstract/?doi=ijds.2011.79.111
INTRODUCTION
Colostrum and milk is a vital nutritional source for the offspring of all mammals, including humans. In addition to its nutritional value, it is a rich source of proteins including Lactoferrin (LF) (Jenssen and Hancock, 2009). Also, milk contains various protective proteins that, if properly activated, can contribute to the preservation of milk (IDF, 1988). Milks from different species differ significantly as to their protective proteins content. Thus human milk contains almost 10 times more LF than doe's bovine milk (Reiter, 1985).
Also, whey proteins are used as common ingredients in various products including infant formulas, specialized enteral and clinical protein supplements and sports nutrition products with the expectation of the therapeutic potential of whey proteins and peptides. LF, one of the major whey proteins, is a red iron-binding protein present mainly in external secretions such as breast milk and in polymorphonuclear neutrophils. This protein plays an important role in the defense mechanism of mucosal surfaces, since in an iron-depleted state it has bacteriostatic properties (Masson and Heremans, 1966). LF is released from polymorphonuclear neutrophils on activation of these cells and its presence in body fluids is proportional to the flux of neutrophils (Rado et al., 1984; Guerrant et al., 1992; Martins et al., 1995; Parsi et al., 2008).
LF belongs to the family of iron-binding proteins and exhibits a wide spectrum of antimicrobial and immunotropic properties. It is particularly resistant to proteolytic degradation in alimentary tract, in contrast to other milk proteins, e.g., casein. In any case, LF-derived peptides also possess potent antibacterial activities which absorbed from the intestine by means of specific receptors located on brush border cells. Administered orally, LF stimulates both local and systemic immune response. It plays a role in the absorption of nutrients. The protein can deliver such metal ions as iron, manganese and zinc and facilitate the absorption of sugars (Artym and Zimecki, 2005).
LF has a truly multifunctional protein that has been studied extensively over the past decades. It is best known for its ability to bind iron which eventually led to the discovery of its biological functions, including antimicrobial, antiviral, antioxidant, antiinflammatory activities, immunomodulation, modulation of cell growth and inhibition of several bioactive compounds, such as lipopolysaccharide (LPS) and glycosamino-glycan (Baveye et al., 1999; Chierici, 2001; Jenssen and Hancock, 2009). It also provides a defense against gastro-intestinal infections, participates in local secretory immune systems (Valenti et al., 1998; Vorland, 1999; Steijns and van Hooijdonk, 2000), in synergism with some immunoglobulins (Igs) such as IgG and other protective proteins, supplies an iron-binding antioxidant protein in tissues and possibly promotes growth of animal cells, such as lymphocytes and intestinal cells (Lonnerdal and Iyer, 1995; Meulenbroek and Zeijlemaker, 1996).
The alteration of the activity of these antimicrobial factors in cow’s milk could have an impact on the shelf life of raw milk and on the development of additional health and functional foods based upon these factors. The composition of different milk samples is usually not uniform; therefore, the concentrations of several milk constituents change during the lactation period and differ from one mother to the next. There are several factors that are known to influence the concentration of milk constituents in predictable ways (Levieux et al., 2002; Cheng et al., 2008). These include lactation stage, breastfeeding routine, parity, age and other maternal characteristics such as regional differences and, in some situations, season of the year and maternal diet.
On the other hand, Igs (antibodies) are protective proteins that are important in the transfer of passive immunity from the mother to the child. The young of many mammalian species are born without an effective immune system, therefore the Igs and LF exhibit antimicrobial activity and protect the neonate from infection until their own immune system has developed. The increasing commercial interest in exploiting the therapeutic value of LF and IgG has stimulated the need for reliable assays for their determination at the endogenous level in milk (Hurley et al., 1993; Elagamy et al., 1996; Indyk and Filonzi, 2005).The in vitro activity of LF also includes transcriptional activation of several genes (Oh et al., 2004). Tomita et al. (1991) found that pepsin-hydrolysate of LF (LFhyd) has more potent antimicrobial activity than the native protein and they purified the active peptide from LFhyd. The antimicrobial peptide derived from Lfhyd was named lactoferricin (LFcin) (Bellamy et al., 1992a). Interestingly, LFcin and its derivatives exhibit various biological activities, like LF. Therefore, the LFcin-region seems likely to be an important functional domain of LF (Wakabayashi et al., 2003).
LF content in milk varies depending on the species. The amount of LF is lower in cows' milk (i.e., 0.1-0.4 mg mL-1) than in human milk (i.e., l-3 mg mL-1). However, a factory scale technology to produce large amounts of bovine LF at high purity from cow’s milk was established over 20 years ago (Law and Reiter, 1977).
Recently it has been recognized that oral administration of LF exerts various health beneficial effects such as anti-infective activities not only in infants but also in adult animals and humans (Tomita et al., 2002; Teraguchi et al., 2004; Weinberg, 2007). Weinberg (2007) reported that recombinant bovine and human LF is available for development into nutraceutical, preservative and pharmaceutical products. Among conditions for which the products were being investigated were: angiogenesis, bone remodeling, food preservation, infection in animals, humans, plants, neoplasia in animals, humans, inflammation in intestine, joints, wound healing as well as enhancement of antimicrobial and antineoplastic drugs and prevention of iron induced oxidation of milk formula.
PROPERTIES OF PHYSICO-CHEMICAL OF LACTOFERRIN HISTORY AND CONCENTRATION OF LF IN DIFFERENT SPECIES
LF is an iron-binding glycoprotein of the TF family which was first fractionated as an unknown “red fraction” from cow’s milk by Sorensen and Sorensen (1940) and later in human milk by Schafer (1951). The red protein from both human and bovine milk was defined as a lactotransferrin-like glycoprotein or lactosiderophilin because of its high similarity to TF and siderophilin in blood and ovotransferrin in egg which in United Kingdom (Groves, 1960), in Sweden (Johansson and Hjerten, 1960b), in France (Montreuil et al., 1960). LF was first isolated from cow’s milk and then from human milk (Ensminger and Esminger, 1986; Wood, 1988; Spik et al., 1998; Shimazaki, 2000a, b).
LF was present in large quantities by several groups not only in milk secreted by the mammary gland but also in various exocrine mammalian secretions such as tears, saliva, seminal fluid, cervical mucus, bronchial secretions and in some white blood cells (i.e., neutrophilic leucocytes) (Levay and Viljoen, 1995; Steijns, 2001). Because LF is predominantly found in the products of the exocrine glands of the digestive, respiratory and reproductive systems, it is thought that LF has a role in the host non-specific defense against invading pathogens.
LF concentration varied between 31.78 and 485.63 (μg mL-1 in milk from normal animals (Cheng et al., 2008). It is concentration is higher than 2 mg mL-1 in human milk (Nagasawa et al., 1972) and in the range of 0.02-0.2 mg mL-1 in bovine milk (Masson and Heremans, 1971; Suzuki et al., 1977). The LF concentration in guinea-pig, mouse and horse milk is in the range of 0.2 to 2 mg mL-1 as well as that in rat, rabbit and dog milk is lower than 0.05 mg mL-1 (Masson and Heremans, 1971).
Colostrum milk contains more LF than mature milk. It should be noted that LF is the second most abundant whey protein in human milk as shown in Fig. 1. While its concentrations in other secretory fluids are shown in Table 1 (Shimazaki, 2000a).
LF was significantly associated with stage of lactation (r = 0.557) and daily milk production (r = -0.472). Nevertheless, there was no significant relationship with parity. Moreover, milk LF concentration tended to be correlated with the somatic cell count score (r = 0.375). This finding suggests that milk LF may be helpful as an indicator for intramammary infection in dairy cows (Cheng et al., 2008).
Harmon et al. (1975) reported that the LF concentration of milk was significantly associated with somatic cell count (SCC), levels of bovine serum albumin, stage of lactation and milk production. Still, LF had a negative relationship with milk production.
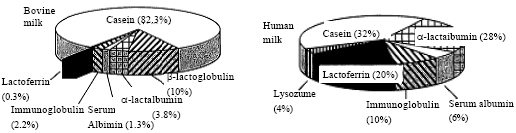 | |
| Fig. 1: | Milk protein fractions content (%) of human and bovine milk. (Total protein concentration in human milk is 1 g 100-1 mL and that in bovine milk is 3.2 g 100-1 mL) |
| Table 1: | Occurrence of lactoferrin in human external fluids |
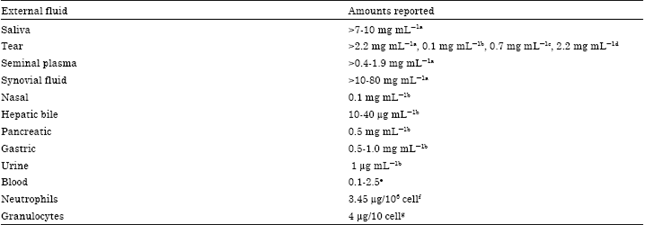 | |
| a: Steijns and van Hooijdonk (2000), b: Masson et al. (1966), c: Takayanagi et al. (1986), d: Kijlstra et al. (1983), e: Malmquist et al. (1978), f: Moguilevsky et al. (1987), g: Bezwoda and Mansoor (1989) | |
Tsuji et al. (1990) reported that the highest LF content in colostrum was observed in second lactation. After the third lactation, no differences in LF content were observed. Hagiwara et al. (2003) reported that the concentration of milk LF was significantly related to the age of cows but not to the stage of lactation; however, because those samples were from different mammary gland quarters and the sample number was relatively small.
SYNTHESIS OF LACTOFERRIN
Teng et al. (2002) reported that the LF synthesis can be continuous (exocrine fluids), under hormonal control (genital tract, mammary gland) [213]. LF is secreted in the apo-form from epithelial cells in most exocrine fluids such as saliva, bile, pancreatic and gastric fluids, tears and milk (Montreuil et al., 1960). LF is mainly synthesized by glandular epithelial cells; its concentration in humans may vary from 1 g L-1 (mature milk) to 7 g L-1 (colostrum). The mean concentration is 30 mg L-1 in mature bovine milk. In addition, LF is synthesized during the transition from promyelocytes to myelocytes and is thus a major component of the secondary granules of PMNs (Masson et al., 1966). LF levels of biological fluids may increase greatly and constitute a marker for inflammatory diseases (Mann et al., 1994; Legrand et al., 2008) noticeable this is in plasma, where the LF concentration can be as low as 0.4-2 mg L-1 under normal conditions but increases to 200 mg L-1 in septicemia.
ISOLATION AND PURIFICATION OF LACTOFERRIN
Acid precipitated casein has been used as the starting material for isolation of LF from bovine milk (Groves, 1960). However, the whey fraction of milk or colostrum is better source to obtain LF on laboratory scale (Law and Reiter, 1977) and cheese whey is another source used to obtain LF on a large scale.
Other methods used include affinity chromatography with immobilized materials such as heparin (Blackberg and Hernell, 1980), blue dye (Bezwoda and Mansoor, 1986), β-lactoglobulin (Ena et al., 1990), antilactoferrin antibody (Kawakami et al., 1987) or single-standard DNA (Hutchens et al., 1989b). Also, metal-chelate affinity chromatography (Hutchens et al., 1989a) and hydroxyapatite column chromatography (Itagaki et al., 1993) have been used to purify LF.
ELISA and other immunodiffusion and immunoelectrodiffusion methods have been employed using anti-lactoferrin antiserum. In order to measure the LF concentrations in dairy products such as cheese, treatment at pH 4.0 to release LF to casein is necessary.
LF is isolated and purified on an industrial scale (approximately 20-30 tons annually worldwide) from cheese whey and skim milk. The concentration of LF in cheese whey is roughly 100 mg L-1. Since LF exists as a cationic protein (isoelectric point of LF is alkaline) in whey, it is readily adsorbed to a cation-exchange chromatography resin and then eluted using salt solutions. The eluted crude LF is desalted and concentrated using ultrafiltration and diafiltration membranes, after which it is subjected to pasteurization. Purified LF powder with a purity of 95% or higher is finally obtained by freeze-drying. In an alternative process, microfiltration and spray-drying are performed instead of pasteurization and freeze-drying, respectively. Pasteurization has come to be considered as very important in order to inactivate not only bacteria but also viruses such as foot and mouth disease virus. While making efforts to develop a practical method for the pasteurization of LF which is stable against heat treatment under acidic conditions (Abe et al., 1991) while heat treatment at a neutral pH causes denaturation of the protein. It is considered that heating at a pH 4 and to a temperature of 90-100°C for 5-10 min as well as the UHT method are suitable and practical methods for the pasteurization of LF. This pasteurization process was patented and it has been applied to the manufacture of a wide variety of commercial products containing LF. A pepsin hydrolysate of LF is produced by treatment with porcine pepsin under acidic conditions (Saito et al., 1991). After hydrolysis has been completed, pepsin is inactivated by heat treatment. Then the reaction mixture is filtered and concentrated by reverse osmosis. Finally, the hydrolysate of LF is obtained by pasteurization and freeze-drying for use in infant formula.
