
J.N. Buah
Department of Crop Science, School of Agriculture, University of Cape Coast, Ghana
J.W. Tachie-Menson
Department of Crop Science, School of Agriculture, University of Cape Coast, Ghana
G. Addae
Department of Biochemistry, School of Biological Sciences, University of Cape Coast, Ghana
P. Asare
Plant Genetic Resource and Research Institute, Ghana
American Journal of Food Technology
Year: 2011 | Volume: 6 | Issue: 8 | Page No.: 685-694







ABSTRACT
The suitability of sugarcane juice for the in vitro culture of bananas and plantains was investigated in this study. Two concentrations of sugar cane juice 5, 10% and 30 g L-1 laboratory sucrose were used. There was also a control medium without carbon source. Explants were prepared from field grown sword suckers. The preparation of the explants involved, sterilization with 70% ethanol, followed by 1% Sodium hypochlorite+1% tween 20. The explants were cultured at 26°C, under 16 h photoperiod, light intensity of 3000 lux and a relative humidity of 60%. Plants that were cultured on 5% sugarcane juice had higher average fresh weight values of 16.6 g per plant compared to those that were cultured on the 30 g L-1 sucrose and 10% sugarcane juice which had fresh weight of 15.6 g per plant and 11.0 g per plant, respectively. Oniba had higher fresh weight on medium supplemented with 5% sugarcane juice compared to the two other cultivars. In terms of dry weight, ‘Oniaba’ again had the highest value of 7.2 g per plant which was obtained on medium supplemented with 5% sugarcane juice. Plants that were cultured on 5% sugarcane juice were better in terms of shoot height and number of shoots per plant than those cultured on 30 g L-1 sucrose and the differences between the 5% sugarcane juice and the other treatments were statistically different. The various carbon sources and their concentrations also affected the water potential of the media. Medium supplemented with 10% sugarcane juice had the lowest water potential of -0.8 MPa compared to the control medium that had a water potential of -0.1 MPa. Medium supplemented with 5% sugarcane juice and 30 g L-1 sucrose had in a medium water potential of -0.3 MPa. Five percent sugarcane juice was found to be a better substitute for laboratory grade sucrose for the in vitro propagation of Musa sp.
PDF Abstract XML References Citation
Received: October 24, 2010;
Accepted: January 12, 2011;
Published: March 26, 2011
How to cite this article
J.N. Buah, J.W. Tachie-Menson, G. Addae and P. Asare, 2011. Sugarcane Juice as an Alternative Carbon Source for in vitro Culture of Plantains and Bananas. American Journal of Food Technology, 6: 685-694.
DOI: 10.3923/ajft.2011.685.694
URL: https://scialert.net/abstract/?doi=ajft.2011.685.694
DOI: 10.3923/ajft.2011.685.694
URL: https://scialert.net/abstract/?doi=ajft.2011.685.694
INTRODUCTION
Plant tissue culture is the aseptic growth of cells, tissues, organs and whole plants on artificial or definable media. The advantages of plant tissue culture are enormous and strategic in plant biotechnology (Okonko et al., 2006).
Tissue culture techniques have become an attractive field of biotechnological research and their roles are particularly appreciated in the areas of large scale clonal propagation and crop improvement. The success of tissue culture among other things relies on shoot regeneration and its efficiency requires a better understanding of the influence of culture conditions on shoot regeneration and development. Plant cells and tissues in vitro culture do not have autotrophic
ability and so require exogenous carbon source for their energy needs. Sucrose has been the main source of carbon for such in vitro culture of plants (Ahmad et al., 2007; Hilae and Te-Chato, 2005). The optimal concentration of sucrose in a medium should be enough to satisfy the energy needs for cell division and differentiation without having any negative osmotic effect on shoot formation (Javed and Ikram, 2008).
The in vitro culture of plants is a technique that has caught up in Ghana with crops like cassava, coconut, plantains/bananas, yam, potatoes, pineapple and many more now being multiplied in vitro as a way of producing large quantities of healthy planting materials. Although in vitro multiplication of plants started some 40 years ago (Bud, 1993) and is promising, extensive use of this technology is limited by the high cost of media components. Media chemicals account for less than 15% while the carbon sources such as laboratory grade sucrose contributes about 34% of the production cost (Demo et al., 2008).
