
Athidtaya Kumvinit
Division of Plant Pathology, Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, Chiang Mai 50200, Thailand
LiveDNA: 66.22018
Angsana Akarapisan
Division of Plant Pathology, Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, Chiang Mai 50200, Thailand
LiveDNA: 66.16079
Asian Journal of Plant Pathology
Year: 2020 | Volume: 14 | Issue: 1 | Page No.: 27-35







ABSTRACT
Background and Objective: Bacillus spp. have a wide range of activities in the biocontrol potential against various phytopathogens. This study was focused to identify the antagonistic bacteria and to evaluate for their efficacy against the bacterial pathogens of potato. Materials and Methods: Bacillus sp. strains BB06, BB10, CF58 and CR74 were previously isolated from the rhizosphere. They were identified by 16S rRNA gene and gyrA gene. The strains were tested for hydrolytic enzyme and plant growth-promoting activities. Also, the strains were analyzed the surfactin antibiotic by LCMS-QTOP. Results: Based on the 16S rRNA gene and gyrA gene sequence analyses, the strains were identified as Bacillus velezensis. All strains showed a broad inhibitory spectrum and produced zones of inhibition against bacterial pathogens of potato, included Pectobacterium carotovorum subsp. brasiliense, Dickeya dadantii, Bacillus altitudinis and Ralstonia solanacearum. The results of in vitro assays showed that all isolates can produce protease, amylase, cellulase, lipase, IAA (auxin), siderophores and phosphate solubilization. Further analysis of the surfactin antibiotic by LCMS-QTOP, revealed the characteristic molecular ions mode [M-H-] with an m/z around 1034. The strains showed analogous biofilm formation to support the biocontrol properties. Conclusion: B. velezensis strain BB06, BB10, CF58 and CR74 can be recommended as potential biocontrols.
PDF Abstract XML References Citation
How to cite this article
Athidtaya Kumvinit and Angsana Akarapisan, 2020. Surfactin Production, Biofilm Formation and Antagonistic Activity Against Phytopathogens by Bacillus velezensis. Asian Journal of Plant Pathology, 14: 27-35.
URL: https://scialert.net/abstract/?doi=ajppaj.2020.27.35
URL: https://scialert.net/abstract/?doi=ajppaj.2020.27.35
INTRODUCTION
Bacillus species are gram-positive, aerobic and endospore-forming bacteria that promote plant growth. Bacillus species are benefits as opposed to other organisms due to their ability to form endospores and can tolerate high temperatures and pH conditions. The Bacillus subtilis species complex is a nonpathogenic bacterial group that associates with plant roots and live in the soil1. The B. subtilis species complex is considered to be important PGPR (Plant growth-promoting rhizobacteria). The PGPR can be categorized into three general forms: biopesticides, phytostimulators and bio-fertilizers. Furthermore, B. subtilis species complex are able to augment plant growth by a wide range of means through siderophores production, phosphate solubilization, biological nitrogen fixation, production of phytohormones [Indole Acetic Acid (IAA), cytokinins, ethylene and gibberellin], production of hydrolysis enzymes (protease, amylase, cellulase and lipase), induction of systemic resistance and inhibiting plant pathogens by antibiosis2-4.
Many strains of Bacillus species have been reported to possess the ability to subdue the growth of fungal and bacterial pathogens on plants. For example, B. subtilis was found to decrease disease severity and in particular suppressed bacterial wilt disease prevalence in potato cultivars. Diamante and Spounta5 B. amyloliquefaciens are endowed with the potential in hindering the development of bacterial soft rot on Chinese cabbage by showing a clear zone of inhibition in vitro6. In many cases, Bacillus spp. showed high antagonistic activity against pathogens of plants, triggered biofilm formation and produced lipopeptides7.
The lipopeptides produced by Bacillus spp. are highly surface-active molecules that belong to the class of surfactin, iturin and fengycin8. Apparently, the surfactants of the lipopeptide group are the most researched in the Bacillus groups. Bacillus species are primary determinants of biocontrol and aggregation in biofilms, especially the surfactin family which is involved in the formation of biofilms. Surfactins play an important role in biofilm formation employing the swarming motility of the cells9. Also, surfactins fashioned in a manner that is specific to a particular species act naturally as a signal for biofilm formation and root colonization7. The B. subtilis strain 168 has srfAA genes that jeopardize surfactin production10.
