
Getachew Tafere Abrha
Department of Biotechnology, Mekelle University, Ethiopia
Firew Mekbib
School of Biotechnology, Haramaya University, Ethiopia
Belayneh Admassu
Holleta Agricultural Research Center, Central Ethiopia
Asian Journal of Plant Sciences
Year: 2013 | Volume: 12 | Issue: 6-8 | Page No.: 262-270







ABSTRACT
Brassica carinata A. Braun is an amphi-diploid species that originated from interspecific hybridization between B. nigra and B. oleracea in the highlands of Ethiopia. The crop has many desirable agronomic traits. Conventional propagation alone was not successful enough in Brassica due to high degree of segregation upon cross-pollination. Size and type of explants affect in vitro response and are limiting factors. Hence, the objective of this study was to establish an in vitro regeneration and propagation protocol. The explants were tested for callus induction, shoot and root regeneration in MS medium with 3% sucrose and 6% agar under different concentrations of PGRs including NAA, 2, 4-D, BAP. Cotyledon was the most responsive for callus induction at a higher rate in short period of time. The highest frequencies of callus growth were observed 80.7% from hypocotyl, 95% from cotyledon with 0.5 mg L-1 2, 4-D. Two types of calli were obtained: White and friable callus with large cells; green and compact callus with smaller cells. For shoot induction, successful shoot regeneration from white/friable callus was achieved at 2 mg L-1 of BAP. Maximum shoot induction was recorded in the hypocotyl (90%) with 2 mg L-1 of BAP. Highest percentage of shoot with root (98.7%) and highest mean number of roots per shoot (9 cm) occurred on medium with 0.3 mg IBA while the maximum root length (4.7 cm) was attained on the free hormone. Plantlets were successfully acclimatized in potting medium containing a mixture of 25% sand: 50% red soil: 25% compost on acclimatization pots (1:2:1).
PDF Abstract XML References Citation
Received: January 21, 2014;
Accepted: February 28, 2014;
Published: May 16, 2014
How to cite this article
Getachew Tafere Abrha, Firew Mekbib and Belayneh Admassu, 2013. In vitro Plant Regeneration from Callus of Hypocotyls and Cotyledonary
Explants in Ethiopian Mustard (Brassica carinata A. Braun), Yellow Dodolla
Cultivar. Asian Journal of Plant Sciences, 12: 262-270.
DOI: 10.3923/ajps.2013.262.270
URL: https://scialert.net/abstract/?doi=ajps.2013.262.270
DOI: 10.3923/ajps.2013.262.270
URL: https://scialert.net/abstract/?doi=ajps.2013.262.270
INTRODUCTION
In Ethiopia, among the oilseeds, Ethiopian mustard stands third next to niger seed and linseed in total production and area coverage (CSA, 2009). Its area and production are estimated at 34,580 ha and 47,721 t, respectively, at private peasants holdings level, with an average productivity of 0.68 t ha-1 (CSA, 2009). This low production is attributed to a number of production constraints such as: Lack of high yielding, early maturing varieties, high erucic acid (C22: 1) content in seed oil and high glucosinolate content in the meal (EARO, 2000).
Ethiopian mustard, B. carinata. A. Braun (BBCC, 2n = 4x = 34) is believed to have originated in the Ethiopia plateau through natural hybridization of B. nigra (BB, 2n = 2x = 16) with B. oleracea (CC, 2n = 2x = 18) and followed by the chromosome doubling of the hybrid plant) which has several agronomical important traits such as non dehiscent siliques and a much more developed and aggressive root system than B. napus. It is re resistant to a wide range of diseases and pests and is tolerant to many abiotic stresses (Katiyar et al., 1986) which makes it a suitable candidate as a food security crop in Ethiopia.
Plant transformation systems have been developed for many economically important species of the genus Brassica such as B. napus (Moloney et al., 1989), B. oleracea (De Block et al., 1989), B. juncea (Barfield and Pua, 1991), B. rapa (Radke et al., 1992), B. nigra (Gupta et al., 1993) and B. carinata (Babic et al., 1998) this technology enables us to obtain transgenic plants with modified agronomic traits. Many genetic improvements, such as herbicide tolerance, improved oil quality and production of pharmacological and industrial products, have been achieved by genetic transformation in the Brassica species. For example, B. napus seeds containing high levels of gamma-linoleic acid were obtained by the introduction of δ12-desaturase genes from the fungus Mortierella alpine (Liu et al., 2001). In addition B. carinata used to produce biodiesel and erucic acid derivatives can be used as chemical additives in plastic, tannery and cosmetic industries (Bozzini et al., 2007). It is also used as a luminant or lubricant in soap making (Greville, 2005).
