
Hong-Quan Chen
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, Peoples� Republic of China
Han-Qin Wei
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, Peoples� Republic of China
Jie Qin
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, Peoples� Republic of China
Hua Chen
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, Peoples� Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 11 | Page No.: 1069-1075







ABSTRACT
To understand prolactin gene variation and its association with egg productivity of goose breeds, the polymorphism of goose prolactin gene and its distribution patterns in different goose breeds were investigated. Six goose breeds selected were Wanxi, Rhine, White Roman, Landaise, Yangzhou and Sichuan. DNA samples of all goose breeds were extracted from goose wing’s blood. The polymorphisms in the 5’-untranslated region of prolactin gene were examined using BstYI enzyme digestion and their distribution patterns were analyzed in different goose breeds. The promoter efficiency of mutational gene was measured using luciferase reporter gene assays. The results showed that a mutation locus, G-10T, located at the bottom nucleotide 10th of the 5’-untranslated region. The G allele frequencies of the six breeds listed above were 0.7857, 0.9778, 0.9493, 0.9054, 0.1479 and 0.2117, respectively. The goose breeds with higher T allele frequencies had higher egg productivity than that with higher G allele frequencies in the practice. The efficiency of promoter with G allele enhanced 26.14% (p<0.01) than that with T allele. The results suggest that the egg productivity of goose breeds may be associated with G-10T locus of prolactin gene.
PDF Abstract XML References Citation
Received: March 18, 2011;
Accepted: August 13, 2011;
Published: September 28, 2011
How to cite this article
Hong-Quan Chen, Han-Qin Wei, Jie Qin and Hua Chen, 2011. The Novel Genetic Change in 5’-untranslated Region of Goose Prolactin Gene and their Distribution Pattern in Different Goose Breeds. Asian Journal of Animal and Veterinary Advances, 6: 1069-1075.
DOI: 10.3923/ajava.2011.1069.1075
URL: https://scialert.net/abstract/?doi=ajava.2011.1069.1075
DOI: 10.3923/ajava.2011.1069.1075
URL: https://scialert.net/abstract/?doi=ajava.2011.1069.1075
INTRODUCTION
In China, there are various goose breeds with egg productivity ranging from 20 to 100 eggs per year (Chen et al., 2004), which makes for significant variations in the geese product usage and affects their breeding and commercial value (Rosinski et al., 2006). Therefore, it is very important to investigate genetic mechanism of the variations.
Prolactin (PRL) can inhibit the development and growth of ovarian follicles, modulate maternal behaviors such as nesting and hatching and affect egg production (Li et al., 2011; Reddy et al., 2006a, 2008). Fang et al. (2005) reported that the PRL level of Wanxi white geese during the nesting period was significantly higher than during egg-laying periods. Reddy et al. (2002) and Eltayeb et al. (2010) found that plasma PRL levels correlated with egg production in hens. There are three promoters in the chicken PRL gene, all of which are located at a locus 278 bp upstream of the DNA coding sequence (CDS) start-point (Kansaku, 2007). These are involved in the modulation of PRL gene transcription (Gerlo et al., 2006) and can change the efficiency of egg production via mutations (Cui et al., 2006). However, the molecular mechanism of PRL gene transcription and translation remains unclear, although many molecular markers correlated with laying performance were found (Gholizadeh and Mianji, 2007; Gholizadeh et al., 2008) of which very few are utilized in breeding practice. Compared with studies in the chicken, duck, turkey and other animals (Chung and Gorewit, 2002; Archawaranon, 2006; Prasad et al., 2007), there are few studies on the goose PRL gene. Only 500 bp of the 5’-flanking region of the PRL gene have been identified (Jiang et al., 2009; Liu et al., 2008) and mutations in this region have not been identified to date.
This study investigated Single Nucleotide Polymorphism (SNP) in 5’-untranslated region (5’-UTR) of the PRL gene, the distribution pattern in goose breeds with different egg productivity.
