
Ling-Bin Liu
College of Animal Science and Technology, Sichuan Agricultural University, Ya�an, Sichuan, 625014, People�s Republic of China
Di-Yan Li
College of Animal Science and Technology, Sichuan Agricultural University, Ya�an, Sichuan, 625014, People�s Republic of China
Xiao-Ling Zhao
College of Animal Science and Technology, Sichuan Agricultural University, Ya�an, Sichuan, 625014, People�s Republic of China
Yi-Ping Liu
College of Animal Science and Technology, Sichuan Agricultural University, Ya�an, Sichuan, 625014, People�s Republic of China
Yan Wang
College of Animal Science and Technology, Sichuan Agricultural University, Ya�an, Sichuan, 625014, People�s Republic of China
Qing Zhu
College of Animal Science and Technology, Sichuan Agricultural University, Ya�an, Sichuan, 625014, People�s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 11 | Page No.: 1183-1190







ABSTRACT
Prolactin Receptor (PRLR), as an important regulatory gene about growth and differentiation, might be a candidate gene for reproductive traits. The purpose in the present study was to analyze the association of Single Nucleotide Polymorphisms (SNPs) of exon 6 in prolactin receptor gene with egg production traits in Erlang mountainous chickens, a native breed from Poultry farm of Sichuan Agricultural University which consisted of two pure lines SD02 and SD03. Polymerase Chain Reaction-single-strand Conformation Polymorphism (PCR-SSCP) and DNA sequencing methods were used. Results showed that One SNP (G1836C) which had not yet been reported was found and two alleles (G, C) and two genotypes (GG, CC) were also detected. It observed significant differences in allelic and genotypic frequencies among all the chicken populations. Genotype frequencies in all chicken for GG and CC were 56.22 and 43.78%, respectively. Associations of polymorphisms of the PRLR gene with egg production traits were analyzed by using a GLM procedure. The CC genotype had extremely significant effect on Age at First Egg (AFE) (p<0.01) and the AFE of CC genotype had 3.75 days earlier than GG genotype. There was no significant association of G1836C with BWFE; body weight at first egg, WFE; weight at first egg, EN; egg number during 300 days after hatch, BWTA; body weight at 300 days of age and WTE; egg weight at 300 days of age. Therefore, the study showed that the PRLR gene may either a major gene that influences age at first egg of chicken or a molecular marker in close linkage with such a gene.
PDF Abstract XML References Citation
Received: May 28, 2012;
Accepted: July 19, 2012;
Published: September 15, 2012
How to cite this article
Ling-Bin Liu, Di-Yan Li, Xiao-Ling Zhao, Yi-Ping Liu, Yan Wang and Qing Zhu, 2012. Polymorphism of Prolactin Receptor Gene and its Association with Egg Production Traits in Erlang Mountainous Chicken. Asian Journal of Animal and Veterinary Advances, 7: 1183-1190.
DOI: 10.3923/ajava.2012.1183.1190
URL: https://scialert.net/abstract/?doi=ajava.2012.1183.1190
DOI: 10.3923/ajava.2012.1183.1190
URL: https://scialert.net/abstract/?doi=ajava.2012.1183.1190
INTRODUCTION
Prolactin (PRL) was a polypeptide hormone secreted by the anterior pituitary gland which had diverse biological actions in vertebrates (Alamer, 2011). The chicken prolactin can modulate maternal behaviors such as affect egg production (Chen et al., 2011). Onset of poultry broodiness was induced by an increase secretion of prolactin (Cui et al., 2006). Broodiness was associated with decreased egg production, frequent nest occupancy, reduced feed and water intake, regressive ovary, aggressiveness or defensive behaviors, characteristic clucking and raised body temperature in chickens (Jiang et al., 2005). Prolactin which bind with prolactin receptor triggered a series of physiological and biochemical responses to exercise functions. Prolactin receptor (PRLR) protein was a kind of separate membrane protein and belonged to superfamily of cytokine receptor. It consisted of membrane domain, trans-membrane domain and intracellular domain (Wells and De Vos, 1996). Literature reported that female mice which had been knockout PRL and PRLR gene showed some exceptional symptoms such as irregular estrus, reduction in ovulation rate, false pregnancy, damaged maternal behavior, infertility and loss of luteal function (Horseman et al., 1997; Kelly et al., 2001).