Genetic improvement of Brassica spp. has been mainly achieved by conventional breeding methods. Recently, genetic engineering opened a new avenue for plant improvement (Hansen and Wright, 1999). Regeneration in Brassica is highly genotype and age dependent and has been reported in several species (Guo et al., 2005). In B. napus cultivar GSL-1 showed better regeneration efficiency than Westar (a standard cultivar for transformation) in one study (Phogat et al., 2000).
In vitro regeneration is a key factor in developing an efficient transformation method in plants. In vitro regeneration in Brassica ssp. is highly genotype-dependent as reported in previous studies for B. napus (Ono et al., 1994; Phogat et al., 2000), B. rapa (Zhang et al., 1998) and B. oleracea (Sparrow et al., 2006). In addition, Dieter et al. (1982) compared 6 species of the genus Brassica for callus growth and plant regeneration and reported a high influence of the genotype, with as much inter-cultivar as inter-species differences in the response to the in vitro culture. However, the available information for the genotype and explant variability for in vitro culture and shoot regeneration in B. carinata is limited to a small number of genotypes.
These genotype and explants type of the in vitro culture are a limiting factor for the application of genetic engineering to a wide number of genotypes. For that reason, it is important to identify highly regenerant genotypes that can be used in transformation via Agrobacterium tumefaciens.
In fact no study has been done on the protocols and the effects of various growth regulators on the callus formation from the hypocotyls and cotyledons explants of Yellow Dodola cultivar. But there are a number of studies have been carried out in the direct regeneration of Brassica spp. and remarkable progress has been achieved. Therefore, the objective of this study was to establish an in vitro regeneration and propagation protocol for the rapseed (B. carinata), Yellow Dodolla cultivar using hypocotyls and cotyledons explants, to identify promising explants with high regeneration potential.
MATERIALS AND METHODS
Plant material: The experiment was conducted in Holleta Agricultural Biotechnology Laboratory. The seed of Yellow Dodola was obtained from the Highland Oil Crops Research Department of Holleta Agricultural Research Center, were used for the experiments.
Surface sterilization and germination: Seeds of Brassica carinata genotype Yellow Dodola were first washed by submerging them in tap water for 5 min to remove the dust. The seeds were transferred to 70% ethanol for 1 min and then in 1% NaOCl bleach for 20 min. The surface sterilized seeds were transferred to autoclaved distilled water and rinsed 2-3 times. The pH of the culture medium was adjusted to 5.8 using 1.0 M HCl or 0.5 M NaOH before autoclaving. The prepared medium was poured into heat-sterilized glass containers; each contained about 40 mL solidified medium and autoclaved at 15 psi at 121°C for 15 min. The seeds were plated for germination on full strength MS free media in culture magentas under laminar flow hood on 6% agar (w/v). Later they were incubated at 25±2°C in a 16/8 h day/night photoperiodic regime under cool white fluorescent lights (2700-Lux) for 5-7 days.
Cotyledonary and hypocotyls leaves were excised in a laminar flow hood from 5-7 days old seedlings and cut into 0.5-1 cm pieces of explants excluding meristematic axillary bud. The 5 segment pieces of explants per magenta were placed horizontally in callus induction medium on MS medium supplemented with different concentrations of 2, 4-D, NAA and BAP.
Callus induction medium: Explants were cultured on the basal MS-medium they were supplemented with three hormones 2, 4-D, BAP and NAA to enhance callus initiation. At the concentration of: 0.1, 0.25, 0.5, 1.0 and 1.5 mg L-1, 0.1, 0.25, 0.5, 1 and 1.5 mg L-1 and 0.5, 1, 1.5 2.0, 2.5 mg L-1, respectively.