However, the identification of useful bacterial antagonists is problematic because in closely related species there is a high percentage of sequence similarity. The use of protein-encoding genes is a common analytical approach and often uses the sequence of the 16S rRNA gene for identifying bacteria and constructing phylogenetic trees11-13. Phylogenetic scrutiny executed with partial 16S rRNA and gyrA gene structures exposed the strains of the genus Bacillus that are linked to plants. The gyrA genetic factor that encodes the subunit A protein of DNA gyrase was used in taxonomic recognition of closely linked bacterial species of the B. subtilis species complex, for example, B. subtilis, B. amyloliquefaciens, B. velezensis, B. licheniformis, B. vallismortis, B. atrophaeus, B. sonorensis and B. mojavensis2. Prior to this study, the genetic characterization of 16S rRNA and gyrA gene sequences of Bacillus spp. was observed. The potential of the Bacillus spp. to control bacterial pathogens of potato was assessed and the production characteristic of lipopeptide antibiotic surfactin in biofilm formation was evaluated.
MATERIALS AND METHODS
Study area: This research worked in Bacterial Plant Pathology Laboratory, Faculty of Agriculture, Chiang Mai University, Thailand during August, 2017 to June, 2020.
Bacterial strain and phenotypic characterization: The four best antagonistic isolates of Bacillus sp. that were used in this study were provided by the Bacterial Plant Pathology Laboratory, Faculty of Agriculture, Chiang Mai University, including BB06 (from the rhizosphere of chili pepper), BB10 (from the rhizosphere of tomato), CF58 and CR74 (from the rhizosphere of potato). The media used were Nutrient Agar (NA) medium (3 g beef extract, 5 g peptone and 15 g agar per liter, pH = 7.0) and Luria-Bertani (LB) medium (10 g tryptone, 5 g yeast extract and 10 g sodium chloride per liter, pH 6.5).
The Bacillus sp. strains were characterized morphologically and biochemically by following Bergey’s Manual of Systematic Bacteriology14. Extraction of the DNA from antagonistic bacteria used a modification of the protocol15,16. Polymerase chain reaction (PCR) was performed to amplify a partial 16S rRNA gene and gyrA gene (subunit A of DNA gyrase). The 16S rRNA set included forward primer 16SF (5’ AGA GTT TGA TCC TGG CTC AG 3’) and reverse primer 16SR (5’AAG GAG GTG ATC CAG CCG CA 3’) for amplification of a 1500 bp region17. PCR reactions were performed as follows: initial denaturation at 94°C for 5 min, followed by 35 cycles consisting of 30 sec at 94°C, 30 sec at 64-64°C and 2 min at 72°C followed by a final extension at 72°C for 10 min. While, the primer set gyrA included forward primer gyrAF (5’ CAG TCA GGA AAT GCG TAC GTC CTT 3’) and reverse primer gyrAR (CAA GGT AAT GCT CCA GGC ATT GCT 3’) for amplification of a 970 bp region12. The conditions for amplification were as follows: initial denaturation at 94°C for 5 min, followed by 35 cycles consisting of 30 sec at 94°C, 30 sec at 60-64°C and 1 min at 72°C followed by a final extension at 72°C for 10 min. The sequences were deposited in the GenBank database and analyzed using a BLASTn algorithm-based program to identify the bacterial strains. Phylogenetic analysis was performed using the neighbour-joining method and the MAGA 10 software18.
Assay of antagonistic activity against bacterial pathogens of potato: In a paper disc method, each bacterial pathogen (1 mL of 108 cfu mL–1) was transferred to 99 mL of Nutrient Agar (NA) and poured into petri-dishes. Almost 20 μL of each Bacillus spp. suspension (108 cfu mL–1) were transferred using a pipette onto sterile filter paper (6 mm diameter) on NA plates containing the target bacterial pathogens of potato. The plates were incubated at 28°C for 3 days. Then the diameter of the clear halo surrounding the filter was measured. The experiment was performed with three replications.
Bacterial pathogens of potato were provided by the Bacterial Plant Pathology Laboratory, Faculty of Agriculture, Chiang Mai University and included Pectobacterium carotovorum subsp. brasiliense strain PY1, Dickeya dadantii strain CK211, Bacillus altitudinis strain CK50718 and Ralstonia solanacearum strain CD1.
Characterization of bacterial isolates for hydrolytic enzyme and plant growth-promoting activities: The antagonistic bacteria were evaluated for hydrolytic enzyme activity including protease, amylase, cellulase and lipase. A 20 μL drop of antagonistic bacterial suspension was dispensed onto a sterile filter paper disc (6 mm diameter) that was placed on the medium. The experiment was performed with four replications19-21.
Protease activity was tested on skim milk agar (28 g skim milk powder, 5 g casein hydrolysate, 25 g yeast extract, 1 g dextrose and 15 g agar per liter). The plates were incubated at 28°C for 48 hrs. A positive result showed a clear zone of inhibition around the disc.
Amylase activity was tested on starch hydrolysis medium (20 g soluble starch, 3 g beef extract, 5 g peptone and 15 g agar per liter). The plates were incubated at 37°C for 24 hrs. Following incubation, the surface of the plates was flooded with iodine solution and then the excess was poured off. A positive result showed a clear zone of inhibition around the disc.