For most of the developing countries like Ghana to benefit from the direct use of tissue cultured material, the cost of commercial micro propagation has to be drastically reduced without compromising the quality of micro propagules (Demo et al., 2008). This can be done through identifying cheaper alternatives to some of the expensive media components.
Sugarcane has been identified as a plant that has high amount of sucrose and other sugars (Walford, 1996) which could possibly be used as an alternative source of carbon. Prakash et al. (2002) and Alkhateeb (2006, 2008) have used date palm syrup as a carbon source for the in vitro culture of date palm.
Sugarcane grows abundantly in Ghana in seven out of the ten regions of the country. The canes are consumed raw for its juice and the bulk is used in the preparation of local gin. The price of the raw sugarcane is very cheap compared to ordinary sugar, therefore if sugarcane juice can be used as an alternative carbon source for the in vitro culture of plants, it will reduce the cost of commercial tissue culture. This study therefore seeks to investigate the possibility of using sugarcane juice as an alternative carbon source for the in vitro culture of plantains and bananas.
MATERIALS AND METHODS
Plant materials and explant preparation: The study was conducted at the tissue culture laboratory of the Plant Genetic Resource and Research Institute Bunso, Ghana from November 2009 to June 2010. Two plantain cultivars Oniaba, Apantu (Musa paradisiaca) and one Dwarf Cavendish banana cultivar (Musa accuminata) were used as the plant materials.
Plant materials were taken from sword suckers of the three cultivars of Musa sp. that had been grown in the open field at Bunso in the Eastern Region of Ghana, under good watering regime. The materials were taken early in the morning by using earth chisel to separate the sucker from the parent at the point of attachment. Before washing with running tap water, the roots and the top of the shoots were trimmed off.
The sheaths that form the pseudostem were carefully removed to reduce the size of the material to about 4 leaf sheaths. They were then sterilized with 70% ethanol for 3 min and washed three times in sterilized distilled water (Buah et al., 2000).
More leaf sheaths were then removed aseptically in a clean bench until about two leaves covered the shoot meristem. This process was followed by sterilization with 1% Sodium hypochlorite solution containing a drop of polyoxyethylenesorbitanmonolaurate (Tween 20) for five minutes with occasional shaking and there after washed three times with sterilized distilled water. Prior to their inoculation on the medium, each shoot tip (about 1cm) was longitudinally divided into two halves and again sterilized with 1% Sodium hypochlorite (NaClo) as above for 1 min.
Preparation of sugarcane juice: Sugarcanes were obtained from a farmer’s field early in the morning. They were cut into pieces and washed thoroughly under running tap for several minutes. The canes were then peeled and further cut into smaller pieces and washed again. They were then placed in a previously cleaned and dried sugarcane crusher to extract the juice. The phenol-sulphuric acid assay spectrophotometer was used to check the sucrose content of the juice (Masuko et al., 2005).
Media composition: MS medium (Murashige and Skoog, 1962) supplemented with 4.5 mg L-1 6-Benzylaminopurine was used. Three different media were prepared according to their carbon source and each treatment was replicated 10 times for each of the three Musa sp. used. The culture bottles were arranged in a completely randomized fashion. As 10% sugarcane juice, 5% sugarcane juice, 30 g L-1 sucrose and No carbon source added (control).
Thirty grams per liter sucrose was used because it had been the optimal sucrose concentration from previous work with Musa species (Buah et al., 2000).
pH of the media used was adjusted at 5.8 and 0.9 g L-1 gerlite was then added to the media before autoclaving for 15 min at 121°C.
The explants were inoculated into the media and kept under a temperature of 26°C, 16 h photoperiod with an intensity of 3000 Lux and a relative humidity of 60%. The initial sub culturing was done 4 weeks after placing the explants in the media and subsequently at two weeks interval. During subculturing, materials with multiple shoots were separately removed and placed into different vessels. In all, seven subcultures were done during which data were taken.