Each magenta contained 5 segments and all the treatments were performed with 3 replications and incubated in a complete dark. The magenta were sealed with parafilm and maintained in darkness for one month. The explants were sub-cultured 4-5 weeks on corresponding medium freshly prepared until shoot was formed. The number of explants that induced calli in (%), nature of Callus and weight (g) were recorded for each treatment.
Shoot induction medium: About 30-48 days after culture, calli were transferred aseptically on to a sterile petri dish and were cut into convenient size by a sterile scalpel or blade. The pieces of calli were plated on to freshly prepared media supplemented with BAP concentrations. Each magenta contained 5 pieces of calli and all the treatments were performed with 3 replications and sealed with parafilm in a growth room under 16 h light at 2700 lux light intensity and 8 h dark cycle at 25±2°C. The explants were Sub-cultured at 2-3 weeks intervals on corresponding medium freshly prepared to obtain shoot regeneration. The parameters recorded were the percentage of explant producing shoots (%), the mean number of shoots produced per explants and shoot height attained in cm.
Root induction medium: Eight weeks later regenerated shoots were carefully removed from the culture magentas and cut from the basal end of the shoots. Then each of the shoots was cultured in freshly prepared MS medium containing different hormonal supplements with IBA and NAA at 0.0, 0.1, 0.3 and 0.5 mg L-1 for root initiation. The media contained 2% sucrose and solidified with 0.6% agar, pH was adjusted at 5.8 and autoclaved at 15 psi at 121°C for 15 min. The data on rooting were collected after four weeks of culture. Parameters recorded were the percentage of explants producing root (%), the mean number of roots produced per explants, root length attained (cm) and the fresh and dry weight of shoots attained (cm). Plantlets with approximate height of 4-7.7 cm, with well developed roots were transferred to small acclimatization pot, containing sterilized soil mix with a proportion of 1:2:1 sand, red soil and compost, respectively.
Experimental design and statistical analysis: The experiments were arranged in a Complete Randomized Design (RCD), with three replications and each replication per treatment contained 5 explants. Data were analyzed using the Analysis of Variance (ANOVA) using Statistical Analysis System (SAS versions) software and the least significance differences among mean value were compared using DMRT at (p≤0.05) for comparison between treatment means.
RESULTS AND DISCUSSION
Callus initiation from hypocotyls and cotyledon explants: It was observed that the explants exhibited an initial swelling followed by callus formation within 1-2 weeks of incubation, except the control treatment. Callus proliferation started from cut ends of hypocotyls and cotyledon on MS medium supplemented with different concentrations of 2, 4-D, BAP and NAA and eventually extended all over the explants. Callus initiation began from cut edges of each explant after 6 days in cotyledon explants and after 12-14 days in hypocotyls explants. Callus produced from cotyledon explants grew faster during the first 7-20 days of culture. Approximately, 3 weeks after culturing, the explants were almost completely converted into callus. Similar results have been described in other reports. Ullah et al. (2004) reported that on B. napus cv. Rainbow explants, callus proliferation started from the cut ends of the hypocotyls and cotyledon. Al-Naggar et al. (2010) reported different results in B. napus: 2, 4-D induced callus production over the entire surface of the cotyledon and hypocotyls.
The type and quantity of callus and callogenesis efficiency depended on the duration of exposure to hormone and on the type of explant. In this experiment, the highest quantity of callus was formed on cotyledons after short induction on MS in 2, 4-D. BAP and NAA require long time to produce callus and caused poor formation of compact calli, with many necrotic sites. Abdellatef and Khalafallah (2008) and Dieter et al. (1982), obtained similar results in cotton and B. napus cv. Evitain, respectively: On MS media supplemented with 2, 4-D and BAP. Highly significant difference in percentage of callus formation was observed on the explants, PGR and genotype (Table 1). These results are in agreement with those reported by Sayed et al. (2010) and Khan et al. (2010).
Effects of 2, 4-dichlorophenoxy acetic acid: The 2, 4-D is among the most widely used auxin used for in vitro callus induction in a wide range of plant species (Al-Naggar et al., 2010; Khan et al., 2002). The effects of 2, 4-D concentrations on callus initiation revealed that Yellow Dodola, cotyledon produced the highest mean percentage of callus (95%) on MS medium supplemented with 0.5 mg L-1 2, 4-D (Table 1). In line with the present results Osman et al. (2010) reported that the most efficient hormone concentration depends on the genotype used and the type of explants. Comparison of callus initiation from hypocotyls and cotyledon explants (Table 1) showed that in 2, 4-D concentrations cotyledon explants gave higher callus initiation percentage than hypocotyls explants. The optimum 2, 4-D concentration for callus initiation from cotyledon explants was 0.5 mg L-1. It had the highest Callus Initiation Frequency (CIF) and produced calli with more mean weight than the other concentration levels. These results are in harmony with those reported by Ali et al. (2007) in B. napus.