Cellulase activity was tested on NA with 1% carboxy methyl cellulose. The plates were incubated at 37°C for 48 hrs.
After that, plates were flooded with 0.1% congo red reagent and left for 20 min and then were washed with 1 mol L–1 of NaCl. A clear zone against congo red was a positive result.
Lipase activity was tested on lipase enzyme medium which includes 10 g peptone, 5 g NaCl, 0.1 g CaCl2 and 15 g agar per liter. After the medium was autoclaved, it was cooled to about 50°C and 10 mL of autoclaved Tween 20 was added. The plates were incubated at 28°C for 48 hrs. A positive result was the production of a crystallized zone around bacterial colonies.
The antagonistic bacteria were further evaluated for their plant growth-promoting activity such as IAA production, siderophores production and phosphate solubilizing. A 20 μL drop of antagonistic bacterial suspension was dispensed onto a sterile filter paper disc (6 mm diameter) that was placed on the medium. The experiment was performed with four replications.
IAA production The bacteria were cultured in 10 mL of LB broth containing 0.1 g L–1 of L-tryptophan and incubated in dark on a shaker at 150 rpm at 30°C for 7 days. The broth was centrifuged at 1000×g for 30 min. The 1 mL of supernatant was added to 40 μL of orthophosphoric acid and 2 mL of Salkowski’s reagent (100 mL, 35% perchloric acid and 2 mL 0.5 mol L–1 FeCl3). After that, the supernatant was incubated in the dark at room temperature for 15 min. The red color development, which indicated IAA production was measured using a spectrophotometer at 530 nm absorbance. A standard curve was prepared using pure indole-3-acetic acid22.
Siderophore production was determined by a modified method on King’s B agar medium (90 mL, 1.8 g peptone, 0.135 g K2HPO4, 0.135 g MgSO4·7H2O and 1.8 g agar) with Chrome Azurol S (CAS) (10 mL, 60.5 mg CAS, 1 m mol L–1 FeCl3·6H2O in 10 m mol L–1 HCl and 72.9 mg HDTMA) and inoculated at 28°C for 72 hrs. The blue color the medium presented was changed to a light yellow or light orange halo surrounding the colony indicating the production of siderophores23.
Phosphate solubilization was tested on Pikovskaya’s agar (10 g glucose, 5 g tri-calcium phosphate, 0.5 g yeast extract, 0.2 g KCl, 0.1g MgSO4, 0.2 g NaCl2, 0.002 g FeSO4.7H2O, 0.002 g MnSO4 and 20 g agar containing 0.25% congo red) incubated at 28°C for 7 days. Phosphate solubilization was indicated by a clear halo24.
Analysis of surfactin: The surfactin antibiotic gene (srfAA gene) was amplified by PCR using the primers SRFA-F (5’ TCG GGA CAG GAA GAC ATC AT 3’) and SRFA-R (5’ CCA CTC AAA CGG ATA ATC CTG A 3’) for amplification of a 201 bp. The PCR amplification consisted of initial denaturation at 95°C for 5 min, followed by 35 cycles of denaturation at 95°C for 45 sec, annealing at 52°C for 45 sec, extension 72°C for 1 min and final extension 72°C for 10 min25. The sequences were deposited in the GenBank database and analyzed using a BLASTx algorithm-based program. Phylogenetic analysis was performed using the neighbour-joining method.
For the LCMS-QTOF analysis, the bacteria were cultivated in 20 mL of LB medium at room temperature for 4 days. (32±2°C). The cultures were incubated under gyratory agitation (200 rpm) in 100 mL Erlenmeyer flasks. After incubation, the bacterial suspension was removed by centrifugation (4000×g for 5 min at 4°C). After that, 15 g of the cell-free supernatant was mixed with an equal volume of acetonitrile (HPLC-grade) for 1 min and then the bacterial suspension was removed by centrifugation (4000×g for 5 min at 4°C). The supernatant was added to 7.5 g of QuEchERS Extract Pouch, AQAC Method (Agilent Technology) and centrifuged at 4000×g for 5 min at 4°C. Three mL of supernatant were loaded on the Agilent Captiva EMR-Lipid system. The 1 mL of the sample was analyzed by Agilent 6545 Series Q-TOF LC/MS (LCMS-QTOF). A 20 μL of sample was injected onto the Poroshell 120 EC-C18 2.7 μm (100×2.1 mm i.d.) column. Chromatographic separation analysis was achieved in 60% aqueous acetonitrile containing 0.1% formic acid over 30 min at a flow rate of 0.4 mL min–1. Electrospray mass spectra were conducted in the negative ionization mode [M–H–]. The MS/MS data used the Agilent mass hunter qualitative analysis workflow program26.