The isopiestic psychrometer (Tang et al., 2002) was used in measuring the water potential of the various media before inoculating them with the explants. Prior to the measurements, the jointed parts of the psychrometer were made airtight by applying vaseline to prevent the escape of water vapor from the samples. The thermocouples were then cleaned with 90% ethanol and rinsed in sterilized distilled water before drying with an air stream. About 0.4 g of each of the media samples were placed one at a time in the chamber and covered with the cylinder containing the thermocouple. A drop of sucrose at a designated concentration was placed at the tip of the thermocouple before inserting it into the cylinder that covers the chamber.
Data were taken from the first subculture on Number of shoots, Fresh weight (g), Shoot height (cm), Shoot dry weight (g) and Media water potential (Mpa).
Data was analyzed with Genstat version 7.1 for analysis of variance and Excel 2007 for the plotting of graphs (Hilbe, 2007).
RESULTS
The various sources of carbon used in this experiment affected some physical properties (water potential) of the medium as well as the growth of the explants. Sugarcane juice was found to be a good alternative to the laboratory sucrose and was evidenced in almost all the parameters measured.
The explants were taken through seven cycles of sub culturing from the 4th to the 16th week after inoculation. Generally, explants of the three Musa sp. cultured on medium supplemented with 5% sugarcane juice had higher fresh weight values than all the other treatments (Fig. 1a-c).
On the average, all the three types cultivars of Musa sp. obtained a fresh weight of 16 g per plant at the end of sixteen weeks, on medium supplemented with 5% sugarcane juice. This was followed by plants cultured on medium supplemented with 30 g L-1 sucrose which had fresh weight of 15 g per plant.
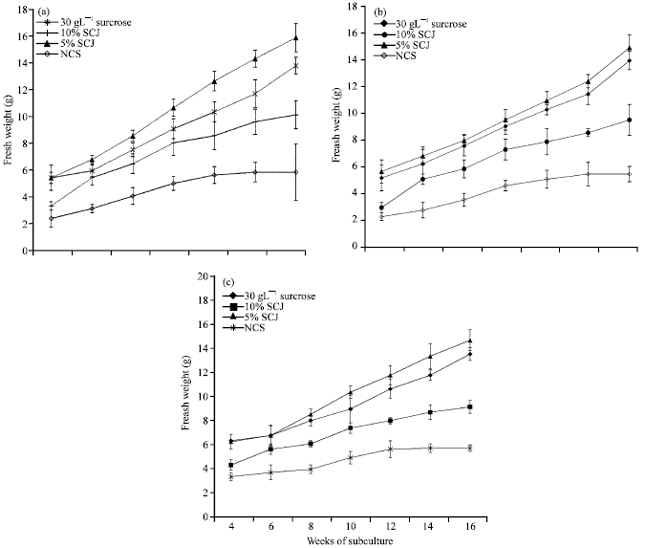 |
| Fig. 1: | Fresh weight g/plant of Musa sp. (a) Apantu, (b) Oniaba and (c) Dwarf cavendish cultured for 16 weeks on media supplemented with different carbon sources. SCJ: Sugarcane juice NCS: No carbon source |
These were better than those cultured on the 10% sugarcane juice which had average fresh weight of 8 g/plant. Plants that were cultured on the medium without a carbon source had the least fresh weight values of between 6.1 and 6.4 g per plant for all the Musa sp.
Oniaba had a slightly higher fresh weight 15.6 and 16.6 g on the 30 g L-1 sucrose and the 5% sugarcane juice respectively (Fig. 1b) compared with Apantu and Dwarf cavendish (Fig. 1a, c).
Even though the shoot height of plants cultured on media supplemented with the different carbon sources increased throughout the experimental period, there were clear differences among the various types of carbon treatments (Fig. 2a-c).