PRLR, as an important regulatory gene about growth and differentiation, might be a candidate gene for reproductive traits (Van Rens et al., 2003; Van Rens and van der Lende, 2002; Vincent et al., 1998) and plays an important role in the PRL signal transduction cascade. PRLR is located on 13th chromosome in mouse (Clarke and Linzer, 1993), 17th chromosome in rat (Shirota et al., 1990), 23th chromosome in cow (Scott et al., 1992), 6th chromosome in human (Boutin et al., 1989), 7th chromosome in pig (Vincent et al., 1998), 16th chromosome in sheep (Jenkins et al., 2000) and Z chromosome in chicken (Tanaka et al., 1992), respectively. In birds, sex is determined by two chromosomes, Z and W. Males are homozygous ZZ and females are hemizygous ZW (Elferink et al. 2008). Because the PRLR gene is on the Z chromosome (Dunn, 1998), the hypothesis of sex-linked inheritance of reproductive traits may relate to PRLR gene.
In farm animals, many polymorphisms of PRLR gene have been identified different organisms as pigs (Vincent et al., 1998; Drogemuller et al., 2001; Putnova et al., 2002), goose (Chen et al., 2012b), goat (Zhang et al., 2007). Compared with other animals, the exon regions of the chicken PRLR (cPRLR) gene was highly polymorphic and the studies showed that these polymorphisms were associated with broodiness, early egg productivity and egg weight average (Dunn, 1998; Jiang et al., 2005; Hong et al., 2007). The objectives of the present study were to identify the SNPs of the chicken exon 6 of PRLR gene (PRLR6) and analyze the associations between polymorphisms of the exon 6 of PRLR gene and egg production traits in chicken.
MATERIALS AND METHODS
Chicken populations and trait measurement: We selected Erlang Mountainous Chickens from Poultry farm of Sichuan Agricultural University which consisted of two pure lines SD02 (N = 200, including 100 the first generation and 100 the second generation) and SD03 (N = 186, including 100 the first generation and 86 the second generation) form May 10, 2011 to July 15, 2011. As a native breed, the Erlang Mountainous Chicken is one cultivated breed which has been successfully developed from the local chicken breeds in Sichuan province (Xiao et al., 2011). The Erlang Mountainous Chicken is a kind of dual purpose type population and has some excellent characteristics, such as large body, high slaughter rate, delicate meat, high nutrition value on egg and so on. SD02 focuses on meat performance at present; in the meantime the quality of egg is the primary breeding goal on SD03 (Jie et al., 2010). All birds were hatched on the same day, housed on the deep-litter bedding and moved to the growing pens at the age of 7 weeks. Birds had access to feed (commercial corn-soybean diets meeting NRC requirements) and water ad libitum. The data of egg production was individually collected from the age at first laying to 300 days. The studied traits included age at first egg; AFE, body weight at first egg; BWFE, egg weight at first egg; WFE, egg number by 300 days after hatch; EN, body weight at 300 days of age; BWTA, egg weight at 300 days of age; WTE. The blood samples from wing vein were finally collected according to strict requirements of animal welfare and stored at 4°C (Chen et al., 2012a). The genomic DNA was isolated by the standard phenol/chloroform method (Han et al., 2011).
The PRLR gene polymorphisms: The PCR primers were designed based on the GenBank PRLR gene sequence (No: AY547323) (Xing et al., 2011).