In addition, LFcin can be purified from this LF hydrolysate by two-step hydrophobic chromatography. The peptide is eluted with an acidic buffer, the eluted solution is concentrated by reverse osmosis and finally, LFcin is produced by freeze-drying as a powder with over 95% purity. This production process for LFcin has also been patented (Bellamy et al., 1992b).
Abd El-Gawad et al. (2003) found that immuno-precipitation of rbLF with anti-bLF in the medium at pH 7.5 showed two forms of LF, LF-a with a molecular weight of 84 kDa and LF-b with 80 kDa, while the immuno-precipitation at pH 8.0 produced one single band of LF-b with 80 kDa.
Adam et al. (2008) employed SDS-PAGE and Experion to isolate LF from the colostrum and normal milk samples with technique ion exchange chromatography, using monolithic column. Coomassie blue staining followed and the expressive band of LF at molecular weight 77 kDa was detectable (Fig. 2a line 2 and 3). Similarly to Coomassie blue staining by silver stained, the majority bands of LF were also visible (Fig. 2b). The result of the electrophoretic analysis is shown in Fig. 2c. Both techniques show that many other proteins are presented in the samples (Fig. 2).
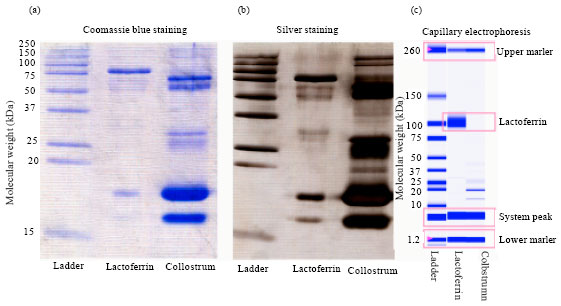 | |
| Fig. 2: | SDS-PAGE with (a) Coomassie blue staining, (b) Silver staining and (c) Experion capillary electrophoresis of ladder, lactoferrin standard (100 μg mL-1) and milk sample |
STRUCTURE OF LACTOFERRIN
LF is a glycoprotein with a molecular weight of average about 80 kDa which shows high affinity for iron. The molecular structure and amino acid sequence of hLF were discovered in 1984. LF was then classified as a member of the TF family, due to its 60% sequence identity with serum TF. Human LF (hLF) has a molecular weight of 82.4 kDa and is composed of 702 (Metz-Boutigue et al., 1984) or 692 (Powell and Ogden, 1990; Rey et al., 1990) amino acid residues, the sequence identity is 69%. Bovine LF (bLF) has a molecular weight of 83.1 kDa and is composed of 689 amino acid residues (Pierce et al., 1991).
Three dimensional structures of hLF (Farnaud and Evans, 2003), bLF (Moore et al., 1997), horse LF and buffalo LF (Sharma et al., 1998) have been determined by X-ray crystallographic analysis. The three dimensional structures of bovine and human LF are very similar but not identical which are shown in Fig. 3.
Three different isoforms of LF have been isolated. LF-α is the iron binding form but has no ribonuclease activity. On the other hand, LF-β and LF-γ demonstrate ribonuclease activity but they are not able to bind iron (Furmanski et al., 1989). LF is comprised of a single polypeptide chain containing 703 amino acids folded into two globular lobes. These lobes, also called C-(carboxy) and N-(amino) terminal regions are connected with α-helix. Each lobe consists of two domains known as C1, C2, N1 and N2. The domains create one iron binding site on each lobe. LF molecules contain (according to the species and protein) varying numbers of sites for potential glycosylation, mostly on the surface of the molecule. The most common saccharine is mannose; around 3% are hexoses and 1% hexosamines. The degree of glycosylation varies and determines the rate of resistance to proteases or to very low pH (Anderson et al., 1987). Jameson et al. (1998) observed two structures for LF: an open conformation, originally described for the iron-free LF and a closed conformation, mainly observed with the iron-saturated molecule. The conformational transition could be involved in basic functions such as transportation and catalysis.
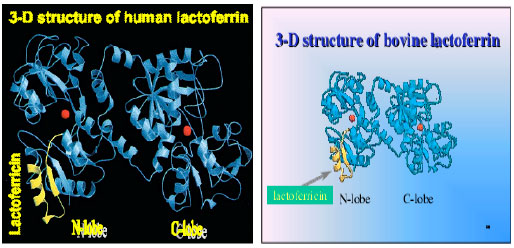 | |
| Fig. 3: | Three dimensional structures of diferric human LF (Farnaud and Evans, 2003) and bovine LF (Moore et al., 1997). The location of lactoferrcin within the protein is shown in yellow and the two ferric ions are in red |
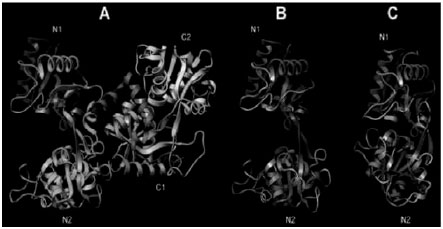 | |
| Fig. 4: | Structure of hLF. (A) Ribbon diagram showing the polypeptide folding of iron-saturated hLF. The N-t lobe is on the left; the polypeptide chain is colored from the N- to the C-terminal end according to a red-shift. (B) Open and (C) closed structures of the N-terminal lobe of hLF (α-helices are colored in magenta and b-sheets in blue). Domains N1, N2, C1 and C2 are indicated. Domains, referred to as N1 and N2, or C1 and C2, delineate a deep cleft within which the iron binding site is located. The a/b fold of each domain consists of a central, mostly parallel b-sheet, with a helix packed against it. The helical N-terminus faces the interdomain cleft, making it somewhat positively charged and one of the helices, H5 from the N2 (or C2) domain, serves as the binding site for the essential carbonate anion at the metal binding site (Legrand et al., 2008) |
According to crystallographic data, the domains move essentially as rigid bodies (Fig. 4) that close over the bound metal or open to release it.
There are three forms of LF according to its iron saturation: apo-LF (iron free), monoferric form (one ferric ion) and holo-LF (binds two Fe3+ ions). The tertiary structure in holo-LF and apo-LF is different (Jameson et al., 1998).
The LF fraction separated from colostrum and milk of buffalo, cow (Local, Friesian and Brown-Swiss breeds), ewe (Rahmani and Ossemi breeds), goat as well as camel were patterned using SDS-PAGE as a single band almost of similar mobility as standard LF. Also, the LF fractions of different species were subjected to reverse phase HPLC and only one peak was apparent. LF concentrations varied considerably from one species to others (Abd El-Gawad et al., 1996).
Immunoprecipitation of recombinant bovine LF with anti-bovine LF in the medium at pH 7.5 showed two forms of LF, the first named LFa (84 kDa) and the second one LFb (80 kDa) while the immunopercipitation at pH 8 produced on single band of LFa (80 kDa) (Abd El-Gawad et al., 2001a).
LACTOFERRCIN (LFCIN) STRUCTURE
Limited proteolysis leads to the release of LF fragments: N-t and C-t lobes, the N-2 domain (Legrand et al., 1984) and LFcin (Bellamy et al., 1992b). Lfcin (Lfcin-B from bovine LF and LFcin-H from human LF) is a 25 amino acid peptide (residues 17-42) including two Cys residues linked by a disulfide bridge and containing many hydrophobic and positively charged residues. The secondary structure of Lfcin is markedly different from the same sequence in intact LF (Gifford et al., 2005). The long α-helix observed in the LF structure is replaced by a single β-sheet strand. This structure seems to be better suited for making contact with bacterial membranes. In biological fluids, LF exists in an iron-free form that is very susceptible to proteolysis. It cannot be overlooked that a posttranslational process of maturation by proteolysis leads to the release of LF-derived active peptides in biological fluids (Goldman et al., 1990).
METABOLISM OF LACTOFERRIN
Levay and Viljoen (1995) observed that there are two ways in which LF can be eliminated from the organism: either through receptor-mediated endocytosis of phagocytic cells (macrophages, monocytes and other cells belonging to the reticuloendothelial system) with subsequent iron transfer to ferritin or through direct uptake by the liver. Endocytosis performed by Kupffer cells, liver endothelial cells and hepatocytes contributes to LF removal. Moreover, Hutchens et al. (1991) achieved that the kidneys seem to be involved in the removal of LF from the circulation since LF and its fragments, mainly of maternal origin, have been found in the urine of breast-fed infants.
RECEPTORS OF LACTOFERRIN
In fact, most molecular targets on the host cells are multiligand receptors and, interestingly, as reviewed hereinafter, many of them were reported as signaling, endocytosis and nuclear targeting molecules. Ling and Schryvers (2006) showed that the search for specific LF receptors, comparable with that of TF, has consistently mobilized the energy of researchers. Surprisingly, LF receptors with the highest specificity were discovered on bacteria, whereas specific mammalian receptors were only encountered on enterocytes (Suzuki et al., 2005).
LF binding proteins have been found on the small-intestinal brush-border membranes of the mouse and piglet (Mazurier et al., 1985; Kawakami et al., 1990), on the amniotic membrane (Otsuki et al., 2000) and on the surface of cells (Legrand et al., 1992) such as hepatocytes, erythro-leukemic cells, monocytes, peritoneal macrophages, activated T-cells, platelets as listed by Brock (1997).
LF binding molecules have been characterized in many types of microorganisms. In Toxoplasma gondii, two LF binding proteins were recently identified as the ROP4 and ROP2 antigens (Dziadek et al., 2007). In viruses, interactions of LF with the V3-loop of gp120 and proteins E1-2 of the Human Immunodeficiency Virus (HIV) and the hepatitis C virus, respectively, have been proposed (Swart et al., 1996; Yi et al., 1997). Strong interactions of bovine LF with adenovirus polypeptides III and IIIa that bind to integrins of host cells have also been demonstrated (Pietrantoni et al., 2003).
Concerning bacteria, many studies reported LF binding to cells and its subsequent bactericidal effect. On Gram-positive Staphylococcus aureus and Streptococcus uberis, both glycosidic and proteic LF binding sites were evidenced but not further characterized (Naidu et al., 1992; Moshynskyy et al., 2003). In the case of Gram-negative bacteria, although evidence was provided that LF binds to the lipid-A moiety and/or the negative charges in the inner core of LPS with a high affinity (Appelmelk et al., 1994), it is unlikely that LF/LPS interactions occur when LPS is integrated in the cell wall of bacteria. Interestingly, it has been hypothesized that LF may use porins as anchoring sites on the surface of bacteria (Erdei et al., 1994; Sallmann et al., 1999).
Also, a number of gram-negative bacterial species (in the families Neisseriaceae and Moraxella) have surface receptors capable of specifically binding LF and biochemical and genetic evidence has confirmed the existence of two LF binding proteins, LbpA and LbpB (Schryvers et al., 1998; Prinz et al., 1999; Wong and Schryvers, 2003). A receptor on Trypanosma cruzi has also been observed (Lima et al., 1988). Unlike the mammalian receptors, the bacterial receptors are species-specific (Ling and Schryvers, 2006).
GLYCOSYLATION OF LACTOFERRIN
All Lfs contain biantennary N-acetyllactosamine-type glycans, α, 1-6 fucosylated on the N-acetylglucosamine residue linked to the polypeptide chain (Spik et al., 1988). Human LF may also possess additional poly-N-acetyllactosamine antennae that may be α, 1-3-fucosylated on N-acetylglucosamine residues, whereas the LF of other species contains additional high-mannose-type glycans (Coddeville et al., 1992). Both the number and location of the glycosylation sites vary among species. Furthermore, heterogeneity in the number of glycosylated sites is observed in individuals. The role of the glycan moiety seems to be restricted to a decrease in the immunogenicity of the protein and its protection from proteolysis (Spik et al., 1988; Van Veen et al., 2004).
