
Hany M. El-Naggar
Department of Floriculture, Faculty of Agriculture, Alexandria University, Egypt
Amira R. Osman
Department of Horticulture, Faculty of Agriculture, Damanhour University, Egypt
Asian Journal of Crop Science
Year: 2014 | Volume: 6 | Issue: 1 | Page No.: 58-66







ABSTRACT
Peperomia obtusifolia is a bushy upright species belong to the Piperaceae, native to Florida, Mexico and the Caribbean, as an alternative to the conventional vegetative propagation methods is becoming an important way to increase shoot proliferation rates and produce of new varieties through transformation; therefore, the purpose of this study was to establish a protocol for in vitro production of Peperomia obtusifolia and to compare between two explants for shoot proliferation and organogenesis. Single node stem segments and leaf segments of Peperomia obtusifolia were cultured on solidified Murashige and Skoog (MS) basal medium supplemented with kinetin (Kin) at 0, 2.5, 5 and 10 mg L-1 and naphthalene acetic acid (NAA) at 0, 1, 2.5 and 5 mg L-1. All explants generated from leaf segments turned dark brown and died no shoots or callus were formed. Percentage of successful shoot regenerated from single node stem segments increased by increasing kinetin concentration then declined at the highest concentration of kinetin (10 mg L-1) while different concentrations of NAA almost had no effect on percentage of shoot formation. Kinetin at 5 mg L-1 combined with NAA at 1 mg L-1 was the most effective concentrations in percentage of shoot proliferation, number of shoots, fresh weight, total leaf area, total chlorophyll and callus weight on the other hand this concentration caused a reduction in shoot length and a delay in days to proliferation.
PDF Abstract XML References Citation
Received: September 21, 2013;
Accepted: November 13, 2013;
Published: March 08, 2014
How to cite this article
Hany M. El-Naggar and Amira R. Osman, 2014. Micro Propagation and Organogenesis of Peperomia obtusifolia. Asian Journal of Crop Science, 6: 58-66.
DOI: 10.3923/ajcs.2014.58.66
URL: https://scialert.net/abstract/?doi=ajcs.2014.58.66
DOI: 10.3923/ajcs.2014.58.66
URL: https://scialert.net/abstract/?doi=ajcs.2014.58.66
INTRODUCTION
The genus Peperomia is one of the largest genera of basal angiosperms, comprising about 1500-1700 pantropically distributed species (Wanke et al., 2006). Peperomia obtusifolia is a bushy upright species belong to the Piperaceae, native to Florida, Mexico and the Caribbean (Holthe et al., 1987). It is an evergreen perennial with a thick erect stems growing to 25 cm tall and broad, cupped leathery waxy leaves and narrow spikes of greenish-white flowers on spikes up to 12 cm long, used as indoor foliage low-maintenance houseplant for indoor and bright indirect light locations (Hui and Zee, 1981).
Kinetin is a synthetic plant growth regulator with cytokinin-like physiological activity and belongs to a group of plant growth regulators known as cytokinins. N6-furfuryladenine (kinetin) belongs to the family of N6-substituted adenine derivatives known as cytokinins. Cytokinins are plant hormones that promote cell growth, development and division (Zhang et al., 1996).
Many of the cytokinins are structurally based on adenine, substituted at the N6-atom, either by an isoprenoid side-chain or an aromatic ring. Cytokinins participate in a number of developmental processes such as cell division and differentiation, end the dormant state of lateral buds, chloroplast development, chlorophyll biosynthesis, cellular senescence delay, differentiation of vascular tissue, as well as having roles in the development of flowers and fruits (Mok and Mok, 2001).
Cytokinins such as kinetin act as cell division mediating factors and upon the differentiation and arrangement of parenchyma cells and procambium necessary for shoot development and regulation, plant meristem activity as the apical meristem decreased in size with reduction of cytokinin levels and are generally known to promote formation of buds and shoot differentiation in tissue cultured organs (Nitsch et al., 1967; Werner et al., 2001; Ishimori et al., 2007).