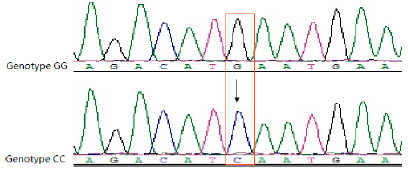 | |
| Fig. 1: | SNP in the deoxyribonucleic acid sequence of PRLR gene. One novel variant was found at 1836 bp (G→C) of the CDS of the PRLR gene |
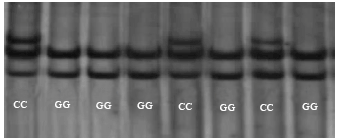 | |
| Fig. 2: | Electropherograms of the polymorphic pattern for the G1836C SNP. The two genotypes of CC and GG are marked in the figure, respectively |
| Table 1: | Genotypes and alleles frequencies of the exon 6 of PRLR single nucleotide polymorphism (SNP) in Erlang Mountainous chickens |
 | |
| PIC: Polymorphism information content, FGE: First generation of Erlang mountainous chickens, SGE: Second generation of Erlang mountainous chickens, Values in parenthesis are represented by the percentage | |
One pair of primer was employed to amplify the fragment (195 bp) of the sixth exon fully containing the coding sequence (CDS) and investigated the genetic variation by using PCR-single-strand conformation polymorphism (PCR-SSCP) method. The primer sequence was as follow:
| • | forward 5'-GCATTCAATGTACTGATTATTT-3' |
| • | reverse 5' -AATTATACAGCACCTCTTACC-3' |
The PCR was performed in a volume of 7 μL containing 0.6 μL (ng L-1) of template, 3 μL of 2xTaq PCR MasterMix (Beijing TIAN WEI Biology Technique Corporation, Beijing), 3 μL of ddH2O and 0.3 μL of each primer (10 pmol μL-1) with the following profile: 94°C for 4 min, 35 cycles at 94°C for 30 sec, 50°C for 30 sec, 72°C for 30 sec an extension at 72° for 10 min.
| Table 2: | Associations of the exon 6 of PRLR single nucleotide polymorphism (SNP) with egg production traits |
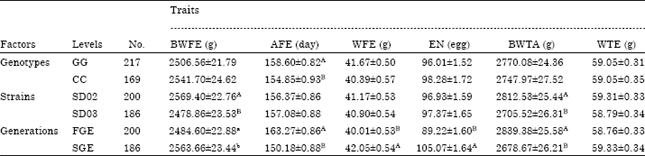 | |
| Values are presented as least squares Means±SD. The capital letters for the same line indicate that multiple comparison is significant at p<0.01 and small letters indicate that multiple comparison is greatly significant at p<0.05, BWFE: Body weight at first egg, AFE: Age at first egg, WFE: Egg weight at first egg, EN: Egg number during 300 days after hatch, BWTA: Body weight at 300 days of age, WTE: Egg weight at 300 days of age | |
The PCR products were screened by using the SSCP method and were separated by 4% polyacrylamide gel electrophoresis (Jun et al., 2010). Genotypes were recorded according to the band patterns. PCR products that had several profiles were further amplified, purified and sequenced by a commercial sequencing company (Invitrogen, Shanghai, China).
Statistical analysis: Data was analyzed with GLM (General Linear Models Procedures) procedures of SAS 8.1 (SAS Institute Inc., Cary, NC) and the genetic effects were analyzed by mixed procedure according to the following model:
Yijk = μ+Bi+Gj+Gek+eijk |
where, Yijk is trait; μ is population mean; Bi is fixed effects of line; Gi is fixed effects of genotype; Ge is fixed effects of generation and eijk is random error. The interaction BixGj, BixGek and GjxGek were not significant for any trait and therefore were not included in the model. The values were presented as least square means standard error (Qiong et al., 2011). The significance of least square means was tested with the Duncan test (p<0.05).
RESULTS
PCR amplification and SSCP detection: The PCR products (169 bp) were amplified as expected, thus SSCP analysis was applied to decide band patterns. PCR products with multiple profiles on gel were sequenced to screen. Sequencing results showed the nucleotide substitutions a G→C mutation at position 1836 bp of the CDS of PRLR gene (Fig. 1) in exon 6 of PRLR gene. The G1836C SNP didn’t lead to an amino acid variation and was a synonymous mutation. Two genotypes GG and CC were detected in the SSCP gel (Fig. 2).
Genotypes and alleles frequencies: Genotypes and alleles frequencies of G1836C mutation in Lines SD02 and SD03 were summarized (Table 1). Genotype frequencies for GG and CC were 56.22 and 43.78%, respectively. And allele frequencies for G and C were 56.22 and 43.78%, respectively. The Polymorphism Information Content (PIC) of the SNP was 0.3711.