MATERIALS AND METHODS
Samples: Yangzhou (n = 71), Sichuan (n = 111), Rhine (n = 45), Landaise (n = 37), White Roman (n = 69) and Wanxi (n = 35) from 6 geese breeding farms of Anhui province (Anhui, China) were selected in 2009 and 2010. Genomic DNA from goose wing’s blood was extracted by phenol-chloroform method (Sambrook and Russell, 2001) and then dissolved in TE buffer [10 mmol L-1 Tris-HCl (pH 8.0), 1 mmol L-1 EDTA (pH 8.0)] and kept at -20°C.
The grade of egg productivity (GEP) of six goose breeds were estimated according to the practical egg-yield and less than 20 eggs per year as level 1, then each additional 15 eggs to improve a grade, respectively.
Primers: Based on the sequence of the magang goose PRL gene (GenBank DQ345782.1), a pair of primers was designed using the primer premier 5.0 program (http://www.premierbiosoft.com/) to amplify the fragment containing 5’ partial regulatory region, 5’-UTR, exon 1 and partial intron 1 of goose PRL gene. The sequence length of PCR product was 407 bp in which fragment length (PRL268) of 5’ regulatory region and 5’-UTR was 268 bp. The primers were synthesized by Shanghai Bioengineering Inc. (Shanghai, China).
| • | Upstream primer: 5’TCTCTTCATCAAACCATACTCAG3’ |
| • | Downstream primer: 5’GTTTCTAGAGTTGTTACCTGTCAGT3’ |
PCR conditions and procedures: PCR was carried out using 25 μL PCR amplification kits: 1 μL genomic DNA (50 ng μL-1), 2.5 μL of 10 x buffer with Mg2+, 2 μL dNTP (2 mM), 0.5 μL (2.5 units μL-1) Taq DNA polymerase (Beijing Dingguo Biotechnology Inc. China), 0.5 μL of each primer and 18 μL double-distilled (dd) H2O. Amplification conditions were: initial denaturation of 5 min at 94°C; 35 cycles of 94°C for 30 sec, 56°C for 30 sec, 72°C for 50 sec, followed by 72°C extension for 10 min. The PCR products were detected using 1.5% agarose gel electrophoresis.
Polymorphism analysis: The PCR products digested with BstYI endonucleases (MBI) and then electrophoresed on 3% agarose gel for genotyping polymorphisms. The gene frequencies of the different goose breeds were analyzed statistically. Allele gene frequency, p and q, was calculated by p = D + 0.5H and q = 1-p, respectively where D and H represent homozygote and heterozygote gene frequency, respectively. The distribution patterns of different genotypes were analyzed in the six breeds.
Dual-reporter gene analysis: According to Chen’s protocol (Chen, 2010), the transcriptional efficacy of luciferase gene with different PRL268 promoters in 293T cells were tested and analyzed using dual-reporter gene assays.
Statistical analysis: The PRL268 fragment was subjected to promoter prediction in BDGP (Reese, 2000, 2001). The ratio of firefly luciferase to renilla luciferin intensity was calculated. For the data of non-normal distribution, the values need be transformed using logarithmic transformation (Olivier et al., 2008) and evaluated for p = 0.05 significant differences between different promoters using student’s t-test.
RESULTS
Based on the sequencing results of mixed DNA samples, a mutation locus was detected and located at 10 bp upstream of the CDS start-point (G-10T). Incision of the PCR product by BstYI revealed three genotypes: TT (384/23bp), TG (384/235/149/23bp) and GG (235/149/23bp). The sequence of PRL268 is shown in Fig. 1.
