
M. F. Ahmed
Department of Agronomy, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
Asian Journal of Crop Science
Year: 2013 | Volume: 5 | Issue: 1 | Page No.: 81-94







ABSTRACT
The study was undertaken to estimate heterosis and combining ability of new maize inbred lines in F1 combinations for some agronomic traits under two sowing dates for developing superior hybrids. Also this investigation studied the possibility of predicting heterosis and combining ability in maize inbred lines via protein electrophoresis. A half diallel set of crosses involving eight maize inbred lines were evaluated under two different sowing dates and the inbred lines were used for protein electrophoresis study. The obtained data revealed highly significant differences between sowing dates for all studied traits except number of rows per ear. The mean squares due to General Combining Ability (GCA) and Specific Combining Ability (SCA) were significant for all studied traits under both planting dates except 100 kernel weight under late planting date for SCA. GCA/SCA ratios were less than unity for plant height, days to 50% silking, 100 kernel weight and grain yield per plant under normal sowing date and ear height, plant height, number of kernel per row, 100 kernel weight and grain yield per plant under late sowing date and the other traits were more than unity. The hybrids P1xP2, P1xP6, P1xP8, P2xP5, P2xP6, P3xP6, P5xP6, P5xP7 and P6xP7 under normal sowing date and P2xP4, P2xP5, P4xP6 and P5xP6 under late sowing date out yielded the check hybrid. The parental lines P2, P5 and P6 seemed to be the best combiners for grain yield per plant and most of the studied traits under normal and late planting dates. Twenty one out of the twenty eight crosses showed desirable significant SCA effects under normal and late sowing dates for grain yield per plant and most of the other studied traits. The electrophoresis patterns could be useful tools for the identification and characterization of these inbred lines and could be used as biochemical genetic markers associated with hybrid vigor and SCA in maize.
PDF Abstract XML References Citation
Received: July 08, 2012;
Accepted: August 25, 2012;
Published: October 13, 2012
How to cite this article
M. F. Ahmed, 2013. Diallel Analysis and Biochemical Genetic Markers for Heterosis and Combining Ability under Two Sowing Dates of Maize Inbred Lines. Asian Journal of Crop Science, 5: 81-94.
DOI: 10.3923/ajcs.2013.81.94
URL: https://scialert.net/abstract/?doi=ajcs.2013.81.94
DOI: 10.3923/ajcs.2013.81.94
URL: https://scialert.net/abstract/?doi=ajcs.2013.81.94
INTRODUCTION
One of the most important cultural practices is sowing date. It has an active role effect on the growth and development of maize plants. Delay or early planting was reported to affect the growth and yield of maize significantly (Lauer et al., 1999; Nielson et al., 2002; Beiragi et al., 2011). The suitable date for maize sowing mainly depends on many factors such as weather conditions (sunshine, temperature and humidity). Either early planting or late planting can result in lower yield because the probability exists that unfavorable climatic conditions can occur after planting or during the growing season. In Egypt maize is planted successfully under irrigation from mid April to mid August, although most of the area is planted between mid May to mid June as optimum period for high or good production, whereas grain yield decreases after this date. The maize breeder is concerned with estimating genetic parameters and their interaction under different environmental conditions to choose the most effective breeding program for his materials. The parents of the best potentiality to transmit desirable traits to their progenies are those exhibiting the highest value for general combining ability effects, whereas combinations of highest specific combining ability effects demonstrate exploitation of heterosis concept. Therefore, many research workers studied heterosis and combining ability under different sowing dates and the interaction between them (Sedhom, 1994; Al-Ahmad et al., 2004; El-Shouny et al., 2008; Younis et al., 2010).
The electrophoretic patterns (SDS-PAGE) for water soluble proteins has been used as biochemical genetic makers associated with heterosis and combining ability. Several investigators (Abdel-Tawab et al., 1989; Abdel-Sattar and Ahmed, 2004; Hosni et al., 2006; El-Shouny et al., 2011) tried to identify and characterize the parental lines of maize using proteins electrophoresis. The present investigation aimed to; (1) evaluate eight maize inbred lines and their 28 F1 hybrids in half diallel cross for heterosis and combining ability under normal and late sowing dates to identify the high GCA lines that could be used as parental lines in breeding program for specific traits and to identify promising hybrids with high SCA that could be used commercially under different sowing dates and (2) study the possibility of predicting heterosis and combining ability in maize via protein electrophoresis.
MATERIALS AND METHODS
Genetic materials: New eight white maize (Zea mays L.) inbred lines (P1, P2, P3, P4, P5, P6, P7 and P8), representing a wide range of diversity for several agronomic characters. These inbred lines in the S8 generation were developed from different sources (P1, P3, P4 and P7 from G.2 variety) and (P2, P5, P6 and P8 from D.C. 215 variety) by Prof. Dr. K.A. El-Shouny through a breeding program at Agronomy Department, Fac. of Agric., Ain Shams Univ. In 2010 season, all possible cross combinations excluding reciprocals were made among the eight inbred lines giving a total of 28 F1 crosses.
Field trials: In 2011 growing season, the seeds of the eight inbred lines, their twenty eight F1 crosses and the check variety Single Cross 10 (S.C.10) were sown at two planting dates, i.e. 21st of May (normal or recommended planting) and 1st of July (late planting) at the Agric. Res. Stat. Fac. of Agric., Ain Shams Univ., Shalakan, Kalubia Governorate, Egypt. A field experiment laid out in a randomized complete block design with three replications was devoted for each planting date. Each experimental plot included one row of four meters long and 70 cm wide. Planting was in hills spaced at 25 cm apart and hills were thinned at one plant per hill. The common agricultural practices of growing maize were applied properly as recommended in the district. Data were recorded on 10 guarded plants for ear height (cm), plant height (cm), days to 50% anthesis, days to 50% silking, number of kernels/row, number of rows