Biofilm formation: The bacterial strains were inoculated into LB medium and grown overnight at 180 rpm for 16 hrs at 37°C. For colony formation, 2 μL was spotted onto LB, LBM (0.1 mM MnSO4·H2O add to LB), LBG (1% glycerol add to LB) and LBGM (0.1 mM MnSO4·H2O and 1% glycerol add to LB) solidified with 1.5% agar. Plates were incubated at 30°C and were imaged after 3 days27,28.
Statistical analysis: The study was conducted using a Complete Randomized Design (CRD). The analysis of variance showed significance at an alpha level of 0.05. For the basic statistical analysis of data such as means, standard deviation and standard curve the MS Excel 2010 was used.
RESULTS
Bacterial strain and phenotypic characterization: The Bacillus spp. strains BB06, BB10, CF58 and CR74 were characterized morphologically and biochemically by following Bergey’s Manual of Systematic Bacteriology. Four strains were identified as gram-positive and spore-forming rods measuring>1 μm width. The isolates were motile and tested positive for catalase activity, starch hydrolysis, citrate utilization, growth on 6.5% NaCl medium at room temperature but did not grow on 6.5% NaCl medium at 55°C (Table 1). The isolates were found to be in the Bacillus subtilis group. All Bacillus strains, including four B. velezensis strains BB06, BB10, CF58 and CR74, demonstrated positive reactions in the hydrolytic enzyme production test.
| Table 1: | Detail of morphological, physiological, plant growth-promoting bacteria and biocontrol potentials of rhizobacteria isolated from the rhizosphere |
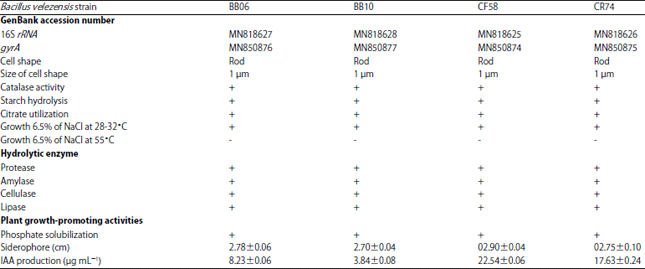 | |
Values were expressed as the Mean±Standard deviation of four replicates, rRNA: Ribosomal ribonucleic acid, gyrA: Subunit A of DNA gyrase, >: Greater than, +: Positive reaction, −: Negative reaction | |
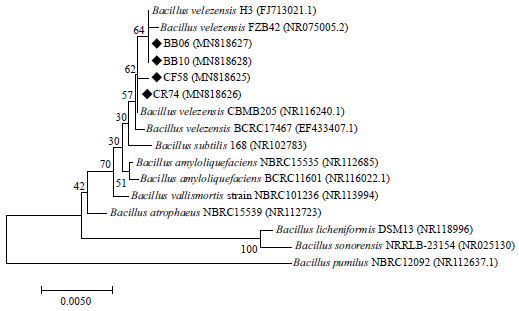 | |
| Fig. 1: | Neighbor-joining phylogenetic tree of the 16S rRNA gene of strains CF58, CR74, BB06 and BB10 based on available reference sequences from the Genbank NCBI database |
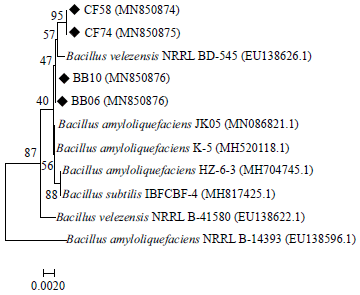 | |
| Fig. 2: | Neighbour-joining phylogenetic tree of gyrA gene sequences of strains CF58, CR74, BB06 and BB10. Bootstrap values are shown as fractions out of 1000 replicates |
All enzyme production test results (protease, amylase, cellulase and lipase) are shown in Table 1. While, all four strains were positive for siderophore production, phosphate solubilization and IAA production which ranged from 3.84 to 22.54 μg mL–1.
Furthermore, 16S rRNA sequencing identified all the isolates (BB06, BB10, CF58 and CR74) as Bacillus velezensis(Accession numbers MN818627, MN818628, MN818625 and MN818626, respectively). Strains BB06 and BB10 clustered closely with B. velezensis strain H3 (FJ713021.1) and strain FZB42 (NR075005.2). Strains CF58 and CR74 clustered closely with B. velezensis strain BCRC17467 (NR116240.1) (Fig. 1).