Effects of benzyl adenine purine: As to the effect of different concentrations of BAP on callus initiation from hypocotyl and cotyledone, 0.5 mg L-1 BAP was effective in inducing callus from cotyledon explants than the hypocotyls. Higher concentration of BAP (1.5 mg L-1) inhibited callus proliferation. BAP at low concentrations were more effective for induction of callus. The present study showed that Callus Initiation Frequency (CIF) was 80 and 50% for Yellow Dodola cotyledon and hypocotyls explants, respectively. This study is consistent with the result of various research reports in B. napus (Datta Gupta and Conger, 1999; Yu et al., 2005; Khan et al., 2010; Pathirana and Eason, 2006; Rao et al., 2006).
| Table 1: | Effect of different concentrations of 2, 4 dichlorophenoxy acetic acid (2, 4-D), Naphthalene Acetic Acid (NAA) and Benzyl Adenine Purine (BAP) on callus induction physical characteristics |
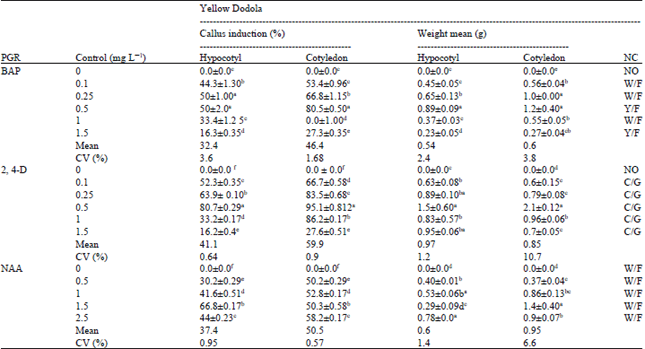 | |
| PGR: Plant growth regulators, NC: Nature of callus, W: White, F: Friable, C: Compact, G: Green. Mean within a column followed by the same superscript letters are not significantly different at (p≤0.05) | |
 | |
| Fig. 1(a-b): | Proliferation of callus from hypocotyl and cotyledon explants of Brassica carinata cultivars after 5 weeks of culture on full MS medium supplemented with 0.5 mg L-1 2, 4-D, (a) Compact/green callus formation from cotyledon of Yellow Dodola and (b) White/friable callus formation from the hypocotyl of Yellow Dodola |
Effects of naphthalene acetic acid: The highest callus induction (73%) was obtained on cotyledon explants cultured on MS medium supplemented with NAA at 1.5 mg L-1 followed by 66% obtained from the same explants by using 1.5 mg L-1 NAA using variety Holleta-1. In this respect, it was found that the best result of callus initiation from cotyledon was obtained in media with 1.5 mg L-1 of NAA. These results are in agreement with those reported by Osman et al. (2010) and Chamandosti et al. (2006).
Two morphological types of callus were noted after 30 days of culture: Calluses derived from hypocotyls segments became soft or friable, sticky, yellow-white to cream colored and nodular type and mostly consisted of large and translucent cells (Fig. 1b). While, calli from cotyledon explants contained small and green cells with nodular, green, organogenetic and compact callus and non friable often developed on media containing 2, 4-D and formed round masses (Fig. 1a).
 | |
| Fig. 2(a-f): | (a-b) New shoot initiation from cotyledon and hypocotyl explant of Yellow Dodola, respectively after 8 week of culture, (c) Shoot regeneration of hypocotyl derived calli on MS medium after 10 weeks of culture, (d) Root induction in MS medium supplemented with IBA, (e) Acclimatization of the in vitro plantlets of Yellow Dodola in the greenhouse after 2 weeks of removing the plastic bag and (f) Well developed plantlets 1 month after acclimatization |
This result is in consistence with the result obtained for sugar beet by Kamal et al. (2007) and Chamandosti et al. (2006) reported similar findings in B. napus, three morphologically distinct types of calli were induced by using different concentrations of 2,4-D and BAP.
