
Sutarno
Department of Biology, Faculty of Mathematics and Natural Sciences, Sebelas Maret University, Solo, Indonesia
A.D. Setyawan
Department of Biology, Faculty of Mathematics and Natural Sciences, Sebelas Maret University, Solo, Indonesia
A.J. Lymbery
Fish Health Unit and Freshwater Fish Group, School of Veterinary and Life Sciences, Murdoch University, Perth, Australia
Asian Journal of Animal Sciences
Year: 2015 | Volume: 9 | Issue: 2 | Page No.: 57-64







ABSTRACT
The long term objective of this study is to assist in the conservation of Indonesian native cattle breeds which are currently facing serious threats as a result of small population sizes, genetic drift and replacement by imported breeds. The study was conducted to evaluate the genetic diversity within and among five breeds of Indonesian native cattle from Madura, Lombok, Sumbawa, Aceh and Java. Five microsatellite loci were amplified in 40 animals from each breed. A total of 74 alleles were detected over all loci in all of the breeds. The mean observed number of alleles was 3.2 across all loci, with a mean observed heterozygosity of 0.77 and mean expected heterozygosity of 0.53. The observed heterozygote excess may be a result of outcrossing among breeds. Madura cattle were the most genetically distinct of all the breeds studied.
PDF Abstract XML References Citation
Received: January 04, 2015;
Accepted: March 07, 2015;
Published: April 13, 2015
How to cite this article
Sutarno, A.D. Setyawan and A.J. Lymbery, 2015. Genetic Diversity of Five Indonesian Native Cattle Breeds at Microsatellite Loci. Asian Journal of Animal Sciences, 9: 57-64.
DOI: 10.3923/ajas.2015.57.64
URL: https://scialert.net/abstract/?doi=ajas.2015.57.64
DOI: 10.3923/ajas.2015.57.64
URL: https://scialert.net/abstract/?doi=ajas.2015.57.64
INTRODUCTION
Livestock breeds are important components of global biodiversity, particularly because they provide the genetic basis to respond to potential changes in future breeding goals (Groeneveld et al., 2010; Lenstra et al., 2012). Most genetic diversity in livestock is contained within and among native domestic breeds in developing countries which have not usually been subjected to intense artificial selection and as a consequence are locally adapted to a wide range of environments (Giovambattista et al., 2001; Hall and Bradley, 1995). Documenting the genetic diversity of native breeds of livestock has therefore become an important goal in conserving livestock breed diversity and the focus of an increasing number of studies.
Microsatellite loci have proved to be useful markers for the analysis of genetic diversity as they are extremely polymorphic and can be readily amplified and typed by PCR. Microsatellites have been widely used to analyse the genetic diversity of many breeds of cattle, including northern European breeds such as Simmental (Hussein et al., 1996) and Galloway (Herraez et al., 2005), Central European cattle (Citek et al., 2006), Creole cattle of Brazil (Steigleder et al., 2004), Chinese native cattle (Zhou et al., 2005), Indian Kherigarh (Pandey et al., 2006) and Zebu cattle (Sodhi et al., 2006), Hariana and Hissar cattle of Pakistan (Rehman and Khan, 2009), wild gaur in Vietnam (Nguyen et al., 2007) and Aceh cattle of Indonesia (Abdullah et al., 2008). This technique has also been successfully used to analyse the genetic diversity in other livestock species, such pigs (Li et al., 2004; SanCristobal et al., 2006), sheep, goats (Mahmoudi, 2010) and horses (Achmann et al., 2004).
The Indonesian archipelago consists of more than 13,000 islands, covering a range of agro-ecological zones from wetland coastal swamps to semi-arid dry land (Martojo, 2003). Livestock breeds are correspondingly diverse, although their origins are often poorly documented. There are a number of indigenous breeds of cattle in Indonesia, including Bali, Sumba-Ongole, PO, Madura and Aceh cattle. Bali cattle, the most well known are a domesticated form of Banteng (Bos javanicus) (Martojo, 2003; Sudardjat, 2003). Banteng are largely restricted to national parks such as Ujung Kulon and Baluran, while different strains of domesticated Bali cattle are found on different islands. Pure Ongole cattle (Bos indicus) were brought to Sumba Island from India, where they became the Sumba-Ongole (Martojo, 2003; Sudardjat, 2003). Sumba-Ongole were subsequently brought to Java and crossed with Bos javanicus to form the Java-Ongole, from which was developed the PO breed (Sudardjat, 2003). Madura cattle, from the island of Madura are a composite breed, developed from Bos javanicus and Ongole cattle (Sudardjat, 2003). Aceh cattle found throughout the Aceh province of northern Sumatra are believed to have a Bos indicus origin (Payne et al., 1988).