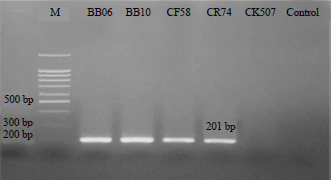 | |
| Fig. 3: | Gel electrophoresis of PCR products for the detection of surfactin antibiotic genes. |
Lane M = 100 bp+1.5 kb DNA Ladder, Lane 1 = B. velezensis strains CF58, Lane 2 = B. velezensis strains CR74, Lane 3 = B. velezensis strains BB06, Lane 4 = B. velezensis strains BB10, Lane 5 = B. altitudinis strain CK507, Lane 6 = Negative control without DNA (distilled water) | |
The gyrA gene coding for DNA gyrase subunit A was selected as an alternative phylogenetic marker of the Bacillus subtilis group. The gyrA gene sequence of all isolates (BB06, BB10, CF58 and CR74) was identified to be that of B. velezensis (Accession numbers MN850876, MN850877, MN850874 and MN850875, respectively) and clustered closely with B. velezensis strain NRRL BD-545 (EU138626.1) (Fig. 2). Bacillus velezensis, B. amyloliquefaciens and B. subtilis are close relatives.
Bacillus strains, including the four B. velezensis strains BB06, BB10, CF58 and CR74 effectively inhibited the growth of bacterial pathogens of potato. All strains showed a broad inhibitory spectrum and produced zones of inhibition against P. carotovorum subsp. brasiliense strain PY1 (2.36-2.93 cm2), D. dadantii strain CK211 (2.54-3.46 cm2), B. altitudinis strain CK507 (3.80-4.90 cm2) and R. solanacearum strain CD1 (1.77-2.65 cm2) (Table 2).
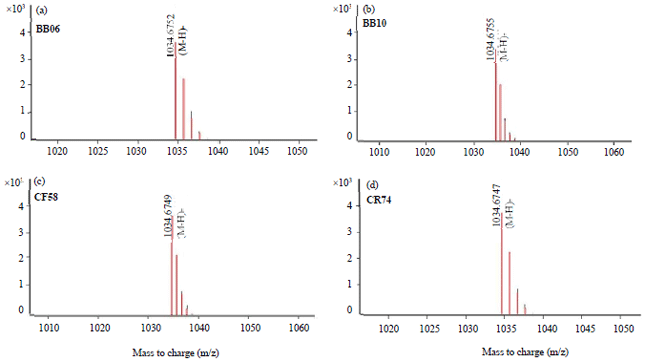 | |
| Fig. 4(a-d): | LCMS-QTOF analysis of the surfactin produced by B. velezensis strain BB06, BB10, CF58 and CR74 |
(a) LCMS-QTOF analysis of the surfactin eluted at 27.90 min, (b) 27.40 min, (c) 27.59 min and (d) 27.18 min. | |
| Table 2: | Growth inhibition of different pathogenic bacteria of potato by B. velezensis strains |
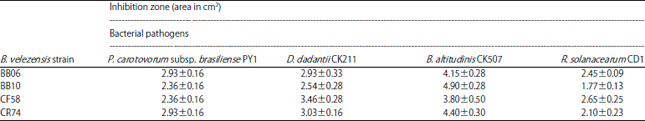 | |
| Values were expressed as the Mean±Standard deviation of three replicates | |
PCR amplification using the primer set srfAA for the surfactin gene, showed that all strains of B. velezensis amplified a 201 bp segment while B. altitudinis strain CK507 and the negative control (distilled water) produced no PCR product (Fig. 3). When the sequences were deposited in the GenBank database and analyzed using a BLASTX algorithm-based program, the protein “HSDTEGLIGMFVNTLALRSSV KQDQTFAGLLGHVRKQVL” clustered at 100% with surfactin non-ribosomal peptide synthetase SrfAA of B. velezensis NJN-6, B. velezensis, B. subtilis, B. amyloliquefaciens, Bacillus sp. LYLB3 and Bacillus sp. BK100 (accession numbers AKD28464.1, WP138118307.1, WP072588783.1, WP015388716.1, WP094032059.1 and WP132106744.1, respectively). B. velezensis and B. altitudinis were analyzed by LCMS-QTOF. The data analysis of the surfactin antibiotic of B. velezensis strains BB06, BB10, CF58 and CR74 revealed the characteristic molecular ions mode [M–H–] (m/z 1034.6752, 1034.6755, 1034.6749 and 1034.6747, respectively) (Fig. 4a-d). However, B. altitudinis strain CK507 did not show surfactin antibiotic production. The surfactin peaks were observed at 27.09, 27.40, 27.59 and 27.18 min, respectively.
Bacillus sp. depends on the ability to form biofilms, the production of a matrix protects the bacteria from environmental stress. The study found that a combination of glycerol and manganese strongly promotes biofilm formation in the LBGM medium. Therefore, B. velezensis strains BB06, BB10, CF58 and CR74 showed robust biofilm formation (wrinkled pellicles) but another species, B. altitudinis strain CK507 did not show biofilm formation.
 | |
| Fig. 5: | A combination of 0.1 mM of manganese and 1% of glycerol added to LB medium on B. velezensis strain BB06, BB10, CF58, CR74 and B. altitudinis strain CK507 promotes robust biofilm formation after incubation for 3 days |
The strain BB06 showed especially strong biofilm formation on the LBGM medium. Both components 1% glycerol and 0.1 mM MnSO4·H2O alone stimulated relatively little biofilm formation (Fig. 5).