Shoot initiation and regeneration via indirect organogenesis
Effect of different concentration of BAP on shoot initiation from the callus: Shoot proliferation occurred through a callus phase. The development of shoots (Fig. 2) from white friable and compact callus (regardless of the explants type) was observed after 3-4 weeks of culture. Shoot development was achieved at all concentration except the control treatment in hypocotyls and cotyledon explants (Table 2). The percentage of explants forming shoots varied greatly between the nature of callus, as well as between different explants (hypocotyl and cotyledon) on MS media supplemented with different concentrations of the BAP. These findings exhibited a varied response (22-90%) to shoot regeneration from the two explants of Yellow Dodolla. Similar results were obtained by Moghaieb et al. (2006) and Kamal et al. (2007) in B. napus.
The explants developed tiny patches of pale green callus bearing several hump-like structures within a period of 20-30 days. Subsequently, these differentiated in to shoot buds. Best shoots elongation was achieved when regenerating segment of the callus was cut in to smaller pieces containing three or five shoots and sub cultured in the basal medium supplemented with 2 mg L-1 BAP. These results are in harmony with those reported by Moghaieb et al. (2006), Zeynali et al. (2010) and Kamal et al. (2007) in B. napus. Results of the effect of BAP on shoot initiation on the two explants of B. carinata (Yellow Dodolla) are presented in Table 2 and Fig. 2. The result revealed that all varieties respond to all treatments. Morphologically best shoots were induced on the medium that contained 2 mg L-1 BAP (Fig. 2). Highest percentage of survival rate of cultured explants was obtained at 2 mg L-1 of BAP.
| Table 2: | Shoot regeneration frequencies of the hypocotyls and cotyledon of Yellow Dodola cultivars obtained from callus in MS medium with different concentration of BAP after 8 weeks of culture |
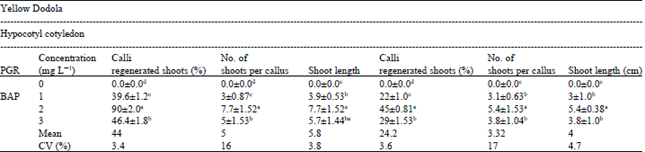 | |
| Mean within a column followed by the same superscript letters are not significantly different at (p≤0.05) | |
This holds true for the two explants in which, Shoot Regeneration Frequency (SRF) is 90% for hypocotyl explants and 45% for cotyledonary explants of Yellow Dodola as evident from (Table 2) at 2 mg L-1 of BAP at 2 mg L-1 of BAP. Based on this result, 2 mg L-1 BAP concentration was used as an optimum PGRs for shoot initiation. Similar result has been reported by Ravanfar et al. (2009) and Chamandosti et al. (2006) in B. oleracea and B. napus.
The rate of shoot regeneration was better in hypocotyl explants than cotyledons. The same results have been reported by Khan et al. (2010) one of the important factors for shoot regeneration were type and and size. It is also in agreement with Moghaieb et al. (2006), Yang et al. (1991) and Yu et al. (2005). Thus it is clearly shown that the regeneration depend on type and size of explant.
Effect of different concentrations of BAP on number and height of shoots: Among the four BAP concentrations 2 mg L-1 of BAP produced significantly maximum mean number of shoots for the two explants (Table 2). hypocotyl explants gave maximum mean Number of shoots (90%), the maximum shoot height (7.7 cm) was attained at 2 mg L-1 of BAP, respectively from the hypocotyls explants (Table 2), the results agreed with Munshi et al. (2007) who found that the highest shoot induction frequency was obtained on the medium containing 2 mg L-1 BAP. Medium supplemented with 2 mg L-1 of BAP was the best in many of the parameters recorded. There was also a significant difference between cultivars in number of shoots developed and shoots length. Hypocotyl was the best in both traits (Table 2).
The responses of explants for shoots regeneration were different. Various authors Munshi et al. (2007) and Gubi et al. (2004) reported that hypocotyl explants were superior to cotyledon in shoot regeneration efficiency and shoot length. MS medium supplemented with 2 mg L-1 BAP was the most effective in adventitious shoot formation and shoot length in both explants. This result agreed with George et al. (2008) who showed that one of the B. napus cultivars named RK-7 had low shoot regeneration (18%) from cotyledonary explants while the same cultivar had a higher (27%) shoot regeneration from the hypocotyl explants.