In Indonesia, many native cattle breeds are endangered, with small population sizes and very restricted distributions. Inbreeding and genetic drift in these small, isolated populations as well as increasing economic interest in their replacement by imported breeds, represent very serious threats to the continued survival of many of these native cattle breeds. Information about the genetic status of Indonesian native cattle is essential to develop strategies for their conservation and effective long-term management but to date there have been very few genetic studies of cattle in Indonesia (Abdullah et al., 2008, 2012; Mohamad et al., 2009). The main objectives of this study were to use microsatellite markers to examine genetic variation within and between five breeds of Indonesian native cattle and from these data to infer the genetic structure of each breed and phylogenetic relationships between the breeds.
MATERIALS AND METHODS
Fresh blood samples were collected from 40 animals of different breeds or strains of native cattle from five different Indonesian islands: Lombok (Lombok Bali cattle), Madura (Madura cattle), Sumbawa (Sumbawa Bali cattle), Sumatra (Aceh cattle) and Java (PO cattle). Individual animals of each breed/strain were randomly chosen from a number of herds, without consideration of the relationships among the animals. Blood was collected by venepuncture into a 50 mL tube containing 2.5 mL of 200 mM EDTA as anticoagulant. White blood cells were then isolated from the collected blood.
Whole blood was dispensed into centrifuge tubes and then spun at 1500 g for 15-20 min. The buffy coat was removed with a pipette, transferred to 20 mL centrifuge tubes, topped up with TE-1 buffer (10 mM Tris, 1 mM EDTA, pH 8) and centrifuged at 2000 g for 10-15 min. The pellet was resuspended in 1 mL of TE-2 buffer (10 mM Tris, 1 mM EDTA and 100 mM NaCl, pH 8.0), transferred to a 1 mL Nunc storage tube and frozen at -84°C. The genomic DNA was extracted using the Wizard genomic DNA purification system (Promega, Madison, WI, USA).
Five microsatellite loci were amplified, using the following primers:
| • | BM 1824: | CGAGCAAGGTGTTTTTCCAATC CATTCTCCAACTGCTTCCTTG |
| • | ETH 225: | GATCACCTTGCCACTATTTCCT ACATGACAGCCAGCTGCTACT |
| • | INRA 005: | CAATCTGCATGAAGTATAAATAT CTTCAGGCATACCCTACACC |
| • | MM 12: | CAAGACAGGTGTTTCAATCT ATCGACTCTGGGGATGATGT |
| • | TGLA 227: | CGAATTCCAAATCTGTTAATTTGCT ACAGACAGAAACTCAATGAAAGCA |
Polymerase Chain Reaction (PCR) was carried out on 100 ng of genomic DNA in a 50 μL reaction volume. The reaction mixture consisted of 200 μM each of dATP, dCTP, dGTP and dTTP, 50 mM KCl, 10 mM TrisHCl (pH 9.0), 0.1% Triton X-100, 1.5 mM MgCl2, 1 unit of Taq DNA polymerase and 4 ng of each primer. The PCR protocol involved an initial denaturation at 95°C for 2 min, followed by 30 cycles of 92°C (1 min), 55°C (45 sec) and 72°C (1 min). An additional elongation step of 10 min was carried out at 72°C.
Ten microliter of PCR products were loaded on to a 2% agarose gel, electrophoresed and visualized under UV light after ethidium bromide staining. A microsatellite primer pair was scored as positive by detection of a discrete band. To test for the presence of polymorphism, the PCR products were resolved on 6% denaturing polyacrylamide gels (Sequi-GT system, Bio-Rad, Richmond, USA). A pGEM DNA marker and four base-pair allelic ladder (Promega, Madison, USA) were used as size standards for sizing PCR products. To visualize the resolved PCR products, gels were stained using a silver staining kit (Promega, Madison, USA). The genotypes were scored manually.