Cytokinins play an important role in the promotion and maintenance of plant cell division in cell and tissue cultures (Ahmadabadi and Bock, 2010).
Natural and synthetic auxins have been used extensively in plant cell, tissue and organ cultures to elicit specific morphogenetic responses (Nissen and Sutter, 1990)
The use of auxins, naphthaleneacetic acid (NAA), has increased because they often appear to be more effective than IAA for induction of morphogenetic responses. This enhanced effectiveness of auxins other than IAA is presumed to be due to their increased stability (Nissen and Sutter, 1990).
This study describes the procedures of in vitro propagation of Peperomia obtusifolia using single node stem segments and leaf segments.
MATERIALS AND METHODS
This study was done in March 2013 at the Tissue Culture and Biotechnology Laboratory of the Horticulture Department, Faculty of Agriculture, Alexandria University, Egypt.
Plant material: Single node stem segments and leaves (fully unfolded leaves from the 4th and 5th node from the growing point) of Peperomia obtusifolia were brought from commercial nursery. All plants were washed in the beginning with home detergent and rinsed 3 times, leaves and shoot cuttings were dipped after that in ethyl alcohol 70% for 30 sec, washed with distilled water and then dipped in Clorox bleach 20% for 10 min then washed 3 times with autoclaved distilled water.
Shooting medium: Single node stem segments and leaf segments (1 cm2 from middle leaf section containing the main midrib) were cultured on solidified media containing 4.43 g L-1 MS with vitamins (Murashige and Skoog, 1962) supplemented with 30 g L-1 sucrose and 8 g L-1 agar, the pH was adjusted at 5.8±0.1 and poured in tubes (2.5 cm diameterx15 cm height).
For shoot proliferation, four different concentrations of kinetin (0, 2.5, 5 and 10 mg L-1) and four different concentrations of NAA (0, 1, 2.5 and 5 mg L-1) were added to the basal medium.
Rooting medium: Rooting was achieved by transferring the regenerated shoot after 3 months (5-7 cm height) to rooting medium containing 4.43 g L-1 MS with vitamins+30 g L-1 sucrose+5 g L-1 agar+0.5 g L-1 NAA and pH was adjusted to 5.8±0.1. 25 mL of medium was poured in tubes (2.5 cm diameterx15 cm height).
All media were autoclaved at 121°C and 15 bar cm-2 for 20 min. Plants were cultured in tubes and placed under cool white florescent light at intensity of 66 to 52 μmol m-2 sec-1 for 16 and 8 h in the dark at 25±1°C (Fig. 1).
 | |
| Fig. 1(a-b): | Direct organogenesis of single node stem segments of (a) and (b) Peperomia obtusifolia |
Hardening: Complete plantlets were taken out from test tubes and medium was removed using tissue paper to absorb medium remains from roots. In vitro cultured plantlets were transplanted in 6 cm pots containing a mixture of peat moss and perlite 2:1 (v/v), commercial fungicide was added to the soil mixture to protect the fragile plantlets from any fungal attack and the plantlets were kept under transparent plastic cups to keep the humidity high for acclimatization for 10 days. Irrigation was done whenever needed, the rooted plantlets were transferred after that to green house (Fig. 2).
Data collected: Data collected were percentage of shoot formation, number of shoots/explant, shoot length, the fresh weight of all aerial parts, total leaf area, total chlorophyll concentration using SPAD-502 meter (Konica-Minolta, Japan), days to shoot proliferation and callus weight.
Statistical analysis: Two experiments, depending on the type of explants, were done; one for the stem cuttings and the other for the leaf segments, each was designed as factorial design with two factors (kinetin at 0, 2.5, 5 and 10 mg L-1 l and NAA at 0, 1, 2.5 and 5 mg L-1) each with 3 replicates. Data were analyzed for significance by analysis of variance (ANOVA) using the SAS program (SAS, 2002) and Tukey’s multiple comparisons method for least significant difference at confidence level of p<0.05 (Tukey, 1994).