The obtained data by Abdel-Salam et al. (2003) indicated that recombinant bLF (rbLF) from cell lysate contained more in the mannose-rich and less in the complex form, also, N-glycosylation may play an important role in the transport of rbLF in the presence of glycosidase inhibitors. Moreover, Abd El-Gawad et al. (2003) reported that rbLF was correctly folded and transported efficiently to the Golgi apparatus to become complex glycosylation and ultimately secreted into medium.
Impaired N-glycosylation may be played important role in the transport of recombinant bovine LF in the presence of the glycosides inhibitors (Abd El-Gawad et al., 2001b).
HEAT TREATMENTS EFFECTS ON LACTOFERRIN
The question of heat stability is very important when LF is used as a bioactive component of foods. Since LF is reported to be easily inactivated by heat treatment, it has been found that apo-LF is resistant to denaturation by heating (90-100°C/5min) at pH 4 and could pasteurize LF by heat treatment or sterilize it by a UHT method without any significant loss of biological properties (Abe et al., 1991). Heat denaturation data obtained by differential scanning calorimetry (Ruegg et al., 1977) and by fluorometry (Baer et al., 1979) have also been reported.
The thermal stability of bovine apo-LF and iron saturated LF have been investigated in relation to antibacterial activity and/or bacterial interaction. UHT treatment (135°C/4 sec) abolished the ability of iron-saturated LF to bind to bacteria as well as the bacteriostatic activity of apo-LF but standard pasteurization regimes used in dairy industry had practically no effect on LF structure. However a heat treatment at 137°C/8s do have only little effect on the ability of LF to stimulate cell proliferation. Kinetic approaches allowed Kussendrager (1994) to conclude that thermal stability of LF is affected by environmental conditions such as pH, salts and whey protein. Consequently the parameters of the heat-induced denaturation of LF have to be determined under conditions of the application of interest.
LF was estimated in camel’s milk from Kazakhstan, where two species of camels (Camelus bactrianus, Camelus dromedarius) and their hybrids cohabit. The concentrations of LF were determined according to three variation factors: region, season and species, the mean values in raw camel’s milk were 0.229±0.135 mg mL-1. The seasonal effect was the only significant variation factor observed with the highest values in the spring. The LF concentration varied in 1 week postpartum milk from 1.422 to 0.586 mg mL-1 (Konuspayeva et al., 2007).
BIOLOGICAL FUNCTIONS OF LACTOFERRIN
The interest in LF has been primarily in regard to potential capability of acting in iron transport and as an antimicrobial agent related to its iron chelating ability, thus depriving microorganisms of a source of iron. It has been shown to have a number of other physiological and biological functions (Shinmoto et al., 1992; Kussendrager, 1993; Brock, 1995; Lonnerdal and Iyer, 1995; Adamik and Walszczyk, 1996; Shinoda et al., 1996).
Although human and bovine LF differs, there is increasing evidence that both serve similar biological functions (Adamik and Walszczyk, 1996; Shinoda et al., 1996; Miyauchi et al., 1997).
Many roles have been proposed and continue to be proposed, for LF (Fig. 5). Although some of these are clearly related to its iron-binding properties, for example its ability to provide bacteria with a source of iron and therefore act as a promicrobial, others appear to be independent of iron binding. The antimicrobial activity of LF is well established. For many years this activity was attributed to the ability of LF to sequester iron thereby depriving potential pathogens of this essential nutrient.
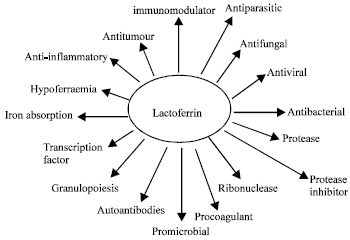 | |
| Fig. 5: | Proposed roles of lactoferrin by Brock (2002) |
LACTOFERRIN AND HOST DEFENSE
Legrand et al. (2005) revealed that the LF can both positively and negatively influence immune system cells and cells involved in the inflammation reaction, caused by its iron binding properties and interactions with target cells and molecules. In one way, LF may support the proliferation, differentiation and activation of immune system cells and strengthen the immune response. On the other side, LF acts as an anti-inflammatory factor. Also, LF may prevent the development of inflammation and subsequent tissue damage caused by the release of pro-inflammatory cytokines and reactive oxygen species.
Machnicki et al. (1993) and Haversen et al. (2002) noted that the protective effect of LF is manifested in a reduced production of some pro-inflammatory cytokines such as tumor necrosis factor (TNFα) or interleukins IL-1β and IL-6. Also, an increased amount of anti-inflammatory interleukin IL-10 has been reported in several cases. Iron is essential as a catalyst for the production of reactive oxygen species. Therefore, LF can diminish the harmful influence of reactive oxygen species produced by leukocytes at the sites of inflammation (Ward et al., 2005).
LACTOFERRIN AND IRON BINDING
Anderson et al. (1987) and Baker and Baker (2005) demonstrated that the iron binding site has the same composition and geometry in both lobes of LFs and TFs which comprises four protein ligands (2 Tyr, 1 Asp and 1 His) that provide three negative charges to balance the 3+ charge of Fe3+, together with the side chain of an Arg residue whose positive charge balances the negative charge of a CO32- anion.
In the "natural state" bovine LF is only partly saturated with iron (15-20%) and has a salmon pink colour. Iron-depleted LF with less than 5% iron saturation is called apo-LF, whereas iron-saturated LF is referred to as holo-LF. In breast milk the LF found is essentially apo-LF. The affinity of LF for iron is very high, about 260 times that of blood serum TF. The iron-binding capacity of LF is dependent of the presence of bicarbonate. The binding site appears to be optimized for the binding of ferric iron and bicarbonate but other cations may bound in the cleft; Al3+, Ga3+, Cu2+, Mn3+, Co3+ and Zn2+ etc. (Baker, 1994; Brodie et al., 1994). Also, Bagby and Bennett (1982) reported that the LF can bind metal ions other than iron including copper, zinc, aluminum, gallium, vanadium and calcium. Calcium ion-dependent oligomerizetion of LF has been reported, LF forms a tetramer in the presence of calcium ions (10 mM).
LF molecule can bind two Fe3+ ions, one within each lobe and the HCO3- ion is necessary for iron binding (Legrand et al., 1988; Spik et al., 1994). Normally, LF contains iron ions and the degree of iron saturation is 10 to 30% in milk. TF releases iron ions at pH 4 but LF holds iron ions at pH>2 and it is reported that the iron binding strength of LF is 260 folds stronger than that of TF (Aisen and Liebman, 1972). Also, Baker and Baker (2005) reported that the iron release depends on the destabilization of the closed form, in the absence of receptor binding (as is the case for TF) while Mazurier and Spik (1980) decided that the release is triggered by lowering the pH. It has been reported that the stability of binding or the strength of iron binding to LF varies depending on species, in the following order: human and equine LF>bovine LF>bovine TF (Shimazaki et al., 1993).
A lot of substances have been found to bind with LF including many kinds of small molecules and biopolymers (Brock, 1997), such as trypan blue dyes (Malmquist and Johansson, 1971), some kinds of drugs (Atkinson and Begg, 1988), ferritin (Pahud and Hilpert, 1976), immunoglobulins (Watanabe et al., 1984; Ena et al., 1990), albumin (Ena et al., 1990), β-Lg (Ena et al., 1990), DNA (Hutchens et al., 1989b), LPS (Elass-Rochard et al., 1995; Wang et al., 1995), lipids (Appelmelk et al., 1994), agar/agarose (Johansson and Hjerten, 1960a), carrier ampholytes (Shimakaki et al., 1991) and heparin (Zou et al., 1992; Van Berkel et al., 1997).
The iron content and iron saturation of different LFs of buffalo, cow (Local, Friesian and Brown-Swiss breeds), ewe (Rahmani and Ossemi breeds), goat as well as camel were markdleyblower in colostrum than in normal milk (Mahfouz et al., 1997).
ANTIMICROBIAL ACTIVITY
Kirkpatrick et al. (1971) observed that LF affects the growth and proliferation of a variety of infectious agents including gram-positive and negative bacteria, viruses, protozoa and fungi. Legrand et al. (2005) and Valenti and Antonini (2005) explained that the LF is considered to be a part of the innate immune system. It also takes part in specific immune reactions but in an indirect way which due to its strategic position on the mucosal surface. It represents one of the first defense systems against microbial agents invading the organism mostly via mucosal tissues.
It has been widely accepted for many years that LF displays antimicrobial activity against many different infectious agents. This activity was originally attributed to its ability, in common with TF, to sequester iron with a high affinity and unlike TF, retain its bound iron under acidic conditions.
LF inhibits the growth of many Gram-positive and -negative bacteria, some yeast and molds (Table 2) and some kinds of parasites. LF is known to act synergistically with lysozyme (Carlsson and Bjorck, 1987) and IgA (Akin et al., 1994) in various secretory fluids. The bacteriostatic effects of LF are thought to be due to its ability to sequester environmental iron (Arnold et al., 1980) because some kinds of bacteria, such as E. coli, secrete chelators to enhance iron uptake.
The antimicrobial activity of rbLF, its peptic digest LFcin-B recombinant glycosylated human lysozyme (rhLz) and mixture of LFcin-B+rhLz+EDTA against Staphylococcus aureus, Listeria monocytogenes, Escherichia coli, Salmonella typhimurium, Salmonella enteritidis, was investigated by El-Sayed et al. (2003).
Yekta et al. (2010) studied the ability of bovine and human lactoferrin, two natural antimicrobial proteins present in milk, to inhibit E. coli O157:H7 growth and attachment to a human epithelial colorectal adenocarcinoma cell line (Caco-2). The direct antibacterial effect of bLF on E. coli O157:H7 was stronger than that of hLF. Nevertheless, both lactoferrins had bacteriostatic effects even at high concentrations (10 mg mL-1), suggesting blocking of LF activity by a yet undefined bacterial defense mechanism.
| Table 2: | Lactoferrin-susceptible and -resistant microorganisms |
 | |
| a: Arnold et al. (1980), b: Ellison (1994) | |
Abdel-salam et al. (2006) shows that transformation of yeast cells with cDNA encoding bovine lactoferrin insert with eukaryotic expression vector occurred. Also, HPLC analysis data showed that the rbLf extracted from yeast media had the same retention time and molecular weight of the bovine lactoferrin standard. Recombinant lactoferrin extracted from medium demonstrated a greater inhibition effect than recombinant lactoferrin extracts from the cell lysates against Bacillus subtilis and Escherichia coli.
ANTIBACTERIAL ACTIVITY
The antibacterial activity of LF was initially ascribed to its ability to bind and sequester environmental iron, thereby depriving potential pathogens of this essential nutrient. The ability of LF to inhibit bacterial growth in vitro was indeed one of the earliest functions described for the protein. The antimicrobial activity of LF was demonstrated towards a number of bacteria and apo-LF was found to be bactericidal for Streptococcus mutans and Vibrio cholerae but not for Escherichia coli (Arnold et al., 1977). Further studies have shown that LF was bactericidal only when in its iron-free state and that iron-saturated LF has a reduced antimicrobial activity (Arnold et al., 1980; Kalmar and Arnold, 1988; Yamauchi et al., 1993). Iron-independent killing by LF was, however, first demonstrated by Arnold and co-workers in 1982. Previous study in 1981 had shown that LF was bactericidal for Streptococcus mutans even when exogenous iron was added to the experimental media (Arnold et al., 1981). Bortner et al. (1989) proposed that this iron-independent LF killing was a result of a direct interaction of LF with the bacterial surface. Lassiter et al. (1987) had previously suggested that the target site on bacteria would be anionic. Further studies have attempted to elucidate the mechanisms for this direct effect. LF was shown to interact with LPS of the Gram-negative bacterial membrane of E. coli (Appelmelk et al., 1994), with the release of the LPS from the membrane. However, this release was blocked by addition of Ca2+ and Mg2+ ions. The presence of Ca2+ ions also inhibited the ability of LF to increase the susceptibility of E. coli to the antibiotic rifampicin (Ellison et al., 1990). The importance of these divalent cations as modulators for the antimicrobial activity of LF has been further reported by Bortner et al. (1986, 1989) who showed that LF sensitive strain of Legionella pneumophila was protected from killing by addition of calcium chloride, magnesium nitrate and magnesium chloride, however addition of sodium chloride had no effect. Kalmar and Arnold (1988) showed that Mg2+ ions decreased LF killing, whereas the addition of Ca2+ or K+ ions had no affect on the antimicrobial activity of LF.