Similar to the pattern found with the fresh weight, plants cultured on 5% sugarcane juice and 30 g L-1 sucrose grew taller than those cultured on 10% sugarcane juice and the control medium. Plants cultured on 5% sugarcane juice had higher values of 17.9, 16.9 and 14.6 cm for Apantu, Oniaba and Dwarf Cavendish, respectively (Fig. 2a-c) compared with 17, 16.0 and 14.2 cm for
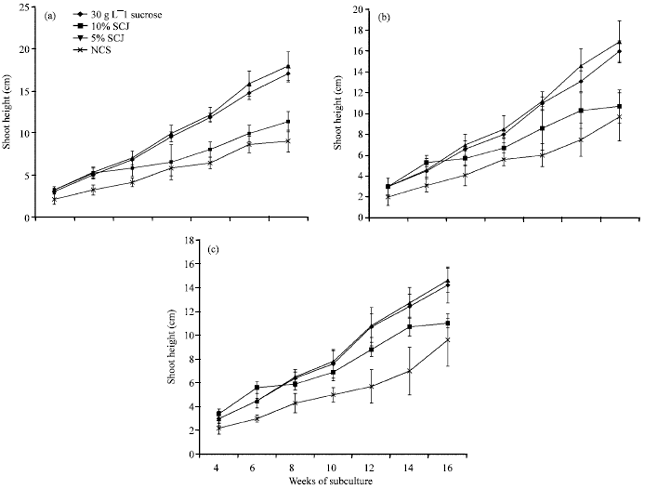 |
| Fig. 2: | Shoot height (cm) of Musa sp. (a) Apantu, (b) Oniaba and (c) Dwarf cavendish cultured for 16 weeks on media with different carbon sources. SCJ: Sugarcane juice NCS: No carbon source |
Apantu, Oniaba and Dwarf Cavendish that were cultured on 30 g L-1 sucrose. Among the three Musa sp., Apantu and Oniaba grew slightly taller than the dwarf Cavendish on media supplemented with 5% sugarcane juice and 30 g L-1 sucrose.
Media supplemented with 10% sugarcane juice and the control gave the least shoot height among the various sugar concentrations. However, plants on 10% sugarcane juice had higher values of about 11 cm compared to plants on the control medium which had values of about 9.5 cm. It was observed that plants cultured on 10% sugarcane juice grew taller initially than the other treatments and this however declined after the second sub culture.
All the three Musa sp. produced more shoots on medium supplemented with 5% sugarcane juice than those on the other media. The highest number of shoots per explant was recorded with Oniaba which produced 20.72 shoot per explant, followed by Dwarf Cavendish and Apantu with 20.10 and 15.53 shoot per explant respectively as shown in Table 1. The number of shoots produced by Oniaba (20.72) and Dwarf Cavendish (20.10) on medium supplemented with 5% sugarcane juice were statistically different from those produced by Apantu (Table 1). Also, the number of shoots produced by the three Musa sp. on 5% sugarcane juice were different statistically, from the other media treatments.
| Table 1: | Effect of carbon source on mean number of shoots for three cultivars of Musa sp. cultured for 16 weeks |
 | |
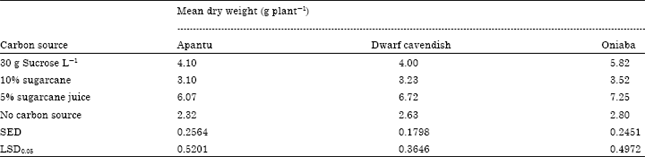
| Table 2: | Effect of carbon source on mean dry weight for three cultivars of Musa sp. cultured for 16 weeks |
 | |
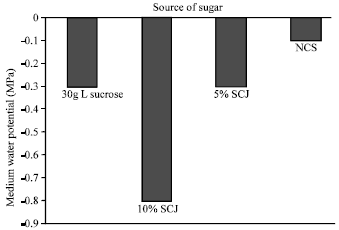 |
| Fig. 3: | Water potential of media supplemented with different carbon sources |
The dry weight of plants cultured on 5% sugarcane juice was higher than those obtained from the three other media. Oniaba had the highest dry weight of 7.25 g per plant compared with 6.72 per plant and 6.07 g per plant for Cavendish and Apantu, respectively (Table 2). The dry weight values for plant cultured on 5% sugarcane juice were statistically different from the other treatments (Table 2). Five percent sugarcane juice was followed in terms of shoot dry weight by the 30 g L-1 sucrose which had dry weight values of 4.10, 4.00 and 5.82 g per plant for Apantu, Dwarf Cavendis and Oniaba, respectively and these were also statistically higher than the values obtained for 10% sugarcane juice and the control treatments.