Root organogenesis
Effect of NAA and IBA on root induction: The rooting result revealed that plantlets cultured on rooting media induced roots in all media supplemented with NAA and IBA including hormone free media. The primary roots were observed after 2 weeks of culturing on MS medium supplemented by different concentration of IBA and NAA. However, well developed and mature roots were obtained after a month. Rooting of plantlets was usually attained at half MS medium and low concentration of auxins and sucrose. Similar result was reported by Khan et al. (2002).
The treatment containing 0.3 mg L-1 IBA and the control (MSO) gave the highest percentage of plantlets producing roots and mean number of roots per plantlet in both genotypes (Table 3). Among the different auxins, IBA was the best for root induction. The maximum number of plantlets producing roots and the highest mean number of roots per shoot was achieved at a concentration of 0.3 mg L-1 (Table 3). The highest percentage of explant producing roots (98%) and mean number of roots produced per explants (9) was observed on the cultivar supplemented with IBA (0.3. mg L-1) and the lowest frequency of response (41%) on the cultivar fortified with NAA (0.5 mg L-1). NAA supplemented medium resulted in low root induction and shoots in the medium had a tendency to call us at the base of shoots and produced stumpy and thick roots. These results are in agreement with the findings of Ravanfar et al. (2009) and Caboni and Tonelli (1999) who reported that IBA is the most effective auxin for root induction in a wide range of plant species. IBA is superior is stability to NAA. The present study also proved that IBA was better than NAA for root induction. Similarly Ali et al. (2007) reported a similar result by using 0.3 mg L-1 of IBA in B. napus (Westar) 90% root formation was recorded and Salman (2002) also reported similar results in B. oleracea.
| Table 3: | Effect of auxin types on percentage of shoots with root (%) mean number of roots produced per shoot, root length (cm) and fresh and dry weight (g) of Yellow Dodola after 4 weeks of culture |
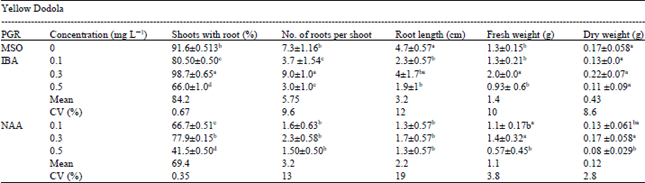 | |
| Means within the same superscript letter were not significantly different at p≤0.05 probability level according to DMRT | |
The second maximum frequency of root formation (91%) of Yellow Dodola was achieved on half strength MS medium without growth regulators in both genotypes and maximum root length (4.7 cm) was attained on MSO medium. It is assumed that root formation on auxin free medium may be due to the availability of higher-quantity of endogenous auxin in shootlets raised in vitro. Similar results were reported by Ali et al. (2007) in B. napus. Both treatments showed significant difference with respect to the percentage of shoots forming roots, mean number of roots produced per shoot, root length, fresh and dry weight. In this study the highest increase in fresh and dry weight was observed in medium supplemented with 0.3 mg L-1 IBA. There was a significant difference between treatment IBA and NAA on root length attained. NAA produced fewer roots than that of IBA. Similar results were obtained by various authors (Moghaieb et al., 2006; Chamandosti et al., 2006; Kamal et al., 2007).
CONCLUSION
Callus proliferation started from cut end edges and wound part of the hypocotyl and cotyledon explants gave higher callus initiation percentage than hypocotyl explants. The BAP and NAA took longer time to produce callus and caused the formation of poor and compact calli with many necrotic sites. Successful shoot regeneration was achieved from white compact callus. The 2 mg L-1 BAP is better for shoots induction in B. carinata cultivars and B. carinata cultivars is response to shoots formation and shoots length. The highest percentage of shoots producing roots and mean number of roots per explants was achieved on hormone free. The survival rate of regenerated plants was 100%.
ACKNOWLEDGMENTS
Special thanks to the Plant Biotechnology Laboratory of Holleta Agricultural Research center for their generous help and we wish to thank Dr. Adefris Teklewold from the Ethiopian Institute of Agricultural Research.