It has been discovered that LFcin, a cationic peptide generated by the pepsin digestion of LF, has more potent bactericidal activity than the native protein. There are two forms known at present: LFcin H (derived from human LF) and LFcin B (of bovine origin) (Bellamy et al., 1992a).
The proteolytic activity of LF is considered to inhibit the growth of some bacteria such as Shigella flexneri or enteropathogenic E.coli through degrading proteins necessary for colonization. However, this can be disabled by serine protease inhibitors (Orsi, 2004; Ward et al., 2005).
ANTIVIRAL ACTIVITY
LF has been shown to be effective in protecting against a number of different viruses (Marchetti et al., 1996; Shimizu et al., 1996).
Marchetti et al. (1996) reported that both human and bovine LF were effective against the herpes simplex virus type 1 (HSV-1) by inhibiting adsorption of the virus. This activity was independent of the iron with-holding, since both iron saturated and apo-LF was equally effective. Swart et al. (1998) found that LF from both human and bovine sources was able to completely block HCMV replications and to inhibit HIB-1 induced cytopathic effects.
Evidence suggests that LF contributes to the host defense against viral infections. It has been reported that LF is effective against Hepatitis C Virus (HCV) (Tanaka et al., 2000a; Yi et al., 1997), Herpes Simplex Virus (HSV) (Hammer et al., 2000), Feline Immunodeficiency Virus (FIV) (Sato et al., 1996), Human Immunodeficiency Virus (HIV)-l and human cytomegalovirus (Harmsen et al., 1995). With respect to the anti-HCV effect of LF, oral administration of bovine LF (1.8 to 3.6 g day-1) was found to result in a decrease in serum alanine transaminase and HCV RNA concentrations in 4 of 7 patients (Tanaka et al., 2000b). These effects are thought to be attributable to the interaction between LF and an HCV envelope protein (Tanaka et al., 2000b) or between the glycan chains of LF and HCV (Valenti et al., 1998).
The effects of recombinant Lf on the growth of human parainfluenza virus type 2 (hPIV-2) in LLCMK2 cells were investigated by Yamamoto et al. (2010), using a recombinant, green fluorescence protein-expressing hPIV-2 (rghPIV-2), it was found that virus entries into cells were considerably inhibited by Lf but cell-to-cell spread was not inhibited.
ANTIINFLAMMATORY PROPERTIES
LF has a beneficial effect on infections and protects animals against a lethal dose of LPS (Zagulski et al., 1989; Dial et al., 2005). Additionally, LF plays antiinflammatory roles in noninfectious pathologies such as rheumatoid arthritis, inflammatory bowel disorders, neurodegenerative diseases and skin allergies. It has been shown that administration of LF protects against chemically induced cutaneous inflammation (Cumberbatch et al., 2003) and no steroidal antiinflammatory drug-induced intestinal injury (Dial et al., 2005). In collagen-induced and septic arthritis mouse models, peri-articular injection of human LF reduced inflammation (Guillen et al., 2000).
ANTIOXIDANT ACTIVITY
Bannister et al. (1982) reported that the generation of hydroxyl radicals by the xanthine-xanthine oxidase reaction has been shown to be increased by iron-saturated LF. The antioxidant activity of LF has been attributed to its sequestration of free iron ions. Apo-LF sequesters any free iron in secretions, so it may protect mucus glycoproteins from active oxygen species generated in iron-catalyzed reactions (Clamp and Creeth, 1984). It is expected that such mucoprotective action would be overcome during infections. The suppression of lipid peroxidation has also been observed by Shinmoto et al. (1992).
ANTIPARASITIC ACTIVITY
LF acts against parasites in various ways. For example, the infectivity of Toxoplasma gondii and Eimeria stiedai sporozoites is reduced after their incubation with LF B. It is thought that LFcin breaches parasitic membrane integrity causing subsequent changes in interactions between the host and the parasite (Omata et al., 2001). The competition for iron between the parasite and LF is the basis of its antiparasitic activity against Pneumocystis carinii (Cirioni et al., 2000). In contrast, some parasites such as Tritrichomonas foetus are able to use LF as a donor of ferric ions (Tachezy et al., 1996).
ANTIFUNGAL ACTIVITY
LF was first reported to have anti-fungal activity by Kirkpatrick et al. (1971). In combination with fluconazole, LF was shown to reduce the minimum inhibitory concentration at which fluconazole killed a number of clinical isolates of Candida species, suggesting that LF may have a potential use in combination therapy against drug-resistant Candida infections (Kuipers et al., 1999). The synergistic action of LF with antibiotics, antifungals and antibacterial agents already in use increases their efficacy (Ellison and Giehl, 1991; Naidu and Arnold, 1994; Wakabayashi et al., 1996, 1998). The combined use of LF and antifungal against severe infections with Candida species is hence an attractive therapeutic option. Since fluconazole-resistant Candida species have been frequently reported in HIV-infected patients, the addition of LF which is one of the host specific defense factors present in saliva that exhibit antifungal activity, may delay the occurrence of resistant species (Kuipers et al., 1999). Al-Sheikh (2009) reported that lactoferrin showed significant antifungal effect on the three pathogenic Candida species viz., C. albicans, C. krusei and C. tropicalis while the addition of iron enhances the multiplication of Candida species.
ANTICANCER ACTIVITY
Administration of bovine LF has been found to be effective in reducing the number of Aden carcinomas in the large intestine chemically induced in rats (Tsuda et al., 2000) and in inhibition of tongue carcinogenesis in rat (Tanaka et al., 2000c) and in suppression of the spontaneous development of jejunely polyps in mice (Tsuda et al., 2000) and in inhibition of lung metastasis of colon carcinoma cells in mice (Tsuda et al., 2000). It has been proposed that these effects may be due to an increase in cytotoxic cells in peripheral blood.
Tsuda et al. (2002) and Mohan et al. (2006) reported that the concomitant administration of bovine LF and carcinogens to rodents inhibits the induction of activating enzymes for carcinogenic heterocyclic amines, modulates lipid per oxidation and activates antioxidant and carcinogen detoxification enzyme activities, blocking cancer development.
There is increasing evidence, based on animal studies, that LF may have therapeutic value in treatment of different types of cancer (Sekine et al., 1997; Uchida et al., 1999). LF has great potential therapeutic use in cancer disease prevention and/or treatment, namely as a chemo-preventive agent (Rodrigues et al., 2009).
CELL GROWTH PROMOTING ACTIVITY
LF is known to act as a negative feedback regulator of myelopoiesis (Broxmeyer et al., 1980) and as a growth factor for human lymphocytic cell lines in serum-free medium (Hashizume et al., 1983). It is reported that hLF has greater growth stimulatory activity than hTF (Hashizume et al., 1983). Also, LF shows promotion of nerve growth factor synthesis and secretion in mouse fibroblast L-M cells (Shinoda et al., 1993) and promotes endometrial cell proliferation (Yanaihara et al., 2000).
BONES REMODELING ACTIVITY
LF has been identified as a potent anabolic factor affecting osteocytes. It is stimulates osteoblast proliferation, enhances thymidine incorporation into osteocytes and reduces apoptosis of osteoblasts by 50-70%. A similar effect was also recorded in chondrocytes (Cornish et al., 2004). LF reduces or even inhibits osteoclas-togenesis in a concentration-dependent fashion. On the other hand, LF shows no influence on the bone resorption performed by mature osteoclasts (Lorget et al., 2002).
Cornish et al. (2004) showed that LF may affect bone cells through the inhibition of osteolytic cytokines such as TNFα or IL-1β, whose levels rise during inflammation. Thus, LF contributes to the stabilization of the osseous tissue. Because of these aforementioned properties, LF might be potentially useful in the treatment of diseases such as osteoporosis in the future.
ENZYMATIC ACTIVITY
LF has the ability to function as an enzyme in some reactions. LF is the milk protein with the highest levels of amylase, DNAse, RNAse and ATPase activities (Devi et al., 1994; Kanyshkova et al., 2003). However, these are not the only enzymatic activities of LF. The basis for LF various enzymatic activities is unknown. However, the variety of activities can be attributed to variations in the nature of the protein: multiple isoforms; degrees of glycosylation; tertiary structure (holo-or apo- LF) and the degree of oligomerisation. For instance, the LF molecule capable of hydrolysing RNA has an isoform that is incapable of Fe3+ binding (Furmanski et al., 1989).
LF has demonstrated remarkable resistance to proteolytic degradation by trypsin and trypsin-like enzymes. The level of resistance is proportional to the degree of iron saturation (Brock et al., 1976; Brines and Brock, 1983; Lyer and Lonnerdal, 1993). It is known that holo-LF shows greater resistance to protease digestion than apo-LF (Brock et al., 1976) and LF is not cleaved by plasmin or chymosin.
The discovery of LF enzymatic activities has helped to explain several of its physiological mechanisms, such as protection against microbial pathogens where LF might inhibit growth partly through hydrolysis of viral, bacterial, fungal and parasitic nucleic acids.
A remarkable similarity in some motifs between LF and ribonuclease A has been revealed and LF is, indeed, capable of RNA hydrolysis. The ribonuclease activity varies depending on the type of RNA. mRNA is the most sensitive to LF, whereas tRNA is the least. The non-iron-binding isoforms of LF seem to be responsible for RNA degradation (Furmanski et al., 1989; Devi et al., 1994).
ANTIMICROBIAL ACTIVITY OF LACTOFERRIN-DERIVED PEPTIDES
The hydrolysates produced by pepsin cleavage of bovine and human LF were found to contain a potent bactericidal peptide, named LFcin B and LFcin H (Bellamy et al., 1992a). The action of pepsin on LF produces peptides that have enhanced antimicrobial action as compared to LF (Jones et al., 1994; Wakabayashi et al., 1994; Facon, 1996; Facon and Skura, 1996; Dionysius and Milne, 1997; Tomita et al., 1998). A peptide with 28 residues from the N-terminal of LF has been names LFcin B and has much stronger antibiotic activity than LF (Jones et al., 1994). Multiple peptides have been isolated from pepsin hydrolysates of LF, with varying microbial activities (Dionysius and Milne, 1997) isolated 3 peptides with different structures and activities. Peptide I had essentially the same structure as LFcin B. The three peptides displayed varying antibacterial activity against a number of different spoilage and pathogenic organisms. Peptide I was most effective against Listeria monocytongenes. Antimicrobial activity shown in media is lost in food products and inhibited by 5 mM calcium and bile salts (Facon, 1996; Facon and Skura, 1996).
LFcin B has been shown also to be effective at concentrations as low as 3 μg L-1 against a number of different strains of yeasts and filamentous fungi (Bellamy et al., 1994).
CATIONIC PEPTIDES
A wide variety of organisms produce antimicrobial peptides as a primary innate immune strategy (Hancock and Lehrer, 1998). Hundreds of such peptides have been isolated throughout nature, from single celled microorganisms, mammals, amphibians, birds, fish and plants (Hancock and Chapple, 1999), indicating their importance in the innate immune system (Bevins, 1994; Hancock and Diamond, 2000). Typically, these peptides are relatively short (less than 100 amino acids), positively charged, amphiphilic and are reported to be active against bacteria, fungi, viruses and protozoa (Martin et al., 1995). They display a great structural diversity and although their modes of action can vary and are not fully understood, their main site of action is thought to be the cell membrane. Several mechanisms have been proposed for cationic antimicrobial peptides. Hydrophobicity, cationicity and secondary structure have been implicated in the antimicrobial effect. Although certain peptide structural groups have been noted, including amphipathic-helices, -structures, extended structures and loops, no overall conservation of amino acids exists (Boman, 1995; Hancock, 1997).
LACTOFERRICIN
The presence of stable antimicrobial peptides resulting from the proteolysis of LF was shown more than a decade ago when Saito et al. (1991) demonstrated that limited acid proteolysis of bovine LF yielded a hydrolysate that had greater antibacterial activity than LF. Bellamy et al. (1992a, b) described the generation, after pepsin hydrolysis, of fragments from human and bovine LFcin that showed enhanced antimicrobial activity but no iron-binding capacities. The fragments were characterized and named human and bovine LFcin. Both peptides are derived from the N-terminal region of the N-lobe and have greater antibacterial activity than their parent proteins LFcin H corresponds to amino acid residues 1-47 from the N-terminal region of the protein (Fig. 2) and includes an 18-residue loop formed by an internal disulphide bridge. Residues 1-11 constitute a separate fragment which remains bound to the main loop by a disulfide bridge. The slightly more potent LFcin B from bovine LF comprising only residues 17-41, consists primarily of the 18-residue loop stabilized by a disulfide bridge.
