
Angsana Akarapisan
Division of Plant Pathology, Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, 50200 Chiang Mai, Thailand
LiveDNA: 66.16079
Athidtaya Kumvinit
Division of Plant Pathology, Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, 50200 Chiang Mai, Thailand
Soraya Ruamrungsri
H.M. The King`s Initiative Centre for Flower and Fruit Propagation, 50230 Chiang Mai, Thailand
Asian Journal of Plant Pathology
Year: 2017 | Volume: 11 | Issue: 2 | Page No.: 95-101







ABSTRACT
Background and Objective: Amaryllis production has dropped due to bulb rot in the cropping areas of Chiang Mai, Thailand. It has become a recurrent problem of amaryllis bulb production. It is very difficult to develop effective management strategies for soil-borne pathogens that produce sclerotia because of their long-term survival in the soil. Fungicides are among the most effective options for the control of diseases caused by soil-borne pathogens. The objectives of this study were to identify the pathogen of amaryllis bulb rot and to evaluate fungicides for their efficacy against the pathogen. Methodology: The experiments identified the pathogen based on morphological characteristics and PCR analysis using specific primers. The pathogenicity of fungal isolates was tested in the greenhouse and compared with the non-treated control. Five fungicides including captan, copper oxychloride, mancozeb, carbendazim and etridiazole+quintozene were tested by the poison agar method, this analysis was arranged in a completely randomized design with 5 replicates of each treatment. Results: The initial disease symptom was water-soaked lesions in bulbs followed by total rotting of bulbs and leaf sheath blight. White cottony mycelia formed on the blubs of diseased plants. Clamp connections were common at every septum. The isolate was identified as Sclerotium rolfsii. A pathogenicity test was conducted in the greenhouse, where the symptoms of leaf sheath blight were observed. Also, bulb fresh weight, circumference and number of bulblets were decreased after pathogen inoculation. Effective and efficient integrated management of crop diseases often includes the use of fungicides. At recommended doses, etridiazole+quintozene, captan and mancozeb significantly reduced the growth of the fungus by 100, 68.85 and 64.56%, respectively. Conclusion: The results revealed that S. rolfsii was the causal agent of bulb rot of amaryllis in Northern Thailand. The most effective fungicide treatment against S. rolfsii from rotted amaryllis bulbs was etridiazole+quintozene.
PDF Abstract XML References Citation
Received: February 14, 2017;
Accepted: March 09, 2017;
Published: March 15, 2017
How to cite this article
Angsana Akarapisan, Athidtaya Kumvinit and Soraya Ruamrungsri, 2017. Bulb Rot of Amaryllis Caused by Sclerotium rolfsii and Effect of Fungicides on in vitro Inhibition of Mycelial Growth. Asian Journal of Plant Pathology, 11: 95-101.
URL: https://scialert.net/abstract/?doi=ajppaj.2017.95.101
URL: https://scialert.net/abstract/?doi=ajppaj.2017.95.101
INTRODUCTION
Amaryllis (Hippeastrum × hybridum) belongs to the Amaryllidaceae family and has been economically exploited as an ornamental plant. It is also known as ‘naked lily’ because of its lovely trumpet-shaped flowers and is indigenous to Southern Africa1. Amaryllis is a popular plant grown in gardens for beautiful blooms of various colours in the Netherlands, the USA and South Africa and is now emerging as an important cut flower for the floriculture industry in India. Amaryllis bulbs are available from North America, the Netherlands, Israel, Brazil and South Africa2. Hippeastrum is a genus of about 90 species and 600 hybrids and cultivars, the first hybrids were made in the Netherlands3. In recent years, amaryllis bulb production in Thailand for exports markets has grown rapidly and its contract farming has also increased.
Sclerotium rolfsii is a soil-borne plant pathogen causing disease on a wide range of agricultural crops in the Southern United States, West Indies, Southern Europe, Central and South America, countries bordering the Mediterranean, Africa, Hawaii, Japan, Philippines and India. The fungus attacks more than 500 plants species in ca. 100 families4-6. Sclerotium rolfsii is characterized by white fluffy, branched, septate mycelia and small, round, brown sclerotia7. The sclerotia of S. rolfsii are 0.5-1.99 mm in diameter and are most often found around the base and on underground storage organs of the host plant. Primary hyphae have clamp connections at the septa8,9.