Genotypes were assigned for each individual based on allele size data. Genotype and allele frequencies were calculated for all breeds at all microsatellite loci. The Polymorphism Information Content (PIC) was calculated for each locus using the formula of Botstein et al. (1980):
![]()
where, pi and pj are frequencies of ith and jth alleles.
Genetic diversity within breeds was described by the observed number of alleles (Na), effective number of alleles (Ne) (Kimura and Crow 1964), observed heterozygosity (Ho) and expected heterozygosity (He) (Nei, 1978) at each locus, computed using Microsatellite Toolkit version 3.1. Deviations of genotype frequencies from Hardy-Weinberg equilibrium were tested by the exact test of Guo and Thompson (1992), with a Bonferroni correction to obtain an experiment-wide type I error rate of 5%. The extent of the deviation was expressed for each locus by Wright’s fixation index (F) and across all loci for each breed, F-values were summarized by the weighted mean, FIS, calculated using Genepop 4.1.
Genetic differentiation among breeds was assessed by Wright’s FST (Wright, 1951) calculated by the method of Weir and Cockerham (1984) and implemented in Genepop 4.1. Genetic identities and distances among all pairwise combination of breeds were calculated by the method of Nei et al. (1983) and used to construct a phylogram by the neighbor-joining method, applying NTSYSpc version 2.02i.
RESULTS AND DISCUSSION
Microsatellite loci were successfully amplified in all five breeds of Indonesian native cattle and measures of genetic diversity for each locus over all breeds are shown in Table 1. All loci were polymorphic with a mean of 4.2 alleles observed per locus and a mean expected heterozygosity of 0.55 per locus. The loci ETH225, INRA005 and MM12 appear to be particularly informative for breed characterization and potentially, for individual identification (PIC>0.5, Botstein et al., 1980).
Measures of genetic diversity for each breed are shown in Table 2. For most loci, in most breeds, there were more heterozygotes than expected. Heterozygote deficiencies are more commonly found than heterozygote excesses in domesticated livestock and are usually ascribed to the effects of inbreeding, resulting from small population sizes and genetic improvement programs (Martinez et al., 2000; Mukesh et al., 2004; SanCristobal et al., 2006).
Heterozygote excesses in natural populations are usually explained by direct or associative overdominant selection (Mitton, 1989; Nei, 1987) negative assortative mating (Hartel and Clark, 1989) or divergent allele frequencies in male and female parents when a small number of breeders produce the next generation (Pudovkin et al., 1996). Overdominance is usually locus-specific and is therefore unlikely to explain the effects found across all microsatellite loci in the present study. Negative assortative mating through outcrossing with other breeds or binomial sampling error in allele frequencies from using a small number of breeders are more likely explanations. At this stage there are no information which would enable us to distinguish these possibilities, although there is anecdotal evidence of many past crosses among Indonesian cattle breeds (Martojo, 2003; Mohamad et al., 2009).
For polymorphic loci, the mean number of effective alleles per locus ranged from 1.1-4.2 and the mean expected heterozygosity from 0.05 to 0.78 (Table 2) which is a similar level of diversity to that found in other native breeds of cattle from Asia and Africa (Abdullah et al., 2008; MacHugh et al., 1997; Mohamad et al., 2009; Mukesh et al., 2004) but greater than that typically found in European cattle breeds (Hanslik et al., 2000; MacHugh et al., 1994, 1997). Among the five native Indonesian breeds of cattle that were studied, genetic diversity was greatest in Madura cattle and least in Bali cattle from Sumbawa, with moderate levels in PO cattle, Aceh cattle and Bali cattle from Lombok.