BOVINE LACTOFERRICIN
LFcin B, like other antimicrobial peptides that display membrane-disruptive properties, contains a high proportion of basic amino acid residues. It has been demonstrated that this highly cationic portion of LF is responsible for the ability of LF to bind glycosaminoglycan (Mann et al., 1994), heparin and LPS (Elass-Rochard et al., 1995). LFcin B is proposed to exert its effect at the surface of the bacterial membrane (Bellamy et al., 1993) and positive charges within the peptide are thought to aid interaction with membrane components. The greater the number of positive charges, the greater the number of interactions with negatively charged membrane components (Nikaido and Vaara, 1985; Hwang et al., 1998). Numerous studies have been undertaken to identify which regions of the bovine peptide are important for its antimicrobial activity (Hoek et al., 1997; Schibli et al., 1999).
HUMAN LACTOFERRICIN
Due to the limited supply of human milk and the difficulties in extracting the native peptide, less research has been undertaken on LFcin H. It is a 47 amino acid peptide produced under the same conditions as LFcin B though of lower antimicrobial potency. However, the larger concentration of LF observed in human milk, compared to the respective concentration in bovine milk, may make the antimicrobial activity of hLF and LFcin relevant in vivo. The peptide is similar to LFcin B in that it has a loop region held together by a disulphide bridge. It has been suggested that antimicrobial activity was independent of the presence of this bridge (Bellamy et al., 1992a, b). Although the total and net positive charges carried by the LFcin H are higher than those carried by LFcin B, the proportion of basic amino acid residues is greater in the bovine peptide. The lower cationicity within the 18 residue-loop might be at the heart of the difference in antimicrobial activity exhibited by human LF-derived peptides.
NUTRITIONAL AND APPLICATIONS OF LACTOFERRIN
Milk proteins have many nutritional, functional and biological functions in humans and animals and could also be used in practical applications. LF is one such milk protein (Hyvnen, 2010). The LF-derived peptide Lfcin is released through proteolysis by pepsin (Korhonen and Pihlanto, 2006), has received much attention recently due to its various roles in host defense (Gifford et al., 2005). bLF is a by-product of the dairy process and as such is readily available. rhLf can be expressed in milk, rice or microorganisms and is also commercially available. Both can be added to foods and different health products (Lonnerdal, 2009). The suggested applications for LF are for example from food preservatives to health-promoting foods and supplements, infant foods, iron supplements, paharmaceuticals, sport foods, nutritional foods, chewing gums, healthcare products, such as toothpaste, mouthwashes and cosmetics (Marnila and Korhonen, 2009). The Food and Drug Administration (FDA) of the United States has assessed LF and considers it to be generally safe (U.S. Food and Drug Administration, 2001). In the meat industry, LF can be used as spray applied to carcasses to decrease the growth of contaminant bacteria and to extend the shelf-life (Naidu, 2002). LF could replace food and feed additives as a preservative, because it is a natural protein of milk. LF has been used as a supplement in calf feed, to prevent neonatal diarrhea and to increase weight gain (Joslin et al., 2002; Robblee et al., 2003). Calves receiving supplemental bLF in colostrum and milk replacer had fewer days of diarrhea with less serious clinical signs than the control calves. LF can also stimulate carbohydrate absorption and increase small intestine epithelial cell size (Zhang et al., 2001), resulting a faster growth rate of mice.
Iron-saturated LF may serve a nutritional function as a source of iron or LF may regulate iron absorption. It is reported that the clearance of LF intravenously injected into mice is due to a hepatic receptor that specifically binds oligosaccharides containing Fuc (α-1, 3) GlcNAc linkages (Prieels et al., 1978). On the other hand, another mode of recognition of hLF which is avidly taken up by the mouse liver, has been proposed. It is suggested that the integrity of the protein moiety of LF is required for its effective uptake by the liver (Moguilevsky et al., 1985). The effectiveness of bLF in prevention of intestinal infections and in inhibition of bacterial translocation from the gut lumen into host tissues has been demonstrated in experiments using mice (Teraguchi et al., 1995).
There are two ways in which LF can be eliminated from the organism: either through receptor-mediated endocytosis of phagocytic cells (macrophages, monocytes and other cells belonging to the reticuloendothelial system) with subsequent iron transfer to ferritin or through direct uptake by the liver. Endocytosis performed by Kupffer cells, liver endothelial cells and hepatocytes contributes to LF removal (Levay and Viljoen, 1995).
The large-scale preparation of bLF from cheese whey or skim milk (up to 100 metric tones per year) and of recombinant hLF produced in microorganisms and plants makes LF available for human and animal (fish farming) health purposes and commercial applications. The first major application of bLF was the supplementation of infant formulas but it is now added to cosmetics, pet care supplements and immune system-enhancing nutraceuticals, including drinks, fermented milks and chewing gums. In all these media, LF is expected to exert its natural antimicrobial, antioxidative, antiinflammatory, anticancerous and immunomodulatory properties. Furthermore, clinical trials demonstrated the efficiency of LF against infections and in inflammatory diseases. For example, a recent clinical study concluded that the combination of LF and fluconazole at the threshold minimal inhibitory concentrations elicited potent synergism, leading to total fungistasis of C. albicans and C. glabrata vaginal pathogens (Naidu et al., 2004). Also, LF was reported as a potent molecule in the treatment of common inflammatory diseases (Legrand et al., 2005). In addition, extensive clinical trials are underway in Japan to further explore its preventive potential against colon carcinogenesis (Tsuda et al., 2002).
On the other hand, LF offers applications in food preservation and safety, either by retarding lipid oxidation (Medina et al., 2002) or by limiting the growth of microbes. For example, incorporation of LF into edible films has a great potential to enhance the safety of foods since the film can function as a physical barrier as well as an antimicrobial agent. LF can be also directly used as a spray applied to beef carcasses (Taylor et al., 2004).
Lastly, LF can be used as a clinical marker of inflammatory diseases since LF levels in blood and biological fluids may greatly increase in septicemia or during Severe Acute Respiratory Syndrome (Reghunathan et al., 2005). In the same way, fecal LF levels quickly increase with the influx of leukocytes into the intestinal lumen during inflammation. Fecal LF is thus used as a noninvasive diagnostic tool to evaluate the severity of intestinal inflammation in patients presenting with abdominal pain and diarrhea (Greenberg et al., 2002). This biomarker has been shown to be a sensitive and specific marker of disease activity in chronic inflammatory bowel disease (Kane et al., 2003) and in Crohn’s disease (Buderus et al., 2004).
Several evidences indicating benefits of bLF supplementation in infant formula were reported, including an improvement in intestinal microbialflora (Kawaguchi et al., 1989; Roberts et al., 1992), enhanced serum ferritin (Chierici et al., 1992) and hematocrit (King et al., 2007) levels and reduced lower respiratory tract illnesses (King et al., 2007). Addition of bLF also inhibits lipid oxidation of infant formula (Satue-Gracia et al., 2000). Currently, bLF containing infant formulas are sold in Indonesia and Korea, as well as in Japan. Other bLF containing products include yogurt, skim milk, milk-type drinks, supplemental tablets, pet food and cosmetics (Wakabayashi et al., 2006).The beneficial effects of these bLF containing products on the health have been proved in clinical and animal studies. The effect of yogurt on rotaviral gastroenteritis and the effect of tablets on chronic hepatitis C, rotaviral gastroenteritis and Helicobacter pylori gastric infection have been reported (Egashira et al., 2007; Ishii et al., 2003; Okuda et al., 2005). The therapeutic effect of pet food on dermatitis in dogs and cats was also shown (Masada et al., 1995).
Bovine colostrum can be freeze-dried and used as raw materials for human food supplements which participate to improve immunity of human bodies by raising the immunoglobulin and lactoferrin concentrations. The results led to the establishment of limits for the quality of colostrum (rich or poor in IgG), to develop a table of correspondence between IgG, lactoferrin and density that can be used as a practical method for rapid and cheap evaluation of colostrum (Bar et al., 2010).
Industry continues to attempt to improve formula with the addition of compounds such as fatty acids, oligosaccharides, nucleotides and lactoferrin. However, human milk has such far reaching effects on the infant`s immune response that normal development depends heavily on its provision. All mothers should be encouraged and supported to continue breastfeeding for six months and beyond in order to promote the good health of their infants (Oddy, 2002).
CONCLUSIONS
LF has been the focus of intense research of late. Due to its unique antimicrobial, immunomodulatory and even antineoplastic properties, LF seems to have great potential in practical medicine. Nevertheless, much research and many experiments still need to be carried out in order to obtain a better understanding of its activity and interactions and to enable the full and safe utilization of this glycoprotein.
REFERENCES
- Abd El-Gawad, I.A., E.M. El-Sayed, M.B. Mahfouz and A.M. Abd El-Salam, 1996. Changes of lactoferrin concentration in colostrum and milk from different species. Egypt. J. Dairy Sci., 24: 297-308.
Direct Link - Abd El-Gawad, I.A., E.M. El-Sayed, A.M. El-Salam, M.B. Mahfouz and H. Naim, 2001. N-Glycosylation, intracellular transport and sorting of recombinant bovine lactoferrin in polar and non polar epithelial cells. Proceeeings of the 8th Egyptian Conference of Dairy Science and Technology, Nov. 3-5, Cairo, Egypt, pp: 94-95.
- Abdel-Salam, A.M., R. Jacob, A.I.A. El-Gawad, E.M. El-Sayed, M.B. Mahfouz and H.Y. Naim, 2003. N-glycosylation, intracellular transport and sorting of recombinant bovine lactoferrin in polar and non-polar epithelial cells. Milichwissenschaft, 58: 359-363.
Direct Link - Abe, H., H. Saito, H. Miyakawa, Y. Tamura, S. Shimamura, E. Nagao and M. Tomita, 1991. Heat stability of bovine lactoferrin at acidic pH. J. Dairy Sci., 74: 65-71.
CrossRef - Adam, V., O. Zitka, P. Dolezal, L. Zeman and A. Horna et al., 2008. Lactoferrin isolation using monolithic column coupled with spectrometric or micro-amperometric detector. Sensors, 8: 464-487.
Direct Link - Adamik, B. and A. Walszczyk, 1996. Lactoferrin: Its role in defence against infection and immunotropic properties. Postepy Higieny Medycyny Doswiadczalnej, 50: 33-41.
PubMed - Aisen, P. and A. Liebman, 1972. Lactoferrin and transferrin: A comparative study. Biochimica et Biophysica Acta, 257: 314-323.
CrossRef - Akin, D.T., M.Q. Lu, S.J. Lu, S. Kendall, J. Rundegren and R.R. Arnold, 1994. Bactericidal activity of diiferent forms of lactoferrin. Adv. Exp. Med. Biol., 357: 61-70.
PubMed - Al-Sheikh, H., 2009. Effect of lactoferrin and iron on the growth of human pathogenic Candida species. Pak. J. Biol. Sci., 12: 91-94.
CrossRefPubMedDirect Link - Anderson, B.F., H.M. Baker, E.J. Dodson, G.E. Norris, S.V. Rumball, J.M. Waters and E.N. Baker, 1987. Structure of human lactoferrin at 3.2-A resolution. Proc. Natl. Acad. Sci., 84: 1769-1773.
PubMed - Appelmelk, B.J., Y.Q. An, M. Geerts, B.G. Thijs and H.A. De-Boer et al., 1994. Lactoferrin is a lipid A-binding protein. Infect. Immun., 62: 2628-2632.
Direct Link - Arnold, R.R., M. Brewer and J.J. Gauthier, 1980. Bactericidal activity of human lactoferrin: Sensitivity of a variety of microorganisms. Infect. Immun., 28: 893-898.
PubMed - Arnold, R.R., M.F. Cole and J.R. Mcghee, 1977. A bactericidal effect for human lactoferrin. Science, 197: 263-265.
CrossRef - Arnold, R.R., J.E. Russell, W.J. Champion and J.J. Gautheir, 1981. Bactericidal activity of human lactoferrin: Influence of physical conditions and metabolic state of the target microorganism. Infect. Immun., 32: 655-660.
Direct Link - Artym, J. and M. Zimecki, 2005. The role of lactoferrin in the proper development of newborns. Postepy Higienyi Medycyny Doswiadczalnej, 59: 421-432.
PubMed - Atkinson, H.C. and E.J. Begg, 1988. The binding of drugs to major human milk whey proteins. Bri. J. Clin. Pharma., 26: 107-109.