RESULTS
All explants generated from leaf segments turned dark brown and died no shoots or callus were formed also no shoots or callus were formed from single node stem segments when kinetin was used at 0 mg L-1.
 | |
| Fig. 2: | Indirect organogenesis of single node stem segments of Peperomia obtusifolia showing callus formation first then shoot regeneration |
| Table 1: | Effect of kinetin on in vitro culture of shoot cuttings of Peperomia obtusifolia |
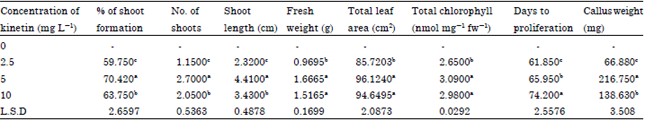 | |
| Same letters in the same column means there are no significant differences between treatments at p<0.05 | |
| Table 2: | Effect of NAA on in vitro culture of shoot cuttings of Peperomia obtusifolia |
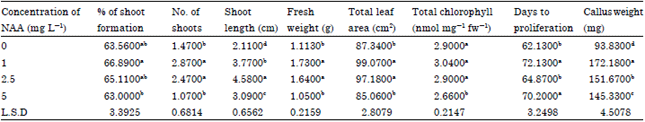 | |
| Same letters in the same column means there are no significant differences between treatments at p<0.05 | |
From the results presented in this study it appeared that the percentage of successful shoot regenerated from single node stem segments increased by increasing kinetin concentration then it was declined at the highest concentration of kinetin (10 mg L-1) while different concentrations of NAA almost had no effect on percentage of shoot formation (Table 1, 2) and the best percentage of shoot regeneration was (71%) at 5 mg L-1 kinetin and NAA at 2.5 mg L-1.
 | |
| Fig. 3: | Root formation on Peperomia obtusifolia after transferring of formed shoots to rooting medium |
| Table 3: | Effect of interaction between kinetin and NAA on in vitro culture of shoot cuttings of Peperomia obtusifolia |
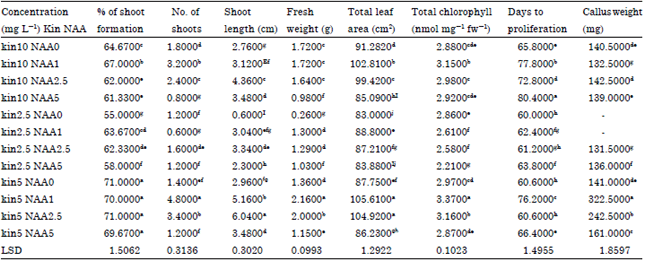 | |
| Same letters in the same column means there are no significant differences between treatments at p<0.05 | |
It was found that the no callus was produced at 0 mg L-1 kinetin under all NAA concentrations also no callus was formed and shoots was formed directly without callus formation (direct organogenesis) at 2.5 mg L-1 kinetin combined with 0 and 1 mg L-1 NAA (Fig. 3). While callus was formed before shoot proliferation at different kinetin concentrations (Fig. 4).
Also it was found that the fresh weight and total leaf area per plant increased with kinetin at 5 and 10 mg L-1 and NAA at 1 or 2.5 mg L-1 as shown in Table 1 and 2 while the best combination was 5 mg L-1 kinetin with 1 mg L-1 NAA (Table 3).
 | |
| Fig. 4(a-b): | In vitro cultured plantlets transplanted in 6 cm pots containing a mixture of peat moss and perlite 2:1 (v/v), (a) Plantlets during acclimatization covered with transparent plastic cups and (b) Plantlets after hardening transferred to green house |
Total chlorophyll content increased by increasing kinetin concentration at (5 and 10 mg L-1) while NAA almost had no effect on chlorophyll concentration since there were no significant differences between 0, 1 and 2.5 mg L-1 (Table 1, 2) and the best chlorophyll content was at 5 mg L-1 kinetin with 1 mg L-1 NAA (Table 3).