Association between genotype and egg production: The results of the GLM analysis of association between the exon 6 of PRLR gene polymorphism, strains and generations with egg production traits were summarized in Table 2. The genotype had extremely significant effect on AFE among the six investigated egg production traits (p<0.01) and AFE of CC genotype (154.85±0.93 days) was earlier than GG genotype (158.60±0.82 days). In contrast, the six traits didn’t significantly differ between GG and CC genotypes. With the exception of extremely significant differ on BWFE and BWTA (p<0.01), others egg production traits did not significantly differ between SD02 and SD03 strains. SD02 had higher BWFE (2569.40±22.76 g) than SD03 (2478.86±23.53 g) and SD02 had higher BWTA (2812.53±25.44 g) than SD03 (2705.52±26.31 g) equally. Without significant effect on BWFE (p<0.05) and insignificant effect on WTE, others egg production traits had extremely significant different (p<0.01) between the first generation (FGE) and the second generation (SGE).
DISCUSSION
The G1836C mutation may impact the exertion of PRL physiological effects, finally resulting in variation of production performance. Compared with the NCBI SNP database, the G1836C mutation is a novel one in chicken. A large number of studies have been carried out on PRLR polymorphisms in recent years. Drogemuller et al. (2001) amplified different strains of German swine and discovered that PRLR gene displayed polymorphism. Five pairs of primers were designed to detect single nucleotide polymorphisms of exon 10 and part of 3' Untranslated Region (UTR) of PRLR gene in goat by PCR-SSCP and 3 primers displayed polymorphisms (Zhang et al., 2007). In the present experiment, two genotypes in one SNP were detected for the first time. Heterozygous genotype (GC) was included impossibly in the population because of PRLR locating on Z chromosome in chicken.
PRLR, as a candidate gene of reproduction, has been researched in some fields. Cao et al. (2004) detected a SNP locus (Alu) in swine PRLR6 gene by using PCR-RFLP and the results implied that there were no significant difference between different genotype sows in the total number born and number born alive but the trend of AA>AB>BB existed. Putnova et al. (2002) had researched association between the alleles and reproduction. Vincent et al. (1998) suggested that exon 10 of PRLR had significant effect on litter size and livestock production in pig. Jiang et al. (2005) discovered three SNPs in PRLR gene of GreenShell and also demonstrated the SNPs had no significant effect on early egg productivity. However, Dunn (1998) discovered that PRLR gene may be a molecular marker related to chicken broodiness QTL and Hong et al. (2007) discovered that the P value of the salience test of the egg weight average at 9-11 weeks after the age at 50% egg was 0.08 in PRLR exon 3. Three genotypes were found in researches with P4 primers of single nucleotide polymorphisms of PRLR10. The results suggested for the first time that PRLR was a promising candidate gene that can affect egg performance in Wanjiang white goose (Chen et al., 2012b). However, the relationship between chicken PRLR6 gene polymorphism and egg production traits have been reported seldom. In the present study, we discovered that the genotype had extremely significant effect (p<0.01) on AFE (age at first egg) in PRLR6 polymorphism association with egg production traits and AFE of CC genotype was earlier than GG genotype. Although the sample quantity was not massive, PIC analysis (PIC = 0.3711) indicated that PRLR polymorphism was abundant. These results preliminarily showed that the PRLR gene may be either a major gene that influences age at first egg of chicken or a molecular marker in close linkage with such a gene. Jiang et al. (2005) discovered 2 SNP in the exon 6 of PRLR in chicken, the SNP of G14820A didn’t contribute to broodiness because it was synonymous mutation; the T14771C, a missense mutation, leads to Leu340Ser. And this amino acid variation occurred in the cytoplasmic tail of the PRLR and may not influence the functional structure. In our study, the G1836C mutation was a synonymous mutation. But, Kimchi-Sarfaty et al. (2006) suggested that synonymous mutation can affect the timing of cotranslational folding and insertion of P-gp into the membrane, thereby change substrate specificity. The G1836C SNP has not yet been reported and it isn’t clear about the process or theory impacting egg production traits. However, it is necessary to carry out new experiments for studying material action of PRLR in chicken. Furthermore, the study discovered that egg production traits had significant difference in different generations and the second generation significantly higher than the first generation. It is implied that the selection effect on Erlang Mountainous Chickens’ breeding is significant.