Management of plant diseases is focused on reducing the economic and aesthetic damage caused by pathogens. Use of fungicides is a common component of integrated disease management. Many fungicides have been used for the control of diseases caused by soil-borne pathogens7,10. Mycelial growth of S. rolfsii can be reduced by several fungicides11. One systemic (carbendazim), three contacts (mancozeb, copper oxychloride and chlorothalonil) and three combinations of systemic and contact fungicides (carboxin+thiram, metalaxyl+mancozeb, cymoxanil+mancozeb) were evaluated against S. rolfsii in the laboratory4. The fungicides thiram, quintozene, captan, carbendazim, benomyl, oxycarboxin and triadimenol have been used to control S. rolfsii in several crops. Therefore, the main purposes of this study were to identify the pathogen of amaryllis bulb rot and to evaluate fungicides for their efficacy against the pathogen in vitro.
MATERIALS AND METHODS
Fungal isolates: The causal fungus was isolated from the sclerotia that formed on the bulb of the infected amaryllis from San Kamphaeng District in Chiang Mai, Thailand in 2015. The sclerotia that had formed on the bulb of the amaryllis were disinfected with a 1% sodium hypochlorite solution for 3 min and rinsed with sterilized water two times. The surface-sterilized sclerotia were placed on Potato Dextrose Agar (PDA) and incubated at room temperature (28±4°C) for 4 days. The hyphal tips of fresh mycelia growing from sclerotia were cut and transferred to new PDA plates and incubated at room temperature. Morphological characteristic such as: mycelium condition and color, presence of hyphal clamp connection and fungal growth were noted12.
Pathogenicity test and determining yield losses: The pathogenicity of the fungus was tested in the greenhouse on ‘Apple blossom’ and ‘Christmas gift’ amaryllis in pots. The test plants were inoculated with the isolated fungus using the following procedures. The fungal inoculum was produced in 200 g of sorghum in a plastic bag which was autoclaved at 121°C for 30 min. The mycelia from 10 discs were mixed with the sorghum and incubated at room temperature for 7 days. The sorghum infested with the fungal mycelia was thoroughly mixed with 3 kg of soil which were used to fill 30 cm diameter pots. One amaryllis bulb was placed in each pot and a CRD design with four replications was used. The ‘Apple blossom’ and ‘Christmas gift’ amaryllis without the pathogen served as controls, plant were grown in greenhouse and watered as required. The experiment ended after 6 months and the bulbs were lifted and their roots removed. All bulblets were counted and then removed. Bulbs were weighed and their circumferences measured.
Effect of temperature on radial growth: A culture of the fungal isolate from infected amaryllis was used. The linear hyphal extension rate (mm day–1) of the fungal isolate was determined between 5 and 35°C on PDA in 9 cm diameter Petri plates. A sterile cork-borer (5 mm diameter) was used to remove an agar disc containing hyphae from the edge of an actively growing culture. The experimental design was a CRD with 5 replications. The daily radial fungal growth rate (mm day–1) was calculated.
PCR amplification
DNA extraction: Fungal isolates were cultured in Potato Dextrose Broth (PDB) at room temperature (30±2°C) for 7 days. DNA was extracted from the isolate using a modification of the protocol of Stewart and Via12. About 0.5 g of mycelia of the fungal isolate were ground with 12 μL of 2 mercaptoethanol and 2.4 mL of DNA extraction buffer (2% w/v CTAB, 1.42 M NaCl, 20 mM EDTA, 2% w/v polyvinylpyrrolidone, 5 mM citric acid and 100 mM Tris-HCl pH 8.0).
| Table 1: | Active ingredients and chemical groups of the fungicides15 |
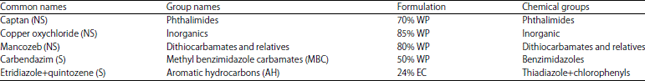 | |
| *NS: Non-systemic fungicide, S: Systemic fungicide, WP: Wettable powder, EC: Emulsifiable concentrate | |
Approximately 500 μL of the extraction solution were removed into a sterile 1.5 mL tube. Then 500 μL of a chloroform and isoamyl alcohol (24:1) solution were added and mixed. The solution was centrifuged at room temperature at 5000 rpm for 5 min. The upper aqueous phase containing the DNA was removed into a new sterile 1.5 mL tube. The genomic DNA was precipitated using 0.7X isopropanol by centrifugation at room temperature at 14,000 rpm for 20 min. Genomic DNA in TE buffer was visualized in a 1% (w/v) agarose gel after ethidium bromide staining. The DNA concentration was determined by gel electrophoresis13.