Number of shoots and shoot length increased significantly at 5 mg L-1 kinetin and 2.5 mg L-1 NAA (Table 1, 2), the best combination for number of shoots formed was kinetin and NAA at 5 and 1 mg L-1 respectively while for shoot length the best combination was 5 mg L-1 kinetin with 2.5 mg L-1 NAA (Table 3).
Number of days needed for shoot regeneration was delayed by increasing kinetin concentration and kinetin at the highest concentration cased a significant delay in days to proliferation (74.2 days) also (Table 1) combining kinetin and NAA at their highest concentrations (10 and 5 mg L-1 respectively) caused a significant delay in days to proliferation (80.4 days) (Table 3).
DISCUSSION
Kinetin at 5 mg L-1 was the most effective concentration for percentage of shoot proliferation from shoot cuttings reaching 70.42% (Table 1) or 71% when combined with NAA at 2.5 mg L-1 (Table 3) then declined at higher kinetin concentration (10 mg L-1) and this is in agreement with Robert and Louis (2012) who mentioned that percentage of shoot response decreased with further increase in the concentration of kinetin also the study of Han et al. (2004), Bacchetta et al. (2003) and Mik et al. (2011) who mentioned that cytokinins in general at high concentrations inhibited shoot formation.
Further increases in kinetin concentration to 8.0 mg L-1, resulted in a decreased proliferation rate of Musa spp., indicating an adverse effect of plant growth regulators beyond the optimal dose (Muhammad et al., 2007).
Number of shoots formed increased significantly by increasing kinetin then the number of shoots was reduced at the highest kinetin concentration (10 mg L-1) also NAA at its highest concentration caused a reduction in shoots number same results were obtained by Robert and Louis (2012), best number of shoot formation occurred at 5 mg L-1 kinetin combined with NAA at 1 mg L-1 (Table 3). Mungole et al. (2009) mentioned that significant number of shoot formation and length can be achieved in the combination of NAA and kinetin.
Shoot length increased by increasing kinetin concentration then declined at 10 mg L-1 kinetin (Table 1) also shoot length increased by increasing NAA reaching its peak at 2.5 mg L-1 (4.58 cm) (Table 2) and then declined at higher concentration and the highest shoot length was achieved at 5 mg L-1 kinetin combined with 2.5 mg L-1 NAA (6.04 cm) (Table 3). Same results were reported by Robert and Louis (2012) who found that Dipteracanthus prostrates shoot length increased significantly by increasing kinetin and NAA concentrations then declined by increasing both concentrations.
These results are in agreement with previous findings where levels of kinetin promoted shoot multiplication but prevent shoot elongation at high concentrations. Also with the findings by Campanoni and Nick (2005) who mentioned that NAA enhanced cell elongation by almost 50% as compared to the control and that NAA triggers cell elongation.
Cytokinin in in vitro growth culture played a key role in de-etiolation of the hypocotyls length of Arabidopsis plant were increasingly shorter with increasing concentration of kinetin (Chory et al., 1994).
Fresh weight increased by increasing kinetin concentration (Table 1) same observations were stated by Tounekti et al. (2011) who found that applying kinetin improve considerably the total leaf fresh weight of Salvia officinalis, NAA also increased fresh weight at low concentration 1 mg L-1 then the weight declined at higher concentrations (Table 2) the results are in agreement with Negahdar et al. (2012) and this might be due to the ability of kinetin to increase leaf water content also the effect of plant growth regulator to enhance cell division and elongation and increase plant biomass.