PubMed - Baer, A., M. Oroz and B. Blanc, 1979. Serological and fluorescene studies of the heat stability of bovine lactoferrine. J. Dairy Res., 46: 83-93.
CrossRef - Bagby, G.C. and R.M. Bennett, 1982. Feedback regulation of granulopoiesis: Polymerization of lactoferrin abrogates its ability to inhibit CSA production. Blood, 60: 108-112.
PubMed - Baker, E.N., 1994. Structure and reactivity of transferrins. Adv. Inorganic Chem., 41: 389-463.
CrossRef - Baker, E.N. and H.M. Baker, 2005. Molecular structure, binding properties and dynamics of lactoferrin. Cell. Mol. Life Sci., 62: 2531-2539.
CrossRefPubMedDirect Link - Bannister, J.V., W.H. Bannister, H.A. Hill and P.J. Thornalley, 1982. Enhanced production of hydroxyl radicals by the xanthine-xanthine oxidase reaction in the presence of lactoferrin. Biochimica et Biophysica Acta, 715: 116-120.
PubMed - Bar, E., I. Tiris, M. Miron, M. Bojita and G. Mihai, 2010. The relationship between the content of Immunoglobulin (IgG), lactoferrin and density in bovine colostrum-comparative study. Bull. UASVM Anim. Sci. Biotechnol., 67: 70-74.
Direct Link - Baveye, S., E. Elass, J. Mazurier, G. Spik and D. Legrand, 1999. Lactoferrin: A multifunctional glycoprotein involved in the modulation of the inflammatory process. Clin. Chem. Lab. Med., 37: 281-286.
PubMed - Bellamy, W., M. Takase, H. Wakabayashi, K. Kawase and M. Tomita, 1992. Antibacterial spectrum of lactoferricin-B, a potent bactericidal peptide derived from the N-terminal region of bovine lactoferrin. J. Applied Bacteriol., 73: 472-479.
PubMed - Bellamy, W., M. Takase, K. Yamauchi, H. Wakabayashi, K. Kawase and M. Tomita, 1992. Identification of the bactericidal domain of lactoferrin. Biochimica et Biophysica Acta, 1121: 130-136.
CrossRef - Bellamy, W., K. Yamauchi, H. Wakabayashi, N. Takakura, S. Shimamura and M. Tomita, 1994. Antifungal properties of lactoferricin B, a peptide derived from the N-terminal region of lactoferrin. Lett. Applied Microbiol., 18: 230-233.
Direct Link - Bellamy, W.R., H. Wakabayashi, M. Takase, S. Shimamura and M. Tomita, 1993. Role of cell-binding in the antibacterial mechanism of lactoferricin B. J. Applied Microbiol., 75: 478-484.
Direct Link - Bevins, C.L., 1994. Antimicrobial peptides as agents of mucosal immunity. Ciba Foundation Symp., 186: 250-260.
PubMed - Bezwoda, W.R. and N. Mansoor, 1989. Lactoferrin from human breast milk and from neutrophil granulocytes. Comparative studies of isolation, quantitation, characterization and iron biding properties. Biomed. Chromatogr., 3: 121-126.
PubMed - Bezwoda, W.R. and N. Mansoor, 1986. Isolation and characterisation of lactoferrin separated from human whey by adsorption chromatography using Cibacron Blue F3G-A linked affinity adsorbent. Clin. Chem. Acta, 157: 89-93.
CrossRef - Blackberg, L. and O. Hernell, 1980. Isolation of lactoferrin from human whey by a single chromatographic step. Federation Eur. Biochem. Soc. Lett., 109: 180-183.
PubMed - Boman, H.G., 1995. Peptide antibiotics and their role in innate immunity. Annu. Rev. Immunol., 13: 61-92.
CrossRef - Bortner, C.A., R.R. Arnold and R.D. Miller, 1989. Bactericidal effect of lactoferrin on Legionella pneumophilu: Effect of the physiological state of the organism. Can. J. Microbiol., 35: 1048-1051.
PubMed - Bortner, C.A., R.D. Miller and R.R. Arnold, 1986. Bactericidal effect of lactofemn on Legionella pneumophilu. Infect. Immun., 51: 373-377.
PubMed - Brines, R.D. and J.H. Brock, 1983. The effect of trypsin and chymotrypsin on the in vitro antibacterial and iron-binding proprieties of lactoferrin in human milk and bovine colostrum. Unusual resistance of human apo-lactoferrin to proteolytic digestion. Biochimica et Biophysica Acta, 759: 229-235.
PubMed - Brock, J., 1995. Lactoferrin: A multifunctional immunoregulatory protein. Immunol. Todays, 16: 417-419.
CrossRef - Brock, J.H., F. Arzabe, F. Lampreave and A. Pineiro, 1976. The effect of trypsin on bovine transferrin and lactoferrin. Biochimica et Biophysica Acta, 446: 214-225.
CrossRef - Brodie, A.M., E.W. Ainscough, E.N. Baker, H.M. Baker, M.S. Shongwe and C.A. Smith, 1994. Synergism and substitution in the lactofenins. Adv. Exp. Med. Biol., 357: 33-44.
PubMed - Broxmeyer, H.E., M. De-Sousa, A. Smithyman, P. Ralph, J. Hamilton, J.I. Kurland and J. Bognacki, 1980. Specificity and modulation of the action of lactoferrin, a negative feedback regulator of myelopoiesis. Blood, 55: 324-333.
PubMed - Buderus, S., J. Boone, D. Lyerly and M.J. Lentze, 2004. Fecal lactoferrin: A new parameter to monitor infliximab therapy. Digestive Dis. Sci., 49: 1036-1039.
CrossRef - Carlsson, A. and L. Bjorck, 1987. The effect of some indigenous antibacterial factors in milk on the growth of Bacillus stearothermophilus var. calidolactis. Milchwissenschaft, 42: 282-285.
Direct Link - Mohan, C.K.V., R. Kumaraguruparan, D. Prathiba and S. Nagini, 2006. Modulation of xenobiotic-metabolizing enzymes and redox status during chemoprevention of hamster buccal carcinogenesis by bovine lactoferrin. Nutrition, 22: 940-946.
CrossRef - Cheng, J.B., J.Q. Wang, D.P. Bu, G.L. Liu and C.G. Zhang et al., 2008. Factors affecting the lactoferrin concentration in bovine milk. J. Dairy Sci., 91: 970-976.
CrossRef - Chierici, R., G. Sawatzki, L. Tamisari, S. Volpato and V. Vigi, 1992. Supplementation of an adapted formula with bovine lactoferrin: 2. Effects on serum iron, ferritin and zinc levels. Acta Pediatrics, 81: 475-479.
PubMed - Cirioni, O., A. Giacometti, F. Barchiesi and G. Scalise, 2000. Inhibition of growth of Pneumocystis carinii by lactoferrins alone and in combination with pyrimethamine, clarithromycin and minocycline. J. Antimicro. Chemother., 46: 577-582.
PubMed - Clamp, J.R. and J.M. Creeth, 1984. Some non-mucin components of mucus and their possible biological roles. Ciba Foundation Symp., 109: 121-136.
PubMed - Coddeville, B., G. Strecker, J.M. Wieruszeski, J.F. Vliegenthart and H. Van-Halbeek et al., 1992. Heterogeneity of bovine lactotransferrin glycans. Characterization of α-D-Galp-(1->3)- β-D-Gal- and α-NeuAc-(2->6)- β-D-GalpNAc-(1->4)- β-D-GlcNAc-substituted N-linked glycans.Heterogeneity of bovine lactotransferrin glycans. Characterization of α-D-Galp-(1->3)- β-D-Gal- and α-NeuAc-(2->6)- β-D-GalpNAc-(1->4)- β-D-GlcNAc-substituted N-linked glycans. Carbohydrate Res., 236: 145-164.
PubMed - Cornish, J., K.E. Callon, D. Naot, K.P. Palmano and T. Banovic et al., 2004. Lactoferrin is a potent regulator of bone cell activity and increases bone formation in vivo. Endocrinology, 145: 4366-4374.
CrossRefDirect Link - Cumberbatch, M., M. Bhushan, R.J. Dearman, I. Kimber and C.E. Griffiths, 2003. IL-1b-induced Langerhans` cell migration and TNF-a production in human skin: Regulation by lactoferrin. Clin. Exp. Immunol., 132: 352-359.
PubMed - Devi, A.S., M.R. Das and M.W. Pandit, 1994. Lactoferrin contains structural motifs of ribonuclease. Biochimica et Biophysica Acta, 1205: 275-281.
CrossRef - Dial, E.J., A.J. Dohrman, J.J. Romero and L.M. Lichtenberger, 2005. Recombinant human lactoferrin prevents NSAID-induced intestinal bleeding in rodents. J. Pharmacy Pharm., 57: 93-99.
PubMed - Dionysius, D.A. and J.M. Milne, 1997. Antibacterial peptides of bovine lactoferrin: Purification and characterization. J. Dairy Sci., 80: 667-674.
CrossRef - Dziadek, B., J. Dziadek and H. Dlugonska, 2007. Identification of Toxoplasma gondii proteins binding human lactoferrin: A new aspect of rhoptry proteins function. Exp. Parasitol., 115: 277-282.
CrossRef - Egashira, M., T. Takayanagi, M. Moriuchi and H. Moriuchi, 2007. Does daily intake of bovine lactoferrin containing products ameliorate rotaviral gastroenteritis?. Acta Paediatrics, 96: 1238-1244.
PubMed - Elagamy, E.I., R. Ruppanner, A. Ismail, C.P. Champagne and R. Assaf, 1996. Purification and characterization of lactoferrin, lactoperoxidase, lysozyme and immunoglobulins from camel`s milk. Int. Dairy J., 6: 129-145.
CrossRefDirect Link - Elass-Rochard, E., A. Roseanu, D. Legrand, M. Trif and V. Salmon et al., 1995. Lactoferrin-lipopolysaccharide interaction: Involvement of the 28-34 loop region of human lactoferrin in the high-affinity binding to Escherichia coli 055B5 lipopolysaccharide. Biochem. J., 312: 839-845.
Direct Link - Ellison, R.T. III, 1994. The effects of lactoferrin on gram-negative bacteria. Adv. Exp. Med. Biol., 357: 71-90.
PubMed - Ellison, R.T. and T.J. Giehl, 1991. Killing of gram-negative bacteria by lactoferrin and lysozyme. J. Clin. Invest., 88: 1080-1091.
CrossRefDirect Link - El-Sayed, E.M., M.B. Mahfouz, A.M. Abdel-Salam, I.A. Abd El-Gawad and H.Y. Naim, 2003. Antimicrobial activity of recombinant bovine lactoferrin and lactoferricin B and glycosylatioed human lysozyme. Milichwissenschaft, 58: 266-270.
Direct Link - Ena, J.M., H. Castillo, L. Sanchez and M. Calvo, 1990. Isolation of human lactoferrin by affinity chromatography using insolubilized bovine β-lactoglobulin. J. Chromatogr. Biomed. Sci. Appl., 525: 442-446.
PubMed - Erdei, J., A. Forsgren and A.S. Naidu, 1994. Lactoferrin binds to porins OmpF and OmpC in Escherichia coli. Infect. Immunity 62: 1236-1240.
PubMed - Facon, M.J. and B.J. Skura, 1996. Antibacterial activity of lactoferricin, lysozyme and EDTA against Salmonella enteritidis. Int. Dairy J., 6: 303-313.
CrossRef - Farnaud, S. and R.W. Evans, 2003. Lactoferrin-A multifunctional protein with antimicrobial properties. Mol. Immunol., 40: 395-405.
CrossRef - Furmanski, P., Z.P. Li, M.B. Fortuna, C.V.B. Swamy and M.R. Das, 1989. Multiple molecular forms of human lactoferrin. Identification of a class of lactoferrins that possess ribonuclease activity and lack iron-binding capacity. J. Exp. Med., 170: 415-429.
Direct Link - Gifford, J.L., H.N. Hunter and H.J. Vogel, 2005. Lactoferricin: A lactoferrin-derived peptide with antimicrobial, antiviral, antitumor and immunological properties. Cell. Mol. Life Sci., 62: 2588-2598.
CrossRef - Goldman, A.S., C. Garza, R.J. Schanler and R.M. Goldblum, 1990. Molecular forms of lactoferrin in stool and urine from infants fed human milk. Pediatrics Res., 27: 252-255.