The water potential of the culture medium is very important for the growth of plants in vitro. The various carbon sources and their concentrations in this study, affected the water potential of the media (Fig. 3). Medium supplemented with 10% sugarcane juice had the lowest water potential of -0.8 MPa. The highest water potential of -0.1 MPa was obtained from the control medium whilst the 5% sugarcane juice and 30 g L-1 sucrose all had water potential values of -0.3 MPa.
DISCUSSION
The type of sugar and their concentrations in a culture medium affect the growth of plants in vitro. The plant materials that were cultured on the different carbon sources and their concentrations exhibited differences in their growth. This observation has also been made by Sul and Korban (1998) who reported that the type of carbon source affects the in vitro growth of plants and this depends on the plant and the part of the plant that is used as the explant.
Generally, medium supplemented with 5% sugarcane juice and 30 g L-1 sucrose produced plants with higher fresh weight compared to the 10% sugarcane juice and the medium without a carbon source. The superiority of sugarcane juice as reported in our work is also corroborated by Sul and Korban (1998) who found sugarcane juice superior to laboratory grade sucrose. Sugarcane juice has other reducing sugars apart from sucrose, known to speed up cell division thus leading to an increase in the volume and weight of tissues. Moreover, the sugars in sugarcane juice are better translocated and assimilated than pure sucrose. The juice also contains other elements like iron, phosphorus, potassium and sodium compared to the scant traces in laboratory sucrose. Such additional nutrients may further boost growth of in vitro plantlets (Demo et al., 2008).
Kodym and Zapata-Anas (2001) also confirmed this by comparing table sugar, sugar beet and sugarcane juice as carbon substitutes for the in vitro culture of Grand Naine banana and found sugarcane juice and table sugar to be superior.
On the contrary however, other researchers have found laboratory grade sucrose as the best carbon source for the in vitro culture of plants (Bahmani et al., 2009; Esan et al., 2009). Similarly, Rahman et al. (2004) have also found 30 g L-1 laboratory grade sucrose as the best carbon source for the in vitro shoot growth and development. The difference between our work and theirs however, are the plant materials. Whilst Musa sp. were used in this study, Arariae lata was used in the case of Rahman et al. (2004). Their cultures were also placed under different culture conditions and this could explain the differences in our findings with sucrose and theirs. Gawad et al. (2010) and Shatnawi et al. (2006) have also reported the superiority of fructose over laboratory grade sucrose for the in vitro growth of banana plants.
It is believed that the addition of sugar to the culture medium promotes plant growth in vitro and compensate for the low or negative net photosynthetic rate as a result of poor photosynthetic ability, thus increasing the survival rates of tissue sections cultured in vitro (Demo et al., 2008; Kubota, 2002).
The differences in the suitability of the various carbon sources as reported by the various researchers is indicative of the fact that one carbon source may not be suitable for the in vitro culture of all plants but rather the type of plant and the culture conditions may play a role.
The pattern of fresh and dry weights of plants cultured on the various sources of carbon were similar since they all decreased with increasing concentration of sugarcane juice from 5% to 10%.
Similar observation have been made by Asemota et al. (2007) and Bahmani et al. (2009) who reported a decrease in fresh weight of date palm on high concentrations of sucrose and date palm syrup. The detrimental effects of high levels of sugars in culture medium on the development of in vitro plants have also been confirmed by Hilae and Te-Chato (2005) and Yildzil et al. (2007). It is possible that the accumulation of phenolic compounds in the medium had inhibitory osmotic effect which adversely affected shoot growth and development as a result of stress (Karim et al. 2007).
On the other hand however, Baksha et al. (2003) and Shatnawi et al. (2006) have obtained better shoot formation and higher fresh weights in Lilium species that were cultured on higher concentration of sucrose.
Plants which were cultured on our control medium also had low fresh and dry weight values which could be attributed to the unavailability of sufficient energy to carry out the metabolic processes in the no sugar culture environment. In other cases, shoot have failed to form on medium that has no added sugar as was observed by Yildzil et al. (2007).