Total chlorophyll content increased significantly by increasing kinetin application (Table 1) the kinetin alleviated chlorophyll content and enhanced photosynthesis and gas exchange (Tounekti et al., 2011) and the role of cytokinins as retardants of chlorophyll and protein degradation during senescence of leaves was documented by Anderson and Rowan (1968), the primary effect of kinetin upon leaf senescence is the maintenance of RNA synthesis and hence protein synthesis (Von Abrams and Pratt, 1966).
Cytokinins participate in chloroplast development, chlorophyll biosynthesis and cellular senescence delay (Mok and Mok, 2001).
Kinetin and kinetin derivatives also protected membrane lipids against the negative action of the reactive oxygen species that accumulate in tissues during leaf senescence causing delaying in chlorophyll degradation (Mik et al., 2011).
Kinetin used at lower concentrations formed fast division tobacco callus cells, when the concentration was higher, compact callus with differentiation of conducting elements was formed (Angelova et al., 2001). Also, Mik et al. (2011) mentioned that the callus fresh weight of tobacco plants increased by increasing kinetin and then declined at high concentrations. Same results were obtained in this study and the highest callus fresh weight was obtained at 5 mg L-1 kinetin (216.75 mg) Table 1 and (322.5 mg) when 5 mg L-1 kinetin was combined with 1 mg L-1 NAA (Table 3).
Kinetin application enhanced total leaf area at 5 and 10 mg L-1 while NAA at its highest concentration caused a reduction in leaf area (Table 2).
Lou et al. (2012) found that kinetin application increased leaf area also increased the length and width and loquat fruit size since kinetin plays a key role in cell division, delay of senesce and cell enlargement (Rana et al., 2011).
Significant changes on leaf growth, both leaf area and biomass, were accompanied with significant changes in endogenous cytokinin levels, whereas levels of auxin did not change significantly (Juvany et al., 2012).
Pan et al. (2000) mentioned that kinetin increased grain and sweet corn leaf area then leaf area declined at higher concentration.
CONCLUSION
Single node stem segments of Peperomia obtusifolia were better than leaf segments in percentage of shoot proliferation when used as explants reaching 71%. Kinetin at 5 mg L-1 combined with NAA at 1 mg L-1 was the most effective concentrations in percentage of shoot proliferation, number of shoots, fresh weight, total leaf area, total chlorophyll and callus weight on the other hand this concentration caused a reduction in shoot length and a delay in days to proliferation.
REFERENCES
- Ahmadabadi, M. and R. Bock, 2010. Development of a highly responsive leaf-based regeneration system for Peperomia species. Turk. J. Bot., 33: 329-334.
CrossRefDirect Link - Angelova, Y., S. Petkova, E. Zozikova, E. Kotseva and L. Iliev, 2001. Effects of kinetin and 4PU-30 on the growth and the content of polyphenols in tobacco callus tissue. Bulgarian J. Plant Physiol., 27: 36-42.
Direct Link - Campanoni, P. and P. Nick, 2005. Auxin-dependent cell division and cell elongation. 1-Naphthaleneacetic acid and 2,4-dichlorophenoxyacetic acid activate different pathways. Plant Physiol., 137: 939-948.
PubMed - Chory, J., D. Reinecke, S. Sim, T. Washburn and M. Brenner, 1994. A role for cytokinins in de-etiolation in Arabidopsis (det mutants have an altered response to cytokinins). Plant Physiol., 104: 339-347.
Direct Link - Han, B.H., H.J. Yu, B.W. Yae and K.Y. Peak, 2004. In vitro micropropagation of Lilium longiflorum Georgia by shoot formation as influenced by addition of liquid medium. Sci. Hort., 103: 39-49.
CrossRef - Holthe, P.A., L.D.S. Sternberg and I.P. Ting, 1987. Developmental control of CAM in Peperomia scandens. Plant Physiol., 84: 743-747.
Direct Link - Hui, L.H. and S.Y. Zee, 1981. In vitro propagation of Peperomia viridis using medium supplemented with ginseng powder. HortScience, 16: 86-87.