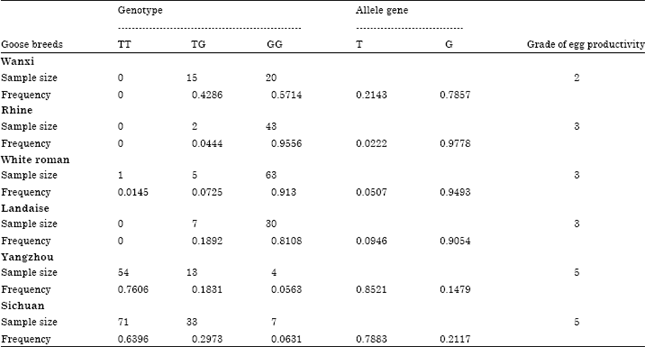
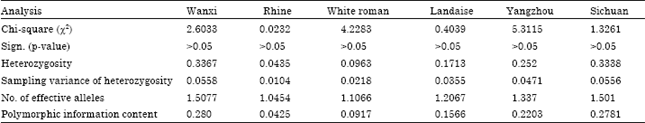
The allele types of G-10T in different breeds were shown in Table 1. The G allele frequencies in Rhine, White Roman and Landaise were all above 0.90 and reached 0.7857 in Wanxi. However, the G allele frequencies in Yangzhou and Sichuan were only 0.1479 and 0.2117, respectively. Hardy-weinberg equilibrium tests (Zhou et al., 2009) and population genetic structure are shown in Table 2. The heterozygosities of Wanxi, Sichuan and Yangzhou were 0.3367, 0.3338 and 0.2520 and their polymorphic information contents were 0.2800, 0.2781 and 0.2203, respectively which showed higher genetic polymorphism comparing with that of Rhine, White Roman and Landaise. The six goose populations all were in Hardy-weinberg equilibrium.
From Table 1, GEPs of Rhine, White Roman, Landaise and Wanxi were listed as lower grade group and higher G allele frequencies, while Yangzhou and Sichuan were referred to as higher grade group and higher T allele frequencies, indicating that the T allele frequency distribution was closely associated with higher egg productivity (Fig. 2). T allele frequencies of higher and lower grade groups were 0.8132 and 0.0833, respectively.
 | |
| Fig. 1: | The sequence and characters of the PRL268 fragment. The italics and bold letters are TATA-box. The capital letters are 5’-UTR of mRNA of prolactin gene. The g/t and G/U are mutation loci. The capital and italics letters are coding region |
| Table 1: | G-10T allele frequencies and grade of egg productivity |
 | |
| Table 2: | Hardy-Weinberg equilibrium tests and population genetic structure |
 | |
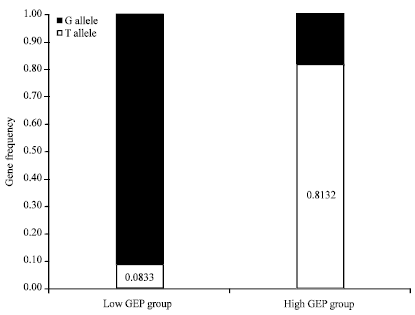 | |
| Fig. 2: | Gene frequency distribution in goose breeds with grade of egg productivity (GEP). Values represent T allele frequency |
| Table 3: | Promoter efficiency of PRL268 using dual reporter gene assays |
 | |
| *Significant difference between means: A>B>C, p<0.01 | |
Table 3 showed the efficacy of PRL268 with T or G allele as promoter of the luciferase reporter gene (PRL268T or PRL268G) in eukaryotic cells. The results shown the promoter efficacy of PRL268G was 5.5140 and increased 26.14% than that of PRL268T (p<0.01), indicating that the T allele affected the transcriptional efficacy of the luciferase reporter gene.
DISCUSSION
There are several reports about the correlation of PRL genotype with avian nesting behavior and egg productivity (Reddy et al., 2006b; Jiang et al., 2009; Eltayeb et al., 2010). A well-known hypothesis is that reduced PRL expression will increase egg productivity. This is consistent with the results that are the Wanxi with higher broodiness and lower GEP in the six goose breeds, followed by the Landaise, White Roman and Rhine with moderate broodiness and GEP and the Yangzhou and Sichuan with the weakest broodiness and higher GEP. However, the linkage between broodiness, egg productivity and PRL gene modulation remains unclear. For a long time, the identifying of the major gene on egg productivity is limited by the population size available (Zou, 2009; Kaiser et al., 2003). Actually, differences in reproductive performance among populations have been reflected in gene frequency via selection pressure. The correlation between GEP and the distribution of PRL gene polymorphisms in six goose breeds showed a consistency in role of PRL gene, although other genes might also have similar effects. In the present study, the distribution pattern of G-10T reflected real differences in GEP. The geese with T allele had higher GEP whereas the geese with G allele exhibited the lower GEP. From the Table 3, the promoter efficacy of PRL268G increased significantly 26.14% than that of PRL268T which showed that T allele might have the role of down regulated transcription of PRL gene while G allele might play the up regulated role. This may be of use in marker assisted selection for the genetic improvement and breed resources protection in goose egg productivity.