CONCLUSION
In summary, one SNP in exon 6 of PRLR was found in Erlang Mountainous Chicken and there was an association between PRLR genotypes and egg production traits. It is predicted that PRLR may be a potential major gene or in close linkage disequilibrium with QTL for age at first egg of chicken. However, in order to make the selection schemes applicable, it would be necessary to further analyze the effects of PRLR polymorphisms by using populations with different genetic backgrounds and increasing the size of samples.
ACKNOWLEDGMENTS
We thank Jang-Yuan Li for sample collecting and Yao Zhang for data collection. This study was funded by Modern System of Agricultural Industry Technology (CARS-41) and Basic Application Project from Department of Education of Sichuan Province (10ZA144).
REFERENCES
- Alamer, M., 2011. The role of prolactin in thermoregulation and water balance during heat stress in domestic ruminants. Asian J. Anim. Vet. Adv., 6: 1153-1169.
CrossRefDirect Link - Cao, G.Q., B.G. Li, J.H. Liu and W.P. Yang, 2004. Study on relationship between Alu-Polymorphism of Prolactin Receptor (PRLR) gene and litter size in swine. J. Shanxi Agr. Univ., 3: 253-255.
Direct Link - Chen, H.Q., H.Q. Wei, J. Qin and H. Chen, 2011. The novel genetic change in 5'-untranslated region of goose prolactin gene and their distribution pattern in different goose breeds. Asian J. Anim. Vet. Adv., 6: 1069-1075.
CrossRefDirect Link - Chen, H.Q., J. Qin, Y.J. Zhu, Z.T. Pan and Y.N. Xie et al., 2012. The polymorphisms of goat THRSP gene associated with ecological factors in Chinese indigenous goat breeds with different lipogenesis ability. Asian J. Aaim. Vet. Adv., 7: 802-811.
CrossRefDirect Link - Chen, J., H. Liu, Y. Cai, G. Wang, H. Liu and J. Li, 2012. Mutations in the exon 10 of prolactin receptor gene change the egg production performance in Wanjiang white goose. Mol. Biol. Rep., 39: 475-483.
CrossRef - Clarke, D.L. and D.I. Linzer, 1993. Changes in prolactin receptor expression during pregnancy in the mouse ovary. Endocrinology, 133: 224-232.
CrossRefPubMedDirect Link - Cui, J.X., H.L. Du, Y. Liang, X.M. Deng, N. Li and X.Q. Zhang, 2006. Association of polymorphisms in the promoter region of chicken prolactin with egg production. Poult. Sci., 85: 26-31.
CrossRefDirect Link - Drogemuller, C., H. Hamann and O. Distl, 2001. Candidate gene markers for litter size in different German pig lines. J. Anim. Sci., 79: 2565-2570.
PubMedDirect Link - Dunn, I.C., 1998. Genetic mapping of the chicken prolactin receptor gene: A candidate gene for the control of broodiness. Br. Poult. Sci., 39: 23-24.
CrossRefDirect Link - Elferink, M.G., A.A.A. Vallee, A.P. Jungerius, R.P.M.A. Crooijmans and M.A.M. Groenen, 2008. Partial duplication of the PRLR and SPEF2 genes at the late feathering locus in chicken. BMC Genomics, Vol. 9.
CrossRefDirect Link - Hong, K.Y., F. Wang, D.B. Yu and W.X. Du, 2007. Polymorphisms in Taihu chicken of PRL, PRLR and FSHβ genes and association with prophase egg production. Acta Agric. Boreali-Occidentalis Sin., 16: 11-14.
Direct Link - Horseman, N.D., W. Zhao, E. Montecino-Rodriguez, M. Tanaka and K. Nakashima et al., 1997. Defective mammopoiesis, but normal hematopoiesis, in mice with a targeted disruption of the prolactin gene. EMBO J., 16: 6926-6935.
CrossRefPubMedDirect Link - Jenkins, Z., H. Henry, J. Sise and G. Montgomery, 2000. Follistatin (FST), growth hormone receptor (GHR) and prolactin receptor (PRLR) genes map to the same region of sheep chromosome 16. Anim. Genet., 31: 280-280.
CrossRefDirect Link - Jiang, R.S., G.Y. Xu, X.Q. Zhang and N. Yang, 2005. Association of polymorphisms for prolactin and prolactin receptor genes with broody traits in chickens. Poult. Sci., 84: 839-845.