PCR amplification: The DNA amplification was performed by Polymerase Chain Reaction (PCR). The specific primer pair for detection of S. rolfsii included the SCR-F primer (5'-CGTAGGTGAACCTGCGGA-3') coupled with the SCR-R primer (‘5-CATACAAGCTA GAATCCC-3'), used to amplify a 540 bp product13. Sclerotium rolfsii specific PCR reactions were performed in a total volume of 25 μL, containing 50 ng of genomic DNA, 10X of PCR buffer, 10 mmol L–1 of each dNTP, 50 mmol L–1 MgCl2, 1 U of Taq DNA polymerase and 0.5 mmol L–1 of each primer. The reaction mixtures were incubated in a Peltier-based Thermal Cycler A100/A200 (LongGene Document Version 1.4). Following an initial denaturation at 94°C for 2 min, the DNA templates were amplified for 35 cycles consisting of 30 sec at 94°C,1 min at 58°C and 1.5 min at 72°C followed by a final extension at 72°C for 8 min. Amplification products were separated in 1% (w/v) agarose gels electrophoresis and viewed under UV light14.
In vitro fungicide bioassay: The isolate of Sclerotium sp. was maintained on PDA slant tubes at room temperature. For the in vitro fungicide bioassay, 5 mm diameter mycelial plugs taken from the edges of actively growing PDA cultures were inverted in the centers of PDA plates amended with fungicides to determine the effect of each fungicide on mycelial growth. Fungicides viz., captan, copper oxychloride, mancozeb, carbendazim and etridiazole+quintozene suspended in sterile distilled water were added to cooled, autoclaved PDA to give the desired concentration of the recommended dose, a ½ dose and a ¾ dose. The PDA without fungicide served as the control. Active ingredients and chemical groups of the fungicides are given in Table 1. There were five replicates of each treatment arranged in a CRD. The percent inhibition of mycelial growth was determined.
Statistical analysis: The study was conducted using a Complete Randomized Design (CRD). The data were subjected to statistical analysis using the Least Significant Different (LSD), the analysis of variance showed significance at an alpha level of 0.05.
RESULTS
Fungal isolates: The occurrence of disease in the bulb and leaves of ‘Apple blossom’ amaryllis was observed in the field. The fungus infects the surface of bulbs resulting in development of lesions. Numerous small white and brown sclerotial growths were observed on amaryllis bulbs, as well as blight along the leaf sheaths (Fig. 1a-b). The sclerotia were nearly round, 1-2 mm in diameter and initially white turning brown (Fig. 1c). The hyphae are 3.75-8.50 μm diameter. Primary hyphae showed clamp connections at the septa (Fig. 1d).
Pathogenicity test and determining yield losses: The pathogenicity test was conducted on ‘Christmas gift’ and ‘Apple blossom’ amaryllis in the greenhouse. White mycelia has started to form on the bulbs with sclerotia at 7 days after pathogen inoculation. After that, the sclerotial primordia developed into brownish sclerotia. Bulb rot symptoms were observed after the mycelium covered the bulb. Yield losses were caused by S. rolfsii in amaryllis 6 months after pathogen inoculation. The symptoms observed included blight along the leaf sheaths. Bulb fresh weight and circumference and number of bulblets were also significantly decreased by S. rolfsii infection compared to the non-treated control (Fig. 2, 3).