The water potential of a medium is very important to the growth and development of plants in vitro since it determines the movement of water and mineral elements into the plant material and also maintains a better turgor for the plant cells. One of the factors that affects the water potential of media is the type and concentration of sugar used (Buah et al., 2000). Water potential of the medium without sugar was the highest -0.1 MPa whilst the medium with the 10% sugarcane juice had the least water potential of, -0.8 MPa. This difference could only be attributed to the concentrations and types of carbon source since they were the only variables. In previous work by Buah et al. (2000), medium supplemented with 30 g L-1 sucrose had a water potential of -.03 MPa and the value obtained in this work confirms this. It is possible that the differences in the growth of the Musa sp. on the various media could be due to the effect these sugar sources and concentrations had on the media water potentials.
Daozhi et al. (2005) and Tang et al. (2002) have reported a decrease in leaf conductance in various plants due to decreased leaf water potential which consequently led to a decrease in the growth of the plant in vitro. Similarly, Tang et al. (2002) have reported a decrease in the fresh and dry weight of rose when leaf water potential decreased from -0.5 to -0.8 MPa. This was attributed to decreased photosynthetic activity which may be the result of decreased leaf conductance. This phenomenon may explain the better growth of plants cultured on 5% sugarcane juice and 30 g L-1 sucrose all of which had a medium water potential of -0.3 MPa.
Even though cost analysis was not done, it is believed that the replacement of laboratory grade sucrose with sugarcane juice could have a positive effect in reducing the cost of in vitro rapid multiplication of plantains and bananas. Preeti et al. (2009) successfully reduced the cost of banana tissue culture by 90% by replacing the tissue culture grade sucrose with table sugar.
CONCLUSION
It has been demonstrated that sugarcane juice as a carbon source for the in vitro culture of plantains and bananas, is comparable or better than laboratory grade sucrose in terms of the growth of the plantlets in vitro. Plants which were cultured on 5% sugarcane juice gave higher fresh and dry weights as well as plant heights compared to those cultured on 30 g L-1 laboratory grade sucrose. Therefore the use of sugarcane juice in place of laboratory grade sucrose could reduce the cost of in vitro multiplication of Musa sp. without compromising on the quality of growth. Comparing the two concentrations of sugarcane juice used (5 and 10%), the 5% was optimal for the growth of Musa sp. in vitro.
ACKNOWLEDGMENTS
Many thanks to the Biochemistry laboratory of the University of Cape Coast for making their laboratory instruments available for the analysis of sugarcane juice used in this work. Also the use of the laboratory of the Plant Genetic Resources and Research Institute of Ghana is much appreciated. Finally, we thank Dr. (Mrs) Van der Puije who took pains to edit this study.
REFERENCES
- Ahmad, T., N.A. Abbasi, I.A. Hafiz and A. Ali, 2007. Comparison of sucrose and sorbitol as main carbon energy sources in micropropagation of peach rootstock GF-677. Pak. J. Bot., 39: 1269-1275.
Direct Link - Alkhateeb, A., 2008. Comparison effect of sucrose and date palm syrup on somatic embryogenesis of date palm (Phoenix dactylifera L.). Am. J. Biotechnol. Biochem., 4: 19-23.
Direct Link - Al-Khateeb, A.A., 2006. Somatic embryogenesis in date palm (Phoenix dactylifera L.) cv. sukary in response to sucrose and polyethylene glycol. Biotechnology, 5: 466-470.
CrossRefDirect Link - Asemota, O., C.R. Eke and J.O. Odewale, 2007. Date palm (Phoenix dactylifera L.) in vitro morphogenesis in response to growth regulators, sucrose and nitrogen. Afr. J. Biotechnol., 6: 2353-2357.
Direct Link - Bahmani, R., O. Karami and M. Gholami, 2009. Influence of carbon sources and their concentrations on rooting and hyperhydricity of apple rootstock MM.106. World Applied Sci. J., 6: 1513-1517.
Direct Link - Buah, J.N., Y. Kawamitsu, S. Yonemori, M. Hayashi and S. Murayama, 2000. Effects of various carbon sources and their combinations on in vitro growth and photosynthesis banana [Musa spp.] plantlets. Plant Prod. Sci., 3: 392-397.