DISCUSSION
The native valuable rhizosphere bacteria compose the immediate protection against the pathogens attacking plants. Several researchers have shown the biocontrol potential of the B. subtilis species complex through the production of antibiotics and secondary metabolites. PGPR are bacteria that are known to generate growth factors, hormones and vitamins that improve plant growth at the same time increasing plant yield29. The present study evaluated four strains of Bacillus spp. that were previously isolated from the rhizosphere. Based on the 16S rRNA and gyrA genes these bacteria were identified as B. velezensis strains BB06, BB10, CF58 and CR74. All strains showed a broad inhibitory spectrum and produced zones of inhibition against bacterial pathogens of potato including P. carotovorum subsp. brasiliense PY1, D. dadantii CK221, B. altitudinis CK507 and R. solanacearum CD1, which ranged from 1.77-4.90 cm2. Bacillus species inhibited R. solanacearum race 1, bv 3, phylotype I under in vitro environments. B. amyloliquefaciens DSBA-11 presented the highest growth repression of R. solanacearum (4.91 cm2) followed by strains DSBA-12 (3.31 cm2) and B. subtilis (3.07 cm2)30. The injection of P. carotovorum subsp. carotovorum EMPCC instigated extensive tissue softening in 48 hrs, whereas the capability of EMPCC to soften potato rhizome tissues was condensed when co-injected with Bacillus sp. DMA13331. Bacillus subtilis, B. thuringiensis, B. megaterium, B. cereus and B. pumilus showed good antagonistic action by producing regions of suppression against P. carotovorum subsp. carotovorum, a pivotal medium in blackleg infection of potato32. Besides, biocontrol activities directed at the bacterial soft rot infection of potato have been reported from several bacterial competitors from the Bacillus genus33,34. Furthermore, the antagonistic activity and efficacy of any biocontrol agents depend essentially upon the production of the precise combination of secondary metabolites active against pathogens.
Current study of bacterial hydrolytic enzyme production showed that all isolates can produce protease, amylase, cellulase and lipase, which is similar to the findings of many researches7. Bacillus species showed the ability to produce hydrolytic enzymes (protease, chitinase and cellulase. Plant growth promotion analysis showed that B. velezensis in this study produced siderophores, phosphate solubilization and IAA production which ranged from 3.84-22.54 μg mL–1. These findings are in congruence that extracted seven similar antagonistic bacterial strains from surface-sterilized healthy roots of sweet potato. Antagonistic bacteria produced IAA that spanned from 0.6-10.73 μg mL–1 and solubilized phosphate22. There have been many reports of antagonistic bacteria that produce hydrolytic enzymes and act as growth promoters and biocontrol agents against plant pathogens. Similarly, 19 isolates of bacteria from various soils that produced phosphate solubilization35.
The antagonistic activity of the B. subtilis species complex against plant pathogens is related to the synthesis of lipopeptide-type compounds such as fengycin, iturin and surfactin families36. Surfactin is a very great surfactant frequently utilized as an antibiotic. Uniquely, surfactin is an antibiotic formed by the Gram-positive bacteria that forms endospore37. Thus, surfactin was found to exhibit effective antibacterial, antiviral and antifungal activity. The four strains of B. velezensis evaluated in this study showed surfactin antibiotic production when the characteristic molecular ions were detected by LCMS-QTOP. The supernatants of bacteria were observed that the lipopeptide until after 3 days of growth38. The lipopeptide surfactins are the primary determinants of biocontrol and aggregation in biofilms9. Surfactin is known to stimulate biofilm formation. The B. velezensis strains used in this study showed robust analogous biofilm formation LBGM medium. These results are similar to those described that the LBGM medium stimulates the vigorous biofilm making of B. subtilis NCIB361028. The medium also arouses biofilm-linked spore formation. Biofilm formation is important in the colonization of plant roots and protecting plans against infection by pathogens. Moreover, B. velezensis demonstrated biocontrol efficacy.
CONCLUSION
B. velezensis strains BB06, BB10, CF58 and CR74 effectively inhibited the growth of bacterial pathogens of potato. This study identified biocontrol agents with plant growth promotion and hydrolytic production capabilities. The bacteria showed the ability to promote plant growth by producing IAA. They formed biofilms and produced a surfactant antibiotic when analyzed through LCMS-QTOP.