| Table 1: | Measures of genetic diversity (Na), observed number of alleles for five microsatellite loci over five breeds of Indonesian native cattle |
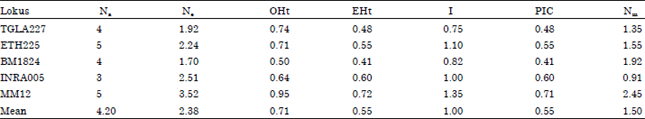 | |
| Ne: Effective number of alleles; Ho: Observed heterozygosity; He: Expected heterozygosity; PIC: Polymorphism information content) Na: Observed number of alleles, Ne: Effective number of alleles [Kimura and Crow (1964)], I: Shannon’s Information index [Lewontin (1972)], OHm: Observed Homozygosity, EHm: Expected Homozygosity, EHt: Expected Heterozygosity, OHt: Observed Heterozygosity. PIC: Polymorphism Information Content, Nm: Gene Flow Estimated from Fst = 0,25 (1-Fst)/Fst | |
| Table 2: | Measures of genetic diversity and fixation indices (F) at five microsatellite loci within five breeds of Indonesian native cattle |
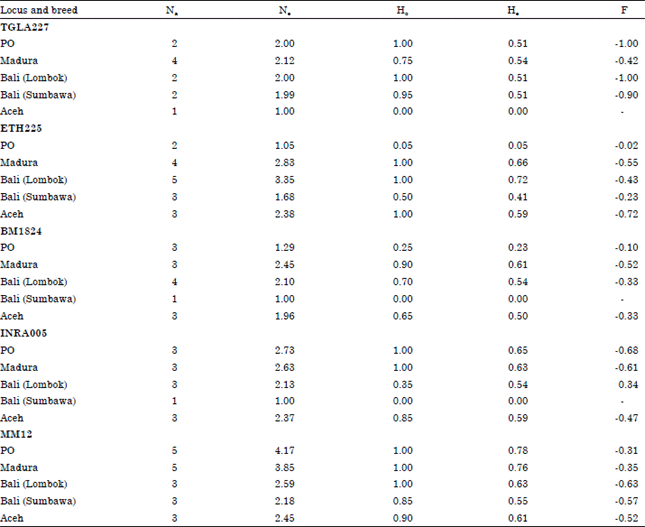 | |
| Na: Observed number of alleles, Ne: Effective number of alleles, Ho: Observed heterozygosity, He: expected heterozygosity | |
Mohamad et al. (2009) also found higher levels of genetic diversity at microsatellite loci in Madura cattle than in a number of other Indonesian breeds which may reflect its composite background, although PO cattle are presumed to be similarly derived from a cross between Bos indicus and Bos javanicus cattle (Sudardjat, 2003).
The mean FST among populations over all loci was 0.143, indicating that over 14% of total genetic diversity was found among the five different breeds. The dendrogram of relationships is presented in Fig. 1.
As expected, the two Bali cattle populations were most closely related as also found by other studies with Aceh and PO breeds also clustered together. These results are similar to those found by Mohamad et al. (2009) and Abdullah et al. (2012), using mitochondrial, Y-chromosome and microsatellite markers and indicate that Aceh cattle may share a similar origin to PO cattle which resulted from the crossing of Indian Ongole (Bos indicus) and Javan Bali cattle (Bos javanicus). Madura cattle, although also of Bos indicusxBos javanicus origin are quite distinct. Madura has a long tradition of cattle breeding and the Madura breed appears to have been established for a long period of time (Mohamad et al., 2009; Nijman et al., 2003; Popescu and Smith, 1988).
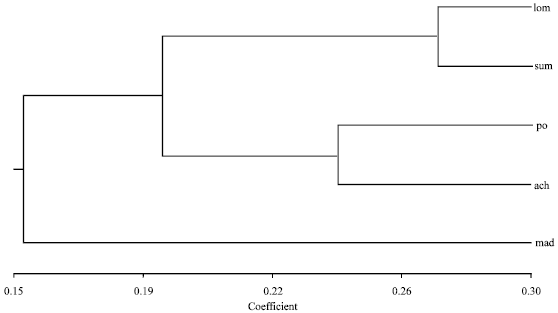 | |
| Fig. 1: | Dendrogram of genetic relationship among five native Indonesian cattle breeds: Madura (mad), Aceh (ach), PO (po), Sumbawa Bali (sum) and Lombok Bali (lon) |
CONCLUSION
In conclusion, our results, combined with previous studies on Indonesian native cattle breeds indicate similar levels of within-breed genetic diversity to that found in other breeds throughout Asia and Africa and substantial among-breed differentiation. There is a need now to use quantitative genetic breeding studies and/or genome-wide molecular analyses to determine the extent to which this genetic distinctiveness at neutral marker loci reflects genetic differences in production traits and local adaptation to different environments, so that we can best conserve these unique genetic resources.