PubMed - Greenberg, D.E., Z.D. Jiang, R. Steffen, M.P. Verenker and H.L. DuPont, 2002. Markers of inflammation in bacterial diarrhea among travelers, with a focus on enteroaggregative Escherichia coli pathogenicity. J. Infect. Dis., 185: 944-949.
PubMed - Groves, M.L., 1960. The isolation of a red protein from milk. J. Am. Chem. Soc., 82: 3345-3350.
CrossRef - Guerrant, R.L., V. Araujo, E. Soares, K. Kotloff, A.A. Lima, W.H. Cooper and A.G. Lee, 1992. Measurement of fecal lactoferrin as a marker of fecal leucocytes. J. Clin. Microbiol., 30: 1238-1242.
Direct Link - Guillen, C., I.B. McInnes, D. Vaughan, A.B. Speekenbrink and J.H. Brock, 2000. The effects of local administration of lactoferrin on inflammation in murine autoimmune and infectious arthritis. Arthritis Rheumatism, 43: 2073-2080.
PubMed - Hagiwara, S., K. Kawai, A. Anri and H. Nagahata, 2003. Lactoferrin concentrations in milk from normal and subclinical mastitic cows. J. Vet. Med. Sci., 65: 319-323.
Direct Link - Hancock, R.E.W. and D.S. Chapple, 1999. Antibiotic peptides. Antimicrobial Agents Chemother., 43: 1317-1323.
Direct Link - Hancock, R.E.W. and R. Lehrer, 1998. Cationic peptides: A new source of antibiotics. Trends Biotechnol., 16: 82-88.
CrossRef - Harmon, R.J., F.L. Schanbacher, L.C. Ferguson and K.L. Smith, 1975. Concentration of lactoferrin in milk of normal lactating cows and changes occurring during mastitis. Am. J. Vet. Res., 36: 1001-1007.
PubMed - Harmsen, M.C., P.J. Swart, M.P. De-Bethune, R. Pauwels, E. De-Clercq, T.H. The and D.K. Meijer, 1995. Antiviral effects of plasma and milk proteins: Lactoferrin shows potent activity against both human immunodeficiency virus and human cytomegalovirus replication in vitro. J. Infect. Dis., 172: 380-388.
PubMed - Hashizume, S., K. Kuroda and H. Murakami, 1983. Identification of lactoferrin as an essential growth factor for human lymphocytic cell lines in serum-free medium. Biochim. Biophys. Acta, 763: 377-382.
PubMed - Hoek, K.S., J.M. Milne, P.A. Grieve, D.A. Dionysius and R. Smith, 1997. Antibacterial activity in bovine lactoferrin-derived peptides. Antimicrob. Agents Chemother., 41: 54-59.
PubMedDirect Link - Hutchens, T.W, J.F. Henry and T.T. Yip, 1989. Purification and characterization of intact lactoferrin found in the urine of human milk-fed preterm infants. Clin. Chem., 35: 1928-1933.
PubMed - Hutchens, T.W., J.F. Henry, T.T. Yip, D.L. Hachey, R.J. Schanler, K.J. Motil and C. Garza, 1991. Origin of intact lactoferrin and its DNA-binding fragments found in the urine of human milk-fed preterm infants. Evaluation by stable isotopic enrichment. Pediatrics Res., 29: 243-250.
PubMed - Hutchens, T.W., J.S. Magnuson and T.T. Yip, 1989. Rapid purification of porcine colostral whey lactoferrin by affinity chromatography on single-stranded DNA-agarose, characterization, amino acid composition and N-terminal amino acid sequence. Biochim. Biophys. Acta, Protein Struct. Mol. Enzymol., 999: 323-329.
CrossRefPubMed - Hwang, P.M., N. Zhou, X. Shan, C.H. Arrowsmith and H.J. Vogel, 1998. Three-dimensional solution structure of lactoferricin B, an antimicrobial peptide derived from bovine lactoferricin. Biochemistry, 37: 4288-4298.
Direct Link - Indyk, H.E. and E.L. Filonzi, 2005. Determination of lactoferrin in bovine milk, colostrum and infant formulas by optical biosensor analysis. Int. Dairy J., 15: 429-438.
CrossRef - Ishii, K., N. Takamura, M. Shinohara, N. Wakui and H. Shin et al., 2003. Long-term follow-up of chronic hepatitis C patients treated with oral lactoferrin for 12 months. Hepatology Res., 25: 226-233.
PubMed - Lyer, S. and B. Lonnerdal, 1993. Lactoferrin, lactoferrin receptors and iron metabolism. Eur. J. Clin. Nutr., 47: 232-241.
PubMed - Jones, E.M., A. Smart, G. Bloomberg, L. Burgess and M.R. Millar, 1994. Lactoferricin, a new antimicrobial peptide. J. Applied Microbiol., 77: 208-214.
CrossRef - Joslin, R.S., P.S. Erickson, H.M. Santoro, N.L. Whitehouse, C.G. Schwab and J.J. Rejman, 2002. Lactoferrin supplementation to dairy calves. J. Dairy Sci., 85: 1237-1242.
Direct Link - Kalmar, I.R. and R.R. Arnold, 1988. Killing of actinobacillus actinomycetemcomitans by human lactoferrin. Infec. Immun., 56: 2552-2557.
PubMed - Kane, S.V., W.J. Sandborn, P.A. Rufo, A. Zholudev and J. Boone et al., 2003. Fecal lactoferrin is a sensitive and specific marker in identifying intestinal inflammation. Am. J. Gastroenter., 98: 1309-1314.
PubMed - Kanyshkova, T.G., S.E. Babina, D.V. Semenov, N. Isaeva and A.V. Valssov et al., 2003. Multiple enzymic activities of human milk lactoferrin. Eur. J. Biochem., 270: 3353-3361.
CrossRefPubMedDirect Link - Kawakami, H., S. Dosako and B. Lonnerdal, 1990. Iron up-take from transferrin and lactoferrin by rat intestinal brush-border membrane vesicles. Am. J. Physiol., 258: 535-541.
PubMedDirect Link - Kijlstra, A., S.H. Jeurissen and K.M. Koning, 1983. Lactoferrin levels in normal human tears. Br. J. Ophthalmology, 67: 199-202.
PubMed - Konuspayeva, G., B. Faye, G. Loiseau and D. Levieux, 2007. Lactoferrin and immunoglobulin contents in camel`s milk (Camelus bactrianus, Camelus dromedarius and hybrids) from Kazakhstan. J. Dairy Sci., 90: 38-46.
Direct Link - Korhonen, H. and A. Pihlanto, 2006. Bioactive peptides: Production and functionality. Int. Dairy J., 16: 945-960.
CrossRefDirect Link - Kuipers, M.E., H.G. de Vries, M.C. Eikelboom, D.K.F. Meijer and P.J. Swart, 1999. Synergistic fungistatic effects of lactoferrin in combination with antifungal drugs against clinical Candida isolates. Antimicrob. Agents Chemother., 43: 2635-2641.
PubMed - Machnicki, M., M. Zimecki and T. Zagulski, 1993. Lactoferrin regulates the release of tumour necrosis factor alpha and interleukin 6 in vivo. Int. J. Exp. Patho., 74: 433-439.
PubMed - Legrand, D., J. Mazurier, A. Elass, E. Rochard and G. Vergoten et al., 1992. Molecular interactions between human lactotransferrin and the phytohemagglutinin-activated human lymphocyte lactotransferrin receptor lie in two loop-containing regions of the N-terminal domain I of human lactotransferrin. Biochemisrty, 31: 9243-9251.
PubMed - Legrand, D., J. Mazurier, M.H. Metz-Boutigue, J. Jolles, P. Jolles, J. Montreuil and G. Spik, 1984. Characterization and localization of an iron-binding 18-kDa glycopeptide isolated from the N-terminal half of human lactotransferrin. Biochim. Biophys. Acta, Protein Struct. Mol. Enzymol., 787: 90-96.
CrossRefPubMed - Legrand, D., J. Mazurier, J. Montreuil and G. Spik, 1988. Structure and spatial conformation of the iron-binding sites of transferring. Biochimie, 70: 1185-1195.
CrossRef - Levieux, D., F. Morgan, N. Geneix, I. Masle and F. Bouvier, 2002. Caprine immunoglobulin G, β-lactoglobulin, α-lactoalbumin and serum albumin in colostrum and milk during the early post partum period. J. Dairy Res., 69: 391-399.
PubMed - Lima, M.F., L.A. Beltz and F. Kierszenbaum, 1988. Trypanosoma cruzi: A specific surface marker for the amastigote form. J. Eukaryotic Microbiol., 35: 108-110.
CrossRef - Ling, J.M. and A.B. Schryvers, 2006. Perspectives on interactions between lactoferrin and bacteria. Biochem. Cell Biol., 84: 275-281.
PubMedDirect Link - Lonnerdal, B., 2009. Nutritional roles of lactoferrin. Curr. Opin. Clin. Nutr. Metab. Care, 12: 293-297.
PubMed - Lonnerdal, B. and S. Iyer, 1995. Lactoferrin: Molecular structure and biological function. Annu. Rev. Nutr., 75: 93-110.
CrossRef - Lorget, F., J. Clough, M. Oliveira, M.C. Daury, A. Sabokbar and E. Offord, 2002. Lactoferrin reduces in vitro osteoclast differentiation and resorbing activity. Biochem. Biophys. Res. Commun., 296: 261-266.
CrossRef - Mahfouz, M.B., E.M. El-Sayed, I.A.A. El-Gawad, H. El-Etriby and A.M.A. El-Salam, 1997. Structural studies on colostrum and milk lactoferrins from different species. Egyptian J. Dairy Sci., 25: 41-53.
Direct Link - Malmquist, J., N.E. Hansen and H. Karle, 1978. Lactoferrin in haematology. Scand. J. Haemato., 21: 5-8.
CrossRef - Malmquist, J. and B.G. Johansson, 1971. Interaction of lactoferrin with agar gels and with trypan blue. Biochim. Biophys. Acta, Protein Struct., 236: 38-46.
CrossRef - Mann, D.M., E. Romm and M. Migliorini, 1994. Delineation of the glycosaminoglycan-binding site in the human inflammatory response protein lactoferrin. J. Biology Chem., 269: 23661-23667.
PubMed - Martin, E., T. Gantz and R. Lehrer, 1995. Defensins and other endogenous peptide antibiotics of vertebrates. J. Leukocytes Biol., 58: 128-136.
PubMedDirect Link - Martins, C.A., M.G. Fonteles, L.J. Barrett and R.L. Guerrant, 1995. Correlation of lactoferrin with neutrophilic inflammation in body fluids. Clin. Diagn. Lab. Immuno., 2: 763-765.
PubMedDirect Link - Masson, P.L., J.F. Heremans and C.H. Dive, 1966. An iron-binding protein common to many external secretions. Clin. Chemis. Acta, 14: 735-739.
CrossRef - Medina, I., I. Tombo, M.T. Satue-Gracia, J.B. German and E.N. Frankel, 2002. Effects of natural phenolic compounds on the antioxidant activity of lactoferrin in liposomes and oil-in-water emulsions. J. Agric. Food Chem., 50: 2392-2399.
CrossRef - Moguilevsky, N., P.L. Masson and P.J. Courtory, 1987. Lactoferrin uptake and iron processing into macrophages: A study in familial haemochromatosis. Br. J. Haemat., 66: 129-136.
CrossRef - Moguilevsky, N., L.A. Retegui and P.L. Masson, 1985. Comparison of human lactoferrins from milk and neutrophilic leucocytes. Relative molecular mass, isoelectric point, iron-binding properties and uptake by the liver. Biochem. J., 229: 353-359.
PubMed - Montreuil, J., J. Tonnelat and S. Mullet, 1960. Preparation and properties of lactosiderophilin (lactotransferrin) of human milk. Biochim. Biophys. Acta, 45: 413-421.
PubMed - Moore, S.A., B.F. Anderson, C.R. Groom, M. Haridas and E.N. Baker, 1997. Three-dimensional structure of diferric bovine lactoferrin at 208A resoluation. J. Mol. Biol., 274: 222-236.
CrossRef - Moshynskyy, I., M. Jiang, M.C. Fontaine, J. Perez-Casal, L.A. Babiuk and A.A. Potter, 2003. Characterization of a bovine lactoferrin binding protein of Streptococcus uberis. Microb. Pathogenesis, 35: 203-215.
CrossRef - Nagasawa, T., I. Kiyosawa and K. Kuwahara, 1972. Amounts of lactoferrin in human colostrum and milk. J. Dairy Sci., 55: 1651-1659.