 | |
| Fig. 1(a-d): | Bulb rot and leaf blight of amaryllis caused by Sclerotium rolfsii (a, b) Symptoms on leaves and bulb of amaryllis, (c) Mycelium growth and sclerotium on potato dextrose agar and (d) Clamp connection formed on the hyphae Scale bar = 10 μm |
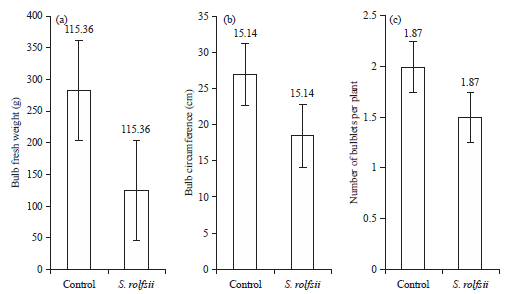 | |
| Fig. 2(a-c): | Effect of S. rolfsii on bulb quality of ‘Christmas gift’ amaryllis plants at 6 months after pathogen inoculation (a) Bulb fresh weight, (b) Bulb circumference and (c) Number of bulblets per plant |
Effect of temperature on radial growth: Temperature significantly affected the linear hyphal extension rate of S. rolfsii. Fungal colonies grew at 10-35°C with a maximum extension rate of 18.1 mm day–1 at 30°C. The isolate of S. rolfsii from amaryllis failed to grow at 5°C. The temperatures favouring the growth of the pathogen and infection of ‘Apple blossom’ amaryllis by S. rolfsii are 25-30°C.
PCR amplification: The isolate of Sclerotium sp. from ‘Apple blossom’ amaryllis was identified by using the SCR-F primer ('5-CGTAGGTGAACCTGCGGA-3') coupled with the primer SCR-R ('5-CATACAAGCTAGAATCCC-3'). This species-specific PCR confirmed that the isolate of Sclerotium sp. from ‘Apple blossom’ amaryllis was Sclerotium rolfsii, which was consistent with the morphological identification. A fragment of about a 540 bp was amplified from the genomic DNA of the S. rolfsii isolate (Fig. 4).
In vitro fungicide bioassay: Mycelium of S. rolfsii covered the petri plate surface after 48 h of incubation on unamended PDA.
| Table 2: | Inhibition in growth of Sclerotium rolfsii on potato dextrose agar containing different concentrations of fungicides |
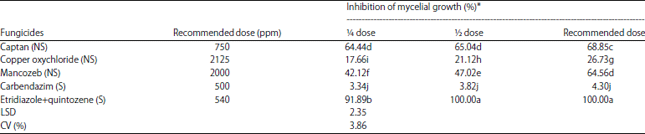 | |
*Values within a column followed by different letters are significantly different (p<0.05) by LSD. The experimental design was a Completely Randomized Design (CRD) NS: Non-systemic fungicide, S: Systemic fungicide | |
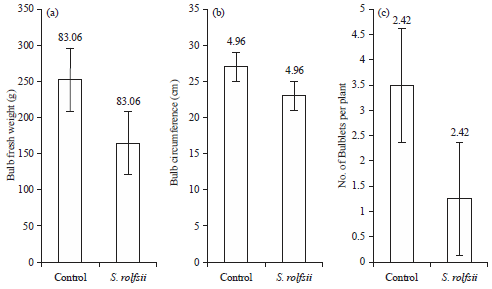 | |
| Fig. 3(a-c): | Effect of S. rolfsii on bulb quality of ‘Apple blossom’ amaryllis plants at 6 months after pathogen inoculation (a) Bulb fresh weight, (b) Bulb circumference and (c) Number of bulblets per plant |
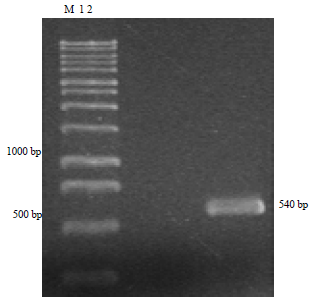 | |
| Fig. 4: | PCR amplification of a specific fragment from Sclerotium rolfsii |
| Lane M: Molecular ladder, Lane 1: Distilled water, Lane 2: Sclerotium rolfsii from ‘Apple blossom’ amaryllis | |
Mycelial growth of S. rolfsii was markedly suppressed on media amended with etridiazole+quintozene, captan, mancozeb and copper oxychloride (Table 2). Increased fungicide concentrations resulted in corresponding reductions in mycelial growth. Etridiazole+quintozene was found to be the most effective treatment and gave 100% reduction in the growth of S. rolfsii when used at the recommended and ½ doses. At ¼ of the recommended dose a 91.89% reduction of mycelial growth was achieved. Captan and mancozeb at the recommended doses were found to be effective in inhibiting the radial growth of S. rolfsii by 68.85 and 64.56%, respectively. Carbendazim showed the lowest inhibition (4.30%) of mycelial growth at the recommended dose compared to the other fungicides tested.