ACKNOWLEDGMENTS
Thanks to the International office of UNS for providing financial support for the research, to Nina Astreani for the dendrogram analysis and Nina Kurnianingrum, Dian, Pramesti, Septiana and Prasasti for assistance in the laboratory.
REFERENCES
- Abdullah, M.A.N., R.R. Noor, H. Martojo and D.D. Solihin, 2008. Genetics characterization of aceh cattle utilizing microsatellite DNA analyses. J. Indonesian Trop. Anim. Agric., 33: 165-175.
Direct Link - Achmann, R., I. Curik, P. Dovc, T. Kavar and I. Bodo et al., 2004. Microsatellite diversity, population subdivision and gene flow in the Lipizzan horse. Anim. Genet., 35: 285-292.
CrossRefPubMedDirect Link - Botstein, D., R.L. White, M. Skolnick and R.W. Davis, 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet., 32: 314-331.
PubMedDirect Link - Citek, J., L. Panicke, V. Rehout and H. Prochazkova, 2006. Study of genetic distances between cattle breeds of Central Europe. Czech J. Anim. Sci., 51: 429-436.
Direct Link - Giovambattista, G., M.V. Ripoli, P. Peral-Garcia and J.L. Bouzat, 2001. Indigenous domestic breeds as reservoirs of genetic diversity: The Argentinean Creole cattle. Anim. Genet., 32: 240-247.
CrossRefDirect Link - Groeneveld, L.F., J.A. Lenstra, H. Eding, M.A. Toro and B. Scherf et al., 2010. Genetic diversity in farm animals: A review. Anim. Genet., 41: 6-31.
CrossRefDirect Link - Guo, S.W. and E.A. Thompson, 1992. Performing the exact test of Hardy-Weinberg proportion for multiple alleles. Biometrics, 48: 361-372.
PubMedDirect Link - Hall, S.J.G. and D.G. Bradley, 1995. Conserving livestock breed biodiversity. Trends. Ecol. Evol., 10: 267-270.
CrossRef - Hanslik, S., B. Harr, G. Brem and C. Schlotterer, 2000. Microsatellite analysis reveals substantial genetic differentiation between contemporary New World and Old World Holstein Friesian populations. Anim. Genet., 31: 31-38.
CrossRef - Herraez, D.L., H. Schafer, J. Mosner, H.R. Fries and M. Wink, 2005. Comparison of microsatellite and single nucleotide polymorphism markers for the genetic analysis of a Galloway cattle population. Zeitschrift Naturforschung C, 60: 637-643.
Direct Link - Hussein, A.A., F. Schmoll, F. Fuhrer, G. Brem and K. Schellander, 1996. Evaluation of seven microsatellite loci in Simmental cattle. J. Vet. Med. Ser. A, 43: 1-8.
CrossRef - Kimura, M. and J.F. Crow, 1964. The number of alleles that can be maintained in a finite population. Genetics, 49: 725-738.
PubMedDirect Link - Lenstra, J.A., L.F. Groeneveld, H. Eding, J. Kantanen and J.L. Williams et al., 2012. Molecular tools and analytical approaches for the characterization of farm animal genetic diversity. Anim. Genet., 43: 483-502.
CrossRef - Li, S.J., S.H. Yang, S.H. Zhao, B. Fan and M. Yu et al., 2004. Genetic diversity analyses of 10 indigenous Chinese pig populations based on 20 microsatellites. J. Anim. Sci., 82: 368-374.
PubMed - MacHugh, D.E., R.T. Loftus, D.G. Bradley, P.M. Sharp and P. Cunningham, 1994. Microsatellite DNA variation within and among European cattle breeds. Proc.: Biol. Sci., 256: 25-31.
Direct Link - MacHugh, D.E., M.D. Shriver, R.T. Loftus, P. Cunningham and D.G. Bradley, 1997. Microsatellite DNA variation and the evolution, domestication and phylogeography of taurine and zebu cattle (Bos Taurus and Bos indicus). Genetics, 146: 1071-1086.
Direct Link - Martinez, A.M., J.V. Delgado, A. Rodero and J.L.Vega-Pla, 2000. Genetic structure of the Iberian pig breed using microsatellites. Anim. Genet., 31: 295-301.