CrossRefPubMedDirect Link - Jie, H, H.D. Yin, Y.P. Liu, Q. Zhu, Y. Zhang and X.L. Zhao, 2010. Correlated research on the early laying performance of erlang mountain chickens. J. Sichuan Agric. Univ., 28: 90-92.
Direct Link - Kelly, P.A., N. Binart, B. Lucas, B. Bouchard and G. Vincent, 2001. Implications of multiple phenotypes observed in prolactin receptor knockout mice. Front. Neuroendocrinol., 22: 140-145.
CrossRefPubMedDirect Link - Jun, L.W., F. Yi, S.Y. Gang, F.G. Xin and T.K. Chuan et al., 2010. A novel 15 bp deletion mutation at KAP16.5 locus in cashmere goat of China. Asian J. Anim. Vet. Adv., 5: 402-409.
CrossRefDirect Link - Putnova, L., A. Knoll, J. Dvorak and S. Cepica, 2002. A new HpaII PCR-RFLP within the porcine prolactin receptor PRLR gene and study of its effect on litter size and number of teats. J. Anim. Breed. Genet., 119: 57-63.
CrossRef - Scott, P., M.K. Kessler and L.A. Schuler, 1992. Molecular cloning of the bovine prolactin receptor and distribution of prolactin and growth hormone receptor transcript in fetal and utero-placental tissues. Mol. Cell. Endocrinol., 89: 47-58.
CrossRefDirect Link - Shirota, M., D. Banville, S. Ali, C. Jolicoeur and J.M. Boutin et al., 1990. Expression of two forms of prolactin receptor in rat ovary and liver. Mol. Endocrinol., 4: 1136-1143.
PubMed - Tanaka, M., K. Maeda, T. Okubo and K. Nakashima, 1992. Double antenna structure of chicken prolactin receptor deduced from the cDNA sequence. Biochem. Bioph. Res. Commun., 188: 490-496.
CrossRef - Van Rens, B.T.T.M. and T. van der Lende, 2002. Litter size and piglet traits of gilts with different prolactin receptor genotypes. Theriogenology, 57: 883-893.
PubMed - Van Rens, B.T.T.M., G.J. Evans and T. van der Lende, 2003. Components of litter size in gilts with different prolactin receptor genotypes. Theriogenology, 59: 915-926.
CrossRefPubMedDirect Link - Qiong, W., F. Chao, L. Wu-Jun, F. Yi and Y. Shi-Gang, 2011. A novel mutation at exon 4 of IGF-1 gene in three indigenous goat breeds in china. Asian J. Anim. Vet. Adv., 6: 627-635.
CrossRefDirect Link - Wells, J.A. and A.M. de Vos, 1996. Hematopoietic receptor complexes. Annu. Rev. Biochem., 65: 609-634.
CrossRef - Xiao, L.H., S.Y. Chen, X.L. Zhao, Q. Zhu and Y.P. Liu, 2011. Association of cellular retinol-binding protein2 (CRBP2) gene polymorphism with egg production in Erlang Mountainous chicken. J. Poultry Sci., 48: 162-167.
CrossRefDirect Link - Xing, G., Q. Zhao, J. Mao, T. Liu and G. Wang, 2011. Identification and characterization of goose prolactin receptor. Poultry Sci., 90: 1050-1057.
CrossRefDirect Link - Zhang, G.X., M.X. Chu, J.Y. Wang, L. Fang and S.C. Ye, 2007. Polymorphism of exon 10 of prolactin receptor gene and its relationship with prolificacy of Jining grey goats. Yi Chuan., 29: 329-336.
PubMedDirect Link - Han, B., H.L. Zhang, L. Zeng, B. Yang, R. Abula, X.L. Xu and Y. Chen, 2011. The Polymorphisms of β2-Adrenergic Receptor Gene on two Cattle Breeds in China Asian J. Anim. Vet. Adv., 6: 715-722.
CrossRef - Kimchi-Sarfaty, C., J.M. Oh, I.W. Kim, Z.E. Sauna, A.M. Calcagno, S.V. Ambudkar and M.M. Gottesman, 2007. A silent polymorphism in the MDR1 gene changes substrate specificity. Sciences, 315: 525-528.
CrossRefDirect Link