DISCUSSION
A bulb rot and sheath blight disease on ‘Apple blossom’ amaryllis was recently noticed causing major crop loss in some commercial fields in San Kamphaeng District in Chiang Mai, Thailand. The initial symptoms were water-soaked lesions in bulbs followed by total bulb rot. White cottony mycelia formed on the bulb and blight occurred along the leaf sheaths, on which small white and brown sclerotia formed. The hyphal diameter was approximately from 3.75 to 8.50 μm. Therefore, the isolate was identified as Sclerotium rolfsii and confirmed by a species-specific PCR. The optimum temperature for growth rate (18.1 mm day–1) was 30°C on PDA.
The pathogen causes significant crop damage worldwide. Similar observations were made by Kwon12, who reported that S. rolfsii was found in garlic cultivated from 2008-2010, in Gyeongnam province, Korea. Early symptoms of this disease were water-soaked spots on bulbs, which were followed by rot, wilt, blight and death. White mycelia spread over the lesions near the soil and sclerotia formed on the stem. Hyphal diameter ranged from 4-8 μm. Sclerotia were round, tan to brown and ranged from 1-3 mm in diameter. Han et al.8 reported that a stem rot disease found in garlic and Cymbidium orchids in Korea, was caused by S. rolfsii (Saccardo). Jeeva et al.14 identified S. rolfsii by PCR analyses using specific primers. A species specific PCR performed by using two primers: SCR-F and SCR-R, amplified a 540 bp product from S. rolfsii DNA isolated from the plant Amorphophallus paeoniifolius.
The current research results showed that the fungicide combination of etridiazole+quintozene was the most effective and gave a 100% reduction in the radial growth of S. rolfsii on PDA when used at the recommended and half-doses. Captan and mancozeb were found to be the effective in inhibiting the radial growth of S. rolfsii at the recommended doses, producing growth inhibitions of 68.85 and 64.56%, respectively. Keyser and Ferreira16 found that quintozene as a dip and soil drench, effectively protected plants against S. rolfsii. Using quintozene before planting in the nursery can be a practical way of controlling the disease where S. rolfsii is a problem. Yaqub and Shahzad17 reported that at 100 ppm Dithane M-45 (mancozeb) showed a 60% inhibition in mycelial growth of S. rolfsii. Similarly, Dithane M-45 at 200 ppm reduced the growth of S. rolfsii18. Captan, chlorothalonil, mancozeb and pentachloronitrobenzene in agar reduced germination of sclerotia by 80-95%19. Mancozeb at the highest concentration of 400 ppm produced an 80.36% radial growth inhibition of S. rolfsii20. Thiophanate methyl, metalaxyl+mancozeb and mancozeb at 50, 100 and 250 ppm concentrations reduced the mycelial growth of S. rolfsii 21. The importance of fungicide mixtures in reducing the probability of resistance/tolerance development in fungi has been discussed22. Moreover, carbendazim, captan and mancozeb inhibited the mycelial growth of the fungus at a 0.1% concentration by 54.3, 67.3 and 58.7%, respectively23. Das et al.24 reported that hexaconazole 5% EC and tebuconazole 25.9 m/m EC appeared to be the best in inhibiting the hyphal growth of S. rolfsii at a 50 ppm concentration while, captan and carbendazim at 100 ppm inhibited growth by 16.0 and 3.1%, respectively. This study showed that carbendazim produced the lowest inhibition (4.30%) of mycelial growth at a 500 ppm concentration. Manu et al.25 found that carbendazim wasn’t inhibitory to the soil-borne pathogen at 250 ppm similar results were obtained by Rakholiya26, carbendazim and copper oxychloride were found to be the least effective against S. rolfsii. Based on the results obtained from this study, we recommend application of etridiazole + quintozene to control S. rolfsii is recommended.
CONCLUSION
Bulb rot disease caused by S. rolfsii has become a serious and recurrent problem in amaryllis production areas of Chiang Mai, Thailand. The pathogen was identified by PCR analyses using specific primers and morphological characteristics. The current research results showed that the most effective treatment against the pathogen was the combination of the systemic fungicides etridiazole +quintozene, which inhibited the fungus in vitro by 100% at the recommended and a half-dose. The non-systemic fungicides captan and mancozeb were of intermediate effectiveness. Therefore, protection of amaryllis against S. rolfsii, may be achieved by treating the bulbs with captan or mancozeb before planting in the plantation field. Etridiazole+quintozene added to soil infested by S. rolfsii may also protect amaryllis from infection.