Direct Link - Martojo, H., 2003. Indigenous Bali Cattle: The Best Suited Cattle Breed for Sustainable Small Farming in Indonesia. In: The Relationship between Indigenous Animals and Humans in APEC Region, Chang, H.L. and Y.C. Huang (Eds.). Chinese Society of Animal Science, Taiwan, ISBN-13: 9789573020721, pp: 21-36.
- Mohamad, K., M. Olsson, H.T. van Tol, S. Mikko and B.H. Vlamings et al., 2009. On the origin of Indonesian cattle. PloS One, Vol. 4.
CrossRef - Mahmoudi, B., 2010. Genetic diversity of lori goat population based on microsatellite marker. Asian J. Anim. Sci., 4: 13-19.
CrossRefDirect Link - Mukesh, M., M. Sodhi, S. Bhatia and B.P. Mishra, 2004. Genetic diversity of Indian native cattle breeds as analysed with 20 microsatellite loci. J. Anim. Breed. Genet., 121: 416-424.
CrossRefDirect Link - Nei, M., 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583-590.
PubMedDirect Link - Nei, M., F. Tajima and Y. Tateno, 1983. Accuracy of estimated phylogenetic trees from molecular data: II. Gene frequency data. J. Mol. Evol., 19: 153-170.
CrossRefPubMedDirect Link - Nguyen, T.T., S. Genini, L.C. Bui, P. Voegeli and G. Stranzinger et al., 2007. Genomic conservation of cattle microsatellite loci in wild gaur (Bos gaurus) and current genetic status of this species in Vietnam. BMC Genet., Vol. 8.
CrossRef - Pandey, A.K., R. Sharma, Y. Singh, B.B. Prakash and S.P.S. Ahlawat, 2006. Genetic diversity studies of Kherigarh cattle based on microsatellite markers. J. Genet., 85: 117-122.
CrossRefDirect Link - Payne, R.C., D.E. Ward, M. Usman, A. Rusli, D. Djauhari and A. Husein, 1988. Prevalence of bovine haemoparasites in Aceh Province of Northern Sumatra: Implications for imported cattle. Prev. Vet. Med., 6: 275-283.
CrossRefDirect Link - Popescu, C.P. and W.G. Smith, 1988. A cytogenetic investigation of Madura cattle. Reprod. Domest. Anim., 23: 145-148.
CrossRefDirect Link - Pudovkin, A.I., D.V. Zaykin and D. Hedgecock, 1996. On the potential for estimating the effective number of breeders from heterozygote-excess in progeny. Genetics, 144: 383-387.
PubMed - Rehman, M.S. and M.S. Khan, 2009. Genetic diversity of Hariana and Hissar cattle from Pakistan using microsatellite analysis. Pak. Vet. J., 29: 67-71.
Direct Link - SanCristobal, M., C. Chevalet, C.S. Haley, R. Joosten and A.P. Rattink et al., 2006. Genetic diversity within and between European pig breeds using microsatellite markers. Anim. Genet., 37: 189-198.
CrossRef - Sodhi, M., M. Mukesh, B. Prakash, S.P.S. Ahlawat and R.C. Sobti, 2006. Microsatellite DNA typing for assessment of genetic variability in Tharparkar breed of Indian zebu (Bos indicus) cattle, a major breed of Rajasthan. J. Genet., 85: 165-170.
CrossRefDirect Link - Steigleder, C.S., E.A. Almeida and T.A. Weimer, 2004. Genetic diversity of a Brazilian creole cattle based on fourteen microsatellite loci. Archivos Zootecnia, 53: 3-11.
Direct Link - Weir, B.S. and C.C. Cockerham, 1984. Estimating F-statistics for the analysis of population structure. Evolution, 38: 1358-1370.
CrossRefDirect Link - Wright, S., 1949. The genetical structure of populations. Ann. Eugenics, 15: 323-354.
CrossRefPubMedDirect Link - Zhou, G.L., H.G. Jin, Q. Zhu, S.L. Guo and Y.H. Wu, 2005. Genetic diversity analysis of five cattle breeds native to China using microsatellites. J. Genet., 84: 77-80.
PubMedDirect Link