REFERENCES
- Abdellatef, E. and M.M. Khalafallah, 2008. Influence of growth regulators on callus induction from hypocotyls of medium staple cotton (Gossypium hirsutum L.). cultivar barac B-67. J. Soil Nat., 2: 17-22.
Direct Link - Ali, H., Z. Ali, H. Ali, S. Mehmood and W. Ali, 2007. In vitro regeneration of Brassica napus L., cultivars (Star, Cyclone and Westar) from hypocotyls and cotyledonary leaves. Pak. J. Bot., 39: 1251-1256.
Direct Link - Babic, V., R.S. Datla, G.J. Scoles and W.A. Keller, 1998. Development of an efficient Agrobacterium-mediated transformation system for Brassica carinata. Plant Cell Rep., 17: 183-188.
CrossRefDirect Link - Barfield, D.G. and E.C. Pua, 1991. Gene transfer in plants of Brassica juncea using Agrobacterium tumefaciens-mediated transformation. Plant Cell Rep., 10: 308-314.
CrossRef - Bozzini, A., F. Calcagno and T. Soare, 2007. [Sincron, a new Brassica carinata cultivar for biodiesel production]. HELIA, 30: 207-214 (In Spanish).
Direct Link - Caboni, E. and M.G. Tonelli, 1999. Effect of 1, 2-benzisoxazole-3-acetic acid on adventitious shoot regeneration and in vitro rooting in apple. Plant Cell Rep., 18: 985-988.
CrossRef - Chamandosti, F., A. Majd and S. Mehrabian, 2006. In vitro plant regeneration from callus of cotyledons in canola (Brassica napus L.). Pak. J. Biol. Sci., 9: 302-306.
CrossRefDirect Link - Gupta, S.D. and B.V. Conger, 1999. Somatic embryogenesis and plant regeneration from suspension cultures of switch grass. Crop Sci., 39: 243-247.
Direct Link - Dietert, M.F., S.A. Barron and O.C. Yoder, 1982. Effects of genotype on in vitro culture in the genus Brassica. Plant Sci. Lett., 26: 233-240.
CrossRefDirect Link - Yu, F., D. Zhang, F. Bai and L. An, 2005. The accumulation of isocamptothecin A and B in suspension cell culture of Camptotheca acuminata. Plant Cell Tissue Organ Cult., 81: 159-163.
CrossRefDirect Link - Gubi, J., Z. Lajchova, J. Farag and Z. Jurekova, 2004. Effect of growth regulators on shoot induction and plant regeneration in tomato (Lycopersicon esculentum Mill.). Biol. Bratislava, 59: 405-408.
Direct Link - Guo, D.P., Z.J. Zhu, X.X. Hu and S.J. Zheng, 2005. Effect of cytokinins on shoot regeneration from cotyledon and leaf segment of stem mustard (Brassica juncea var. tsatsai). Plant Cell Tissue Org. Cult., 83: 123-127.
CrossRefDirect Link - Gupta, V., G.L. Sita, M.S. Shaila and V. Jagannathan, 1993. Genetic transformation of Brassica nigra by Agrobacterium based vector and direct plasmid uptake. Plant Cell Rep., 12: 418-421.
CrossRef - Kamal, G.B., K.G. Illich and A. Asadollah, 2007. Effects of genotype, explant type and nutrient medium components on canola (Brassica napus L.) shoot in vitro organogenesis. Afr. J. Biotechnol., 6: 861-876.
Direct Link - Katiyar, R.K., G. Saran and G. Giri, 1986. Evaluation of Brassica carinata as a new oilseed crop in India. Exp. Agric., 22: 67-70.
CrossRef - Khan, M.R., H. Rashid and A. Quraishi, 2002. Effects of various growth regulators on callus formation and regeneration in Brassica napus Cv. Oscar. Pak. J. Biol. Sci., 5: 693-695.
CrossRefDirect Link - Khan, M.R., H. Rashid and A. Quraishi, 2010. Increased regeneration efficiency of Brassica napus L. cultivars Star, Westar and Cyclone from hypocotyle and cotyledonary explants. Plant Tissue Cult., 12: 131-138.