Direct Link - Robert, J. and C. Louis, 2012. An efficient in vitro plant regeneration of Dipteracanthus prostratus (Poir.) Nees.-a medicinal herb. Asian Pacific J. Trop. Biomed., 2: S484-S487.
CrossRef - Juvany, M., M. Muller and S. Munne-Bosch, 2012. Plant age-related changes in cytokinins, leaf growth and pigment accumulation in juvenile mastic trees. Environ. Exp. Bot., 87: 10-18.
CrossRef - Bacchetta, L., P.C. Remotti, C. Bernardini and F. Saccardo, 2003. Adventitious shoot regeneration from leaf explants and stem nodes of Lilium. Plant Cell Tissue Organ Cult., 74: 37-44.
CrossRef - Lou, H., P. Chen, H. Zheng, C. Xu and H. Lu, 2012. Effect of kinetin on quality and harvest date of loquat fruit. Afr. J. Agric. Res., 7: 1577-1583.
Direct Link - Mik, V., L. Szucovs, M. Smehilovs, M. Zatloukal and K. Dolezal et al., 2011. N9-substituted derivatives of kinetin: Effective anti-senescence agents. Phytochemistry, 72: 821-831.
CrossRef - Mok, D.W.S. and M.C. Mok, 2001. Cytokinin metabolism and action. Ann. Rev. Plant Physiol. Plant Mol. Biol., 52: 89-118.
CrossRefDirect Link - Muhammad, A., H. Rashid, I. Hussain and S.M.S. Naqvi, 2007. Proliferation-rate effects of BAP and kinetin on banana (Musa spp. AAA Group) Basrai. HortScience, 42: 1253-1255.
Direct Link - Mungole, A., R. Awati, S. Dey, A. Chaturvedi and P. Zanwar, 2009. In-vitro callus induction and shoot regeneration in Ipomoea obscura (L.): Potent Indian medicinal plant. Ind. J. Sci. Technol., 2: 24-26.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Negahdar, N., B. Kaviani, D. Hashemabadi and M. Zarchini, 2012. Effect of naphthalene acetic acid (NAA) on micropropagation of acacia (Robinia pseudoacacia L.). Ann. Biol. Res., 3: 4797-4800.
Direct Link - Nissen, S.J. and E.G. Sutter, 1990. Stability of IAA and IBA in nutrient medium to several tissue culture procedures. HortScience, 25: 800-802.
Direct Link - Pan, B., Y.M. Bai, S. Leibovitch and D.L. Smith, 2000. Plant growth promoting rhizobacteria and kinetin as ways to promote corn growth and yield in a short-growing season area. Eur. J. Agronom., 11: 179-186.
Direct Link - Ishimori, T., Y. Niimi and D.S. Han, 2007. Benzyladenine and low temperature promote phase transition from juvenile to vegetative adult in bulblets of Lilium x formolongi White Aga cultured in vitro. Plant Cell. Tissue Organ Culture, 88: 313-318.
CrossRefDirect Link - Tounekti, T., I. Hernandez, M. Muller, H. Khemira and S. Munne-Bosch, 2011. Kinetin applications alleviate salt stress and improve the antioxidant composition of leaf extracts in Salvia officinalis. Plant Physiol. Biochem., 49: 1165-1176.
CrossRef - Wanke, S., M.S. Samain, L. Vanderschaeve, G. Mathieu, P. Goetghebeur and C. Neinhuis, 2006. Phylogeny of the genus Peperomia (Piperaceae) inferred from the trnK/matK region (cpDNA). Plant Biol., 8: 93-102.
CrossRef - Werner, T., V. Motyka, M. Strnad and T. Schmulling, 2001. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA., 98: 10487-10492.
CrossRefDirect Link - Zhang, K., D.S. Letham and P.C. John, 1996. Cytokinin controls the cell cycle at mitosis by stimulating the tyrosine dephosphorylation and activation of p34cdc2-like H1 histone kinase. Planta, 200: 2-12.
CrossRef