Genetic mechanism of T-10G on PRL level and GEP is not clear. The sequence (T/G)ATCTC motif of T-10G locus may serve as the binding of the some transcription factors (Fu and Marzluf, 1990) or ribosomes recognition element in mRNA (Tuller et al., 2009; Baranick et al., 2008). On the other hand, the sequence CAGUAUCUCUACC in 5'-UTR of T allele may produce a potential miRNA targeting locus that is similar to the chicken gga-miR-1688 (Glazov et al., 2008) and then affect the translation of mRNA for PRL. These reviews are in support to the mechanism of T-10G as a marker locus for PRL level and GEP.
CONCLUSION
According to the result of this study, G-10T of prolactin gene is a SNP locus affecting serum PRL level of geese. The T allele frequency is higher in goose breeds with higher GEP. G-10T locus may be associated with egg productivity and broodiness of goose breeds.
ACKNOWLEDGMENTS
The work was supported by The National Key Technology R and D Program of China (2008BADB2B01) and Major Projects of New Varieties of Genetically Modified Organisms of the Ministry of Agriculture (2009ZX08008-007B).
REFERENCES
- Archawaranon, M., 2006. Seasonal changes in gonadotropins, prolactin and sex steroid hormones in captive hill mynah Gracula religiosa. J. Boil. Sci., 6: 375-381.
CrossRefDirect Link - Baranick, B.T., N.A. Lemp, J. Nagashima, K. Hiraoka, N. Kasahara and C.R. Logg, 2008. Splicing mediates the activity of four putative cellular internal ribosome entry sites. Proc. Nat. Acad. Sci., 105: 4733-4738.
CrossRef - Chung, Y. and R.C. Gorewit, 2002. Effects of estrogen and prolactin on bovine BRCA1 gene expression. Pak. J. Nutr., 1: 49-52.
CrossRefDirect Link - Cui, J.X., H.L. Du, Y. Liang, X.M. Deng, N. Li and X.Q. Zhang, 2006. Association of polymorphisms in the promoter region of chicken prolactin with egg production. Poult. Sci., 85: 26-31.
CrossRefDirect Link - Eltayeb, N.M., C.E. Wani and I.A. Yousif, 2010. Assessment of broodiness and its influence on production performance and plasma prolactin level in native chicken of the Sudan. Asian J. Poult. Sci., 4: 1-6.
CrossRefDirect Link - Fang, D.A., Z.Y. Geng, Y. Tao, X.R. Zhang, H. Zhao and X.Y. Chen, 2005. Influence of LHRH-A_3 on WANXI goose reproductive performance and hormone. J. Anhui Agric. Sci., 33: 92-93.
Direct Link - Fu, Y.H. and G.A. Marzluf, 1990. Nit-2, the major positive-acting nitrogen regulatory gene of Neurospora crassa, encodes a sequence-specific DNA-binding protein. Proc. Natl. Acad. Sci. USA., 87: 5331-5335.
PubMed - Gerlo, S., J.R.E. Davis, D.L. Mager and R. Kooijman, 2006. Prolactin in man: A tale of two promoters. Bioessays, 28: 1051-1055.
Direct Link - Gholizadeh, M. and G.R. Mianji, 2007. Use of microsatellite markers in poultry research. Int. J. Poult. Sci., 6: 145-153.
CrossRefDirect Link - Gholizadeh, M., G.R. Mianji and H.S. Zadeh, 2008. Potential use of molecular markers in the genetic improvement of livestock. Am. J. Anim. Vet. Adv., 3: 120-128.