Direct Link - Liu, J. W., S. DeMichele, M. Bergana, E. Bobik Jr. and C. Hastilow et al., 2001. Characterization of oil exhibiting high γ-linolenic acid from a genetically transformed canola strain. J. Am. Oil Chem. Soc., 78: 489-493.
CrossRef - Osman, M.G., E.A. Elhadi and M.M. Khalafalla, 2010. Callus formation and organogenesis of tomato (Lycopersicon esculentum Mill, CV Omdurman) induced by thidiazuron. Afr. J. Biotechnol., 9: 4407-4413.
Direct Link - Moghaieb, R.A.A., M.A. El-Awady, R.G. El-Mergawy, S.S. Youssef and A.M. El Sharkawy, 2006. A reproducible protocol for regeneration and transformation in canola (Brassica napus L.). Afr. J. Biotechnol., 5: 143-148.
CrossRefDirect Link - Moloney, M.M., J.M. Walker and K.K. Sharma, 1989. High efficiency transformation of Brassica napus using Agrobacterium vectors. Plant Cell Rep., 8: 238-242.
CrossRef - Ono, Y., Y. Takahata and N. Kaizuma, 1994. Effect of genotype on shoot regeneration from cotyledonary explants of rapeseed (Brassica napus L.). Plant Cell Rep., 14: 13-17.
CrossRef - Pathirana, R. and J.R. Eason, 2006. Establishment and characterisation of a rapidly dividing diploid cell suspension culture of Arabidopsis thaliana suitable for cell cycle synchronisation. Plant Cell Tissue Org. Cult., 85: 125-136.
CrossRefDirect Link - Phogat, S.K., P.K. Burma and D. Pental, 2000. High frequency regeneration of Brassica napus varieties and genetic transformation of stocks containing fertility restorer genes for two cytoplasmic male sterility systems. J. Plant Biochem. Biotechol., 9: 73-79.
CrossRef - Radke, S.E., J.C. Turner and D. Facciotti, 1992. Transformation and regeneration of Brassica rapa using Agrobacterium tumefaciens. Plant Cell Rep., 11: 499-505.
CrossRef - Rao, A.Q., S.S. Hussain, M.S. Shahzad, S.Y.A. Bokhari and M.H. Raza et al., 2006. Somatic embryogenesis in wild relatives of cotton (Gossypium Spp.). J. Zhejiang Univ. Sci. B, 7: 291-298.
CrossRefDirect Link - Salman, M.N., 2002. Establishment of callus and cell suspension cultures from Gypsophila paniculata leaf segments and study of the attachment of host cells by Erwinia herbicola pv. gypsophilae. Plant Cell Tissue Organ Cult., 69: 189-196.
CrossRef - Ullah, I., R. Hamid and M.R. Khan, 2004. Establishment of tissue culture protocol in brassica (B. napus L.). Pak. J. Biol. Sci., 7: 277-278.
CrossRefDirect Link - Yang, M.Z., S.R. Jia and E.C. Pua, 1991. High frequency of plant regeneration from hypocotyl explants of Brassica carinata A. Br. Plant Cell Tissue Org. Cult., 24: 79-82.
CrossRefDirect Link - Zeynali, M., M.B.M. Zanjani, M.E. Amiri, M. Noruzian and S.M. Aghajari, 2010. Influence of genotype and plant growth regulator on somatic embryogenesis in rapeseed (Brassica napus L.). Afr. J. Biotechnol., 9: 4050-4055.
Direct Link - Zhang, F.L., Y. Takahata and J.B. Xu, 1998. Medium and genotype factors influencing shoot regeneration from cotyledonary explants of Chinese cabbage (Brassica campestris L. ssp. pekinensis). Plant Cell Rep., 17: 780-786.
CrossRef - Ravanfar, S.A., M.A. Aziz, M.A. Kadir, A.A. Rashid and M.H.T. Sirchi, 2009. Plant regeneration of Brassica oleracea subsp. Italica (Broccoli) CV Green Marvel as affected by plant growth regulators. Afr. J. Biotechnol., 8: 2523-2528.
Direct Link - Munshi, S.K., S. Sandhu and S. Sharma, 2007. Lipid composition in fast and slow germinating sunflower (Helianthus annuus L.) seeds. Gen. Appl. Plant Physiol., 33: 235-246.
Direct Link