SIGNIFICANCE STATEMENT
B. velezensis strains BB06, BB10, CF58 and CR74 were identified by 16S rRNA gene and gyrA gene and showed biocontrol agents with plant growth promotion and hydrolytic production capabilities. Four strains have the ability to promote plant growth by producing IAA and produced a surfactin antibiotic. They effectively inhibited the growth of bacterial pathogens of potato. These strains can be recommended as potential biocontrols for reducing bacterial plant pathogens of potato, application of pesticides and chemical fertilizers and supporting sustainable agriculture.
ACKNOWLEDGMENT
This research was partially supported by Innovation Agriculture Research Center, Faculty of Agriculture, Chiang Mai University and Center of Excellence on Agricultural Biotechnology: (AG-BIO/PERDO-CHE), Bangkok, Thailand.
REFERENCES
- Piggot, P.J. and D.W. Hilbert, 2004. Sporulation of Bacillus subtilis. Curr. Opin. Microbiol., 7: 579-586.
CrossRefDirect Link - Bhattacharyya, P.N. and D.K. Jha, 2012. Plant Growth-Promoting Rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol., 28: 1327-1350.
CrossRefDirect Link - Vessey, J.K., 2003. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil, 255: 571-586.
CrossRefDirect Link - Somers, E., L. Vanderleyden and M. Srinivasan, 2004. Rhizosphere bacterial signalling: A love parade beneath our feet. Crit. Rev. Microbiol., 30: 205-240.
CrossRefPubMedDirect Link - Abd-El-Khair, H. and I.H. Seif-El-Nasr, 2012. Applications of Bacillus subtilis and Trichoderma spp. for controlling the potato brown rot in field. Arch. Phytopathol. Plant Prot., 45: 1-15.
CrossRefDirect Link - Cui, W., P. He, S. Munir, P. He and Y. He et al., 2019. Biocontrol of soft rot of chinese cabbage using an endophytic bacterial strain. Front. Microbiol., Vol. 10.
CrossRefDirect Link - Aleti, G., S. Lehner, M. Bacher, S. Compant, B. Nikolic et al., 2016. Surfactin variants mediate species-specific biofilm formation and root colonization in Bacillus Environ. Microbiol., 18: 2634-2645.
CrossRefDirect Link - Solanki, M.K., R.K. Singh, S. Srivastava, S. Kumar, P.L. Kashyap and A.K. Srivastava, 2015. Characterization of antagonistic-potential of Bacillus velezensis SK71 against bacterial brown spot on a terrestrial orchid (Habenaria lindleyana). J. Basic Microbiol., 55: 82-90.
CrossRefDirect Link - Kearns, D.B., F. Chu, R. Rudner and R. Losick, 2004. Genes governing swarmimg in Bacillus subtilis and evidence for phase variation mechanism controlling surface motility. Mol. Microbiol., 52: 357-369.
CrossRefDirect Link - Zeigler, D.R., Z. Prágai, S. Rodriguez, B. Chevreux and A. Muffler et al., 2008. The origins of 168, W23, and other Bacillus subtilis legacy strains. J. Bacteriol., 190: 6983-6995.
CrossRefDirect Link - Christensen, H., S. Nordentoft and J.E. Olsen, 1998. Phylogenetic relationships of Salmonella based on rRNA sequences. Int. J. Syst. Bacteriol., 48: 605-610.
CrossRefDirect Link - Chun, J. and K.S. Bae, 2000. Phylogenetic analysis of Bacillus subtilis and related taxa based on partial gyrA gene sequences. Kluwer Academic Publishers, 78: 123-127.
CrossRefDirect Link - Joung, K.B. and J.C. Cote, 2002. Evaluation of ribosomal RNA gene restriction patterns for the classification of Bacillus species and related genera. J. Applied Microbiol., 92: 97-108.
CrossRef - Sneath, P.H.A., 1986. Bergey's Manual of Systematic Bacteriology. Vol. 2, Williams and Wilkins, Baltimore, ISBN: 9781118960608,.
CrossRef - Cheng, H.R. and N. Jiang, 2006. Extremely rapid extraction of DNA from bacteria and yeasts. Biotechnol. Lett., 28: 55-59.
CrossRefDirect Link - Kumvinit, A. and A. Akarapisan, 2018. Evaluation of Bacillus amyloliquefaciens for control of potato blackleg caused by Pectobacterium carotovorum. J. Appl. Sci.
CrossRefDirect Link - Song, Z., K.Q. Liu, Q.X. Lu, J. Yu, R.C. Ju and X.L. Liu, 2011. Isolation and characterization of a potential biocontrol Brevibacillus laterosporus Afr. J. Microbiol. Res., 5: 2675-2681.
CrossRefDirect Link - Kumvinit, A. and A. Akarapisan, 2019. Characterization of blackleg and soft rot from potato in northern Thailand. J. Phytopathol., 167: 655-666.
CrossRefDirect Link - Nielsen, P. and J. Sorensen, 1997. Multi-target and medium independent fungal antagonism by hydrolytic enzymes in Paenibacillus polymyxa and Bacillus pumilus strains from barley rhizosphere. FEMS Microbiol. Ecol., 22: 183-192.
CrossRef - Silfkin, M., 2000. Tween 80 opacity test responses of various Candida species. J. Clin. Microbiol., 38: 4626-4628.
CrossRefDirect Link - Dawwam, G.E., A. Elbeltagy, H.M. Emara, I.H. Abbas and M.H. Mohamed, 2013. Beneficial effect of plant growth promoting bacteria isolated from the roots of potato plant. Ann. Agric. Sci., 58: 195-201.
CrossRefDirect Link - Schwyn, B. and J.B. Neilands, 1987. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem., 160: 47-56.
CrossRefPubMedDirect Link - Joshi, R. and B.B.M. Gardener, 2006. Identification and characterization of novel genetic markers associated with biological control activities in Bacillus subtilis. Phytopathology, 96: 145-154.
CrossRef - Moran, S., K. Robertson, F. Paradisi, K.D. Rai and D.C. Murphy, 2010. Production of lipopeptides in Bacillus sp. CS93 isolated from Pozol. FEMS Microbiol. Lett., 304: 69-73.
CrossRefDirect Link - Devi, S., T.H. Kiesewalter, R. Kovács, C. Frisvad and T. Weber et al., 2019. Depiction of secondary metabolites and antifungal activity of Bacillus velezensis DTU001. Synth. Syst. Biotechnol., 4: 142-149.
CrossRefDirect Link - M. Shemesh and Y. Chai, 2013. A combination of glycerol and manganese promotes biofilm formation in Bacillus subtilis via histidine kinase KinD signaling. J. Bacteriol., 195: 2747-2754.
CrossRefDirect Link - Sansinenea, E. and A. Ortiz, 2011. Secondary metabolites of soil Bacillus spp. Biotechnol. Lett., 33: 1523-1538.
CrossRefDirect Link - Singh, D., D.K. Yadav, G. Chaudhary, V.S. Rana and R.K. Sharma, 2016. Potential of Bacillus amyloliquefaciens for biocontrol of bacterial wilt of tomato incited by Ralstonia solanacearum. J. Plant Pathol. Microbiol., Vol. 7.
CrossRef - Esmaeil, M., E.S.T. Badraldin and V. Vittorio, 2011. Virulence attenuation of Pectobacterium carotovorum using N-Acyl-homoserine Lactone degrading bacteria isolated from potato rhizosphere. Plant Pathol. J., 27: 242-248.
CrossRefDirect Link - Issazadeh, K., S.K. Rad, S. Zarrabi and M.R. Rahimibashar, 2012. Antagonism of Bacillus species against Xanthomonas campestris pv. campestris and Pectobacterium carotovorum subsp. carotovorum. Afr. J. Microbiol. Res., 6: 1615-1620.
CrossRefDirect Link - Czajkowski, R., W.J. de Boer, J.A. van Veen and J.M. van der Wolf, 2011. Characterization of bacterial isolates from rotting potato tuber tissue showing antagonism to Dickeya sp. biovar 3 in vitro and in planta. Plant. Pathol., 61: 169-182.
CrossRefDirect Link - Rahman, M.M., M.E. Ali, A.A. Khan, A.M. Akanda, M.K. Uddin, U. Hashim and S.B. Abd Hamid, 2012. Isolation, characterization and identification of biological control agent for potato soft rot in Bangladesh. Scient. World J.
CrossRefDirect Link - Park, J., N. Bolan, M. Mallavarapu and R. Nai, 2010. Enhancing the solubility of insoluble phosphorus compounds by phosphate solubilizing bacteria. 19th World Congree of Soil Science, Soil Solution for a Changing World, 1-6 August, Brisbane, Australia, pp: 65-68.
Direct Link - Chen, X.H., A. Koumoutsi, R. Scholz, K. Schneider and J. Vater et al., 2009. Genome analysis of Bacillus amyloliquefaciens FZB42 reveals its potential for biocontrol of plant pathogens. J. Biotechnol., 140: 27-37.
CrossRefDirect Link - Peypoux, F., J.M. Bonmatin and J. Wallach, 1999. Recent trends in the biochemistry of surfactin. Applied Microbiol. Biotechnol., 51: 553-563.
CrossRefPubMedDirect Link - Phister, T.G., D.J. O'Sullivan and L.L. McKay, 2004. Identification of bacilysin, chlorotetaine and iturin a produced by Bacillus sp. Strain CS93 isolated from pozol, a Mexican fermented maize dough. Appl. Environ. Microbiol., 70: 631-634.
CrossRefDirect Link