CrossRefDirect Link - Glazov, E.A., P.A. Cottee, W.C. Barris, R.J. Moore, B.P. Dalrymple and M.L. Tizard, 2008. A microRNA catalog of the developing chicken embryo identified by a deep sequencing approach. Genome Res., 18: 957-964.
PubMed - Jiang, R.S., L.L. Zhang, Z.Y. Geng, T. Yang and S.S. Zhang, 2009. Single nucleotide polymorphisms in the 5'-flanking region of the prolactin gene and the association with reproduction traits in geese: Short communication. S. Afr. J. Anim. Sci., 39: 83-87.
Direct Link - Li, W.L., Y. Liu, Y.C. Yu, Y.M. Huang, S.D. Liang and Z.D. Shi, 2011. Prolactin plays a stimulatory role in ovarian follicular development and egg laying in chicken hens. Domestic Anim. Endocrinol., 41: 57-66.
CrossRef - Liu, Z., Z.D. Shi, Y. Liu, M.Y. Li, Y.M. Huang and B.H. Yao, 2008. Molecular cloning and characterisation of the Magang goose prolactin gene. Gen. Comp. Endocrinol., 155: 208-216.
CrossRef - Olivier, J., W.D. Johnson and G.D. Marshall, 2008. The logarithmic transformation and the geometric mean in reporting experimental IgE results: What are they and when and why to use them?. Ann. Allerg. Asthma Immunol., 100: 333-337.
PubMed - Prasad, S.K., T.N. Qureshi, S. Saxena, S. Qureshi, M. Mehar and S.K. Thakur, 2007. L-dopa feeding induces body growth and reproductive conditions in Japanese quail, coturnix coturnix Japonica. Int. J. Poult. Sci., 6: 560-566.
CrossRefDirect Link - Reddy, I.J., C.G. David, P.V. Sarma and K. Singh, 2002. The possible role of prolactin in laying performance and steroid hormone secretion in domestic hen (Gallus domesticus). Gen. Comp. Endocrirnol., 127: 249-255.
CrossRef - Reddy, I.J., C.G. David and S.S. Raju, 2006. Inter sequence pause days, egg production, steroid and luteinizing hormone in domestic hen (Gallous domesticus) immunized against cprolactin. Int. J. Poult. Sci., 5: 420-427.
CrossRefDirect Link - Reddy, G.J., C.G. David and S.S. Raju, 2006. Chemical control of prolactin secretion and it's effects on pause days, egg production and steroid hormone concentration in girirani birds. J. Poult. Sci., 5: 685-692.
CrossRefDirect Link - Reddy, I.J., G.R. Kiran, S.K. Mondal and S. Anandan, 2008. Immunization against vasoactive intestinal peptide and its effects on prolactin, steroid, luteinizing hormone and egg production during mid laying cycle in Punjab brown (PB3) birds. Int. J. Poult. Sci., 7: 1100-1104.
CrossRefDirect Link - Reese, M.G., 2001. Application of a time-delay neural network to promoter annotation in the Drosophila melanogaster genome. Comput. Chem., 26: 51-56.
CrossRefPubMedDirect Link - Rosinski, A., S. Nowaczewski, H. Kontecka, M. Bednarczyk, G. Elminowska-Wenda, H. Bielinska and A. Maczynska, 2006. Analysis of the laying rhythm and reproductive traits of geese. Folia Biol. (Krakow), 54: 145-152.
PubMed - Tuller, T., E. Ruppin and M. Kupiec, 2009. Properties of untranslated regions of the S. cerevisiae genome. BMC Genomics, 10: 391-391.
PubMed - Zou, F., 2009. QTL mapping in intercross and backcross populations. Methods Mol. Biol., 573: 157-173.
PubMed - Zhou, J.J., K. Lange, J.C. Papp and J.S. Sinsheimer, 2009. A heterozygote-homozygote test of Hardy-Weinberg equilibrium. Eur. J. Hum. Genet., 17: 1495-1500.
CrossRefPubMedDirect Link