CrossRef - Naidu, A.S., M. Andersson and A. Forsgren, 1992. Identification of a human lactoferrin-binding protein in Staphylococcus aureus. J. Med. Microbio., 36: 177-183.
PubMed - Naidu, A.S., J. Chen, C. Martinez, J. Tulpinski, B.K. Pal and R.S. Fowler, 2004. Activated lactoferrin`s ability to inhibit Candida growth and block yeast adhesion to the vaginal epithelial monolayer. J. Reprod. Med., 49: 859-866.
PubMed - Naidu, S.A., 2002. Activated lactoferrin-A new approach to meat safety. Food Tech., 56: 40-45.
Direct Link - Nikaido, H. and M. Vaara, 1985. Molecular basis of bacterial outer membrane permeability. Microbiol. Rev., 49: 1-32.
PubMed - Oddy, W.H., 2002. Why breast milk has health benefits for infants and children: A review. Pak. J. Nutr., 1: 106-118.
CrossRefDirect Link - Oh, S.M., C.W. Pyo, Y. Kim and S.Y. Choi, 2004. Neutrophil lactoferrin upregulates the human p53 gene through induction of NF-kappaB activation cascade. Oncogene, 23: 8282-8291.
PubMed - Pietrantoni, A., A.M. Di Biase, A. Tinari, M. Marchetti, P. Valenti, L. Seganti and F. Superti, 2003. Bovine lactoferrin inhibits adenovirus infection by interacting with viral structural polypeptides. Antimicro. Agents Chemother., 47: 2688-2691.
CrossRef - Prieels, J.P., S.V. Pizzo, L.R. Glasgow, J.C. Paulson and R.L. Hill, 1978. Hepatic receptor that specifically binds oligosaccharides containing fucosyl α1 leads to 3 N-acetylglucosamine linkages. Proc. Natl. Acad. Sci. USA., 75: 2215-2219.
PubMed - Prinz, T., M. Meyer, A. Pettersson and J. Tommassen, 1999. Structural characterization of the lactoferrin receptor from Neisseria meningitidis. J. Bacterio., 181: 4417-4419.
PubMedDirect Link - Rado, T.A., J.S. Bollekens, G. Laurent, L. Parker and E.J. Benz, Jr., 1984. Lactoferrin biosynthesis during granulocytopoiesis. Blood, 64: 1103-1109.
PubMed - Rey, M.W., S.L. Woloshuk, H.A. deBoer and F.R. Pieper, 1990. Complete nucleotide sequence of human mammary gland lactoferrin. Nucleic Acids Res., 18: 5288-5288.
PubMed - Robblee, E.D., P.S. Erickson, N.L. Whitehouse, A.M. McLaughlin, C.G. Schwab, J.J. Rejman and R.E. Rompala, 2003. Supplemental lactoferrin improves health and growth of holstein calves during the preweaning phase. J. Dairy Sci., 86: 1458-1464.
Direct Link - Roberts, A.K., R. Chierici, G. Sawatzki, M.J. Hill, S. Volpato and V. Vigi, 1992. Supplementation of an adapted formula with bovine lactoferrin: 1. Effect on the infant faecal flora. Acta Paediatrics, 81: 119-124.
PubMed - Rodrigues, L., J. Teixeira, F. Schmitt, M. Paulsson and H.L. Mansson, 2008. Lactoferrin and cancer disease prevention. Crit. Rev. Food Sci. Nutr., 49: 203-217.
CrossRefPubMedDirect Link - Ruegg, M., U. Moor and B. Blanc, 1977. A calorimetric study of the thermal denaturation of whey proteins in simulated milk ultrafiltrate. J. Dairy Res., 44: 509-520.
CrossRef - Sallmann, F.R., S. Baveye-Descamps, F. Pattus, V. Salmon, N. Branza, G. Spik and D. Legrand, 1999. Porins OmpC and PhoE of Escherichia coli as specific cell-surface targets of human lactoferrin. Binding characteristics and biological effects. J. Biol. Chem., 274: 16107-16114.
PubMedDirect Link - Sato, R., O. Inanami, Y. Tanaka, M. Takase and Y. Naito, 1996. Oral administration of bovine lactoferrin for treatment of intractable stomatitis in Feline Immunodeficiency Virus (FIV)-positive and FIV-negative cats. Am. J. Vet. Res., 57: 1443-1446.
PubMed - Schibli, D.J., P.M. Hwang and H.J. Vogel, 1999. The structure of the antimicrobial active center of lactoferricin B bound to sodium dodecyl sulfate micelles. Fed. Eur. Biochem. Soc. Lett., 446: 213-217.
CrossRefDirect Link - Schryvers, A.B., R. Bonnah, R.H. Yu, H. Wong and M. Retzer, 1998. Bacterial lactoferrin receptors. Excerpta Med. Int. Congr. Ser., 1195: 55-65.
PubMed - Sekine, K., Y. Ushida, T. Kuhara, M. Iigo and H. Baba-Toriyama et al., 1997. Inhibition of initiation and early stage development of aberrant crypt foci and enhanced natural killer activity in male rats administered bovine lactoferrin concomitantly with azoxymethane. Cancer Lett., 121: 211-216.
CrossRefDirect Link - Sharma, A.K., S. Karthikeyan, S. Sharma, S. Yadav, A. Srinivasan and T.P. Singh, 1998. Structures of buffalo and mare lactoferrins. Similarities, differences and flexibility. Adv. Exp. Med. Biol., 443: 15-21.
PubMed - Shimazaki, K.I., 2000. Lactoferrin. A marvelous protein in milk. Anim. Sci. J., 71: 329-347.
Direct Link - Spik, G., B. Coddeville, J. Mazurier, Y. Boune, C. Cambillaut and J. Montreuil, 1994. Primary and three-dimensional structure of lactotransferrin (lactoferrin) glycans. Adv. Exp. Med. Biol., 357: 21-32.
PubMed - Steijns, J.M. and A.C. van Hooijdonk, 2000. Occurrence, structure, biochemical properties and technological characteristics of lactoferrin. Br. J. Nutr., 84: 11-17.
CrossRefPubMedDirect Link - Swart, P.J., M.E. Kuipers, C. Smit, R. Pauwels and M.P. deBethune et al., 1996. Antiviral effects of milk proteins: Acylation results in polyanionic compounds with potent activity against human immunodeficiency virus types 1 and 2 in vitro. Aids Res. Hum. Retroviruses, 12: 769-775.
PubMed - Swart, P.J., M.E. Kuipers, C. Smit, B.W. Van der Strate, M.C. Harmsen and D.K. Meijer, 1998. Lactoferrin. Antiviral activity of lactoferrin. Adv. Exp. Med. Biol., 443: 205-213.
PubMed - Tanaka, K., M. Ikeda, A. Nozaki, N. Kato, H. Tsuda, S. Saito and H. Sekihara, 2000. Lactoferrin inhibits hepatitis C virus viremia in patients with chronic hepatitis C. Proceedings of the 4th International Conference on Lactoferrin: Structure, Function and Applications, May 18-22, Sapporo, Japan, pp: 361-367.
- Teraguchi, S., K. Shin, T. Ogata, M. Kingaku and A. Kaino et al., 1995. Orally administered bovine lactoferrin inhibits bacterial translocation in mice fed bovine milk. Applied Envirn. Microbiol., 61: 4131-4134.
PubMedDirect Link - Tomita, M., H. Wakabayashi, K. Yamauchi, S. Teraguchi and H. Hayasawa, 2002. Bovine lactoferrin and lactoferricin derived from milk: Production and applications. Biochem. Cell Biol., 80: 109-112.
PubMed - Tsuda, H., K. Sekine, K. Fujita and M. Ligo, 2002. Cancer prevention by bovine lactoferrin and underlying mechanisms-A review of experimental and clinical studies. Biochem. Cell Biol., 80: 131-136.
PubMed - Uchida, Y., K. Sekine, T. Kuhara, N. Takasuka, M. Ligo, M. Maeda and H. Tsuda, 1999. Possible chemopreventive effects of bovine lactoferrin on esophagus and lung carcinogenesis in the rat. Jpn. J. Cancer Res., 90: 262-267.
CrossRefPubMedDirect Link - Valenti, P. and G. Antonini, 2005. Lactoferrin: An important host defence against microbial and viral attack. Cell. Mol. Life Sci., 62: 2576-2587.
CrossRef - Van Berkel, P.H., M.E. Geerts, H.A. van Veen, M. Mericskay, H.A. de Boer and J.H. Nuijens, 1997. N-terminal stretch Arg2, Arg3, Arg4 and Arg5 of human lactoferrin is essential for binding to heparin, bacterial lipopolysaccharide, human lysozyme and DNA. Biochem. J., 328: 145-151.
Direct Link - Van Veen, H.A., M.E.J. Geerts, P.H.C. van Berkel and J.H. Nuijens, 2004. The role of N-linked glycosylation in the protection of human and bovine lactoferrin against tryptic proteolysis. Eur. J. Biochem., 271: 678-684.
CrossRefPubMedDirect Link - Vorland, L.H., 1999. Lactoferrin: A multifunctional glycoprotein. Acta Pathol. Microbiol. Immunol. Scand., 107: 971-981.
PubMed - Wakabayashi, H., S. Abe, S. Teraguchi, H. Hayasawa and H. Yamaguchi, 1998. Inhibition of hyphal growth of azole-resistant strains of Candida albicans by triazole antifungal agents in the presence of lactoferrin-related compounds. Antimicrob. Agents Chemother., 42: 1587-1591.
PubMedDirect Link - Wakabayashi, H., W. Bellamy, M. Takase and M. Tomito, 1994. Inactivation of Listeria monocytogenes by Lactoferricin, a potent antimicrobial peptide derived from cow`s milk. J. Food Prot., 55: 238-240.
Direct Link - Wakabayashi, H., M. Takase and M. Tomita, 2003. Lactoferricin derived from milk protein lactoferrin. Curr. Pharm. Des., 9: 1277-1287.
PubMedDirect Link - Wakabayashi, H., K. Yamauchi and M. Takase, 2006. Lactoferrin research, technology and applications. Int. Dairy J., 16: 1241-1251.
CrossRef - Wang, D., K.M. Pabst, Y. Aida and M.J. Pabst, 1995. Lipopolysaccharide-inactivating activity of neutrophils is due to lactoferrin. J. Leuicoc Biol., 57: 865-874.
PubMed - Ward, P.P., E. Paz and O.M. Conneely, 2005. Multifunctional roles of lactoferrin: A critical overview. Cell. Mol. Life Sci., 62: 2540-2548.
PubMedDirect Link - Weinberg, E.D., 2007. Antibiotic properties and applications of lactoferrin. Curr. Pharma. Des., 13: 801-811.
PubMedDirect Link - Wong, H. and A.B. Schryvers, 2003. Bacterial lactoferrin-binding protein A binds to both domains of the human lactoferrin C-lobe. Microbiology, 149: 1729-1737.
CrossRef - Yamamoto, H., Y. Ura, M. Tanemura, A. Koyama and T.S. Sayaka et al., 2010. Inhibitory effect of bovine lactoferrin on human parainfluenza virus type 2 Infection. J. Health Sci., 56: 613-617.
Direct Link - Yamauchi, K., M. Tomita, T.J. Giehl, R.T. Ellison III, 1993. Antibacterial activity of lactoferrin and pepsin-derived lactoferrin peptide fragment. Infec. Immun., 61: 719-728.
Direct Link - Yekta, M.A., F. Verdonck, W. Van Den Broeck, B.M. Goddeeris, E. Cox and D. Vanrompay, 2010. Lactoferrin inhibits E. coli O157: H7 growth and attachment to intestinal epithelial cells. Vet. Med., 55: 359-368.
Direct Link - Yi, M., S. Kaneko, D.Y. Yu and S. Murakami, 1997. Hepatitis C virus envelope proteins bind lactoferrin. J. Virol., 71: 5997-6002.
PubMed - Zagulski, T., P. Lipinski, A. Zagulska, S. Broniek and Z. Jarzabek, 1989. Lactoferrin can protect mice against a lethal dose of Escherichia coli in experimental infection in vivo. Br. J. Exp. Pathol., 70: 697-704.
PubMed - Zhang, P., V. Sawicki, A. Lewis, L. Hanson, J.H. Nuijens and M.C. Neville, 2001. Human lactoferrin in the milk of transgenic mice increases intestinal growth in ten-day-old suckling neonates. Adv. Exp. Med. Biol., 501: 107-113.
PubMed