Direct Link - Baksha, R., R. Alam, M.Z. Karim, S.K. Mannan and B.P. Podder, 2003. Effects of auxin, sucrose and pH level on in vitro rooting of callus induced micro shoots of sugarcane (Saccharum officinarum). J. Biol. Sci., 3: 915-920.
Direct Link - Daozhi, G.I., S. Kang and J. Zhang, 2005. Responses of canopy transpiration and canopy conductance of peach (Prunus persica) trees to alternate partial root zone drip irrigation. Hydrol. Process., 19: 2575-2590.
CrossRefDirect Link - Demo, P., P. Kuria, A.B. Nyende and E.M. Kahangi, 2008. Table sugar as an alternative low cost medium component for in vitro micro-propagation of potato (Solanum tuberosum L.). Afr. J. Biotechnol., 7: 2578-2584.
Direct Link - Esan, E.B., A.A. Muyiwa and O.J. Lawal, 2009. Performance of tea (Camellia sinensis L.). on culture media modified with locally sourced substitutes. J. Anim. Plant Sci., 4: 299-303.
Direct Link - Abd El Gawad, N.M.A., N.S. Zaied and M.A. Saleh, 2010. A comparative study on different carbon source concentrations and gelling a gent on in vitro proliferation of pineapple (Ananas colossus). Nat. Sci., 8: 127-130.
Direct Link - Hilae, A. and S. Te-chato, 2005. Effects of carbon sources and strength of MS medium on germination of somatic embryos of oil palm (Elaeis guineesis Jacq). Songklanakarin J. Sci. Technol., 27: 629-635.
Direct Link - Sul, I.W. and S.S. Korban, 1998. Effects of media, carbon sources and cytokinins on shoot organogenesis in the christmas tree, scot pine (Pinus sylvestris). J. Hort. Sci. Biotech., 73: 822-827.
Direct Link - Javed, F. and S. Ikram, 2008. Effect of sucrose induced osmotic stress on callus growth and biochemical aspects of two wheat genotypes. Pak. J. Bot., 40: 1487-1495.
Direct Link - Karim, M.Z., S. Yokota, M.M. Rahman, J. Eizawa and Y. Saito et al., 2007. Effects of sucrose concentration and pH level on shoot regeneration from callus in Araria elata Seem. Asian J. Plant Sci., 6: 715-717.
CrossRefDirect Link - Kodym, A. and F.J. Zapata-Arias, 2001. Low cost alternatives for the micropropagation of banana. Plant Cell Tissue Organ Cult., 66: 67-71.
CrossRefDirect Link - Kubota, C., 2002. Photoautotrophic micro propagation: Importance of controlled environment in plant tissue culture. Combined Proc. Int. Plant Propagators Soc., 52: 609-613.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Okonko, I.O., O.P. Olabode, O.S. Okeleyi, 2006. The role of biotechnology in the socio-economic advancement and national development. An overview. Afr. J. Biotechnol., 5: 2354-2366.
Direct Link - Rahman, M.M., M.N. Amin and R. Ahmed, 2004. In vitro rapid regeneration from cotyledon explant of native olive (Elaeocarpus robustus Roxb). Asian J. Plant Sci., 3: 31-35.
CrossRefDirect Link - Tang, A.C., Y. Kawamitsu, M. Kanechi and J.S. Boyer, 2002. Photosynthetic oxygen evolution at low water potential in leaf discs lacking an epidermis. Ann. Bot., 89: 861-870.
CrossRefDirect Link - Yildizi, M., S. Onde and M. Ozgen, 2007. Sucrose effects on phenolic concentration and plant regeneration from sugarbeet leaf and petiole explants. J. Sugar Beet Res., 44: 1-15.
Direct Link - Preeti, J., R. Trivedi and S.D. Purohit, 2009. Micropropagation of Wrightia tomentosa: Effect of gelling agents, carbon source and vessel type. Indian J. Biotechnol., 8: 115-120.
Direct Link - Masuko, T., A. Minami, N. Iwasaki, T. Majima, S.I. Nishimura and Y.C. Lee, 2005. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Anal. Biochem., 339: 69-72.
CrossRefPubMedDirect Link