SIGNIFICANCE STATEMENT
In this study a disease of amaryllis caused by Sclerotium rolfsii was identified. The study showed that, etridiazole+quintozene at a 270 ppm concentration (half recommended dose) was 100% inhibitory to the radial growth of S. rolfsii in vitro.
ACKNOWLEDGMENTS
This research was supported by the Amaryllis Breeding Project and was carried out with the cooperation of Kebol B.V. (the Netherlands), Chiang Mai University (Thailand) and H.M. The King’s initiative Centre for Flower and Fruit Propagation (Thailand).
REFERENCES
- Stirling, A.M. and G.R. Stirling, 2002. Root rot of Hippeastrum sp. caused by lesion nematode (Pratylenchus jordanensis). Aust. Plant Pathol., 31: 427-427.
CrossRefDirect Link - Raj, S.K., S.K. Snehi, S. Kumar and M.S. Khan, 2009. First molecular detection and identification of a Potyvirus associated with severe mosaic disease of amaryllis (Hippeastrum hybridum Hort.) in India. Aust. Plant Dis. Notes, 4: 50-53.
Direct Link - Rani, V.D. and H. Sudini, 2013. Management of soilborne diseases in crop plants: An overview. Int. J. Plant Anim. Environ. Sci., 3: 156-164.
Direct Link - Thiessen, L.D. and J.E. Woodward, 2012. Diseases of peanut caused by soilborne pathogens in the southwestern United States. ISRN Agron., Vol. 2012.
CrossRefDirect Link - Han, K.S., S.C. Lee, J.S. Lee, J.W. Soh and S. Kim, 2012. First report of sclerotium rot on Cymbidium orchids caused by Sclerotium rolfsii in Korea. Mycobiology, 40: 263-264.
CrossRefDirect Link - Kwon, J.H., D.W. Kang, S.D. Lee and J. Kim, 2014. First report of sclerotium rot caused by Sclerotium rolfsii on Yacon in South Korea. Plant Dis., 98: 1143-1143.
CrossRefDirect Link - Kwon, J.H., 2010. Stem rot of garlic (Allium sativum) caused by Sclerotium rolfsii. Mycobiology, 38: 156-158.
CrossRefDirect Link - Jeeva, M.L., A.K. Mishra, P. Vidyadharan, R.S. Misra and V. Hegde, 2010. A species-specific polymerase chain reaction assay for rapid and sensitive detection of Sclerotium rolfsii. Aust. Plant Pathol., 39: 517-523.
CrossRefDirect Link - Yaqub, F. and S. Shahzad, 2006. Effect of fungicides on in vitro growth of Sclerotium rolfsii. Pak. J. Bot., 38: 881-883.
Direct Link - Ohazurike, N.C., 1996. Effect of some fungicides on extracellular enzymes of Sclerotium rolfsii sacc. Mol. Nutr. Food Res., 40: 150-153.
CrossRefDirect Link - Punja, Z.K., R.G. Grogan and T. Unruh, 1982. Chemical control of Sclerorium roljirii on golf greens in northern California. Plant Dis., 66: 108-111.
Direct Link - Bhuiyan, M.A.H.B., M.T. Rahman and K.A. Bhuiyan, 2012. In vitro screening of fungicides and antagonists against Sclerotium rolfsii. Afr. J. Biotechnol., 11: 14822-14827.
Direct Link - Khan, I.H. and A. Javaid, 2015. Chemical control of collar rot disease of chickpea. Pak. J. Phytopathol., 27: 61-68.
Direct Link - Skylakakis, G., 1981. Effects of alternating and mixing pesticides on the buildup of fungal resistance. Phytopathology, 71: 1119-1121.
Direct Link - Bhagat, I. and B. Chakraborty, 2013. Efficacy of fungicides against sclerotial blight of tea plant. Our Nat., 11: 208-210.
CrossRefDirect Link - Das, N.C., B.K. Dutta and D.C. Ray, 2014. Potential of some fungicides on the growth and development of Sclerotium rolfsii Sacc. in vitro. Int. J. Scient. Res. Public., 4: 1-5.
Direct Link - Rakholiya, K.B., 2015. Screening of fungicides against Sclerotium rolfsii causing stem rot of groundnut. Bioscan, 10: 691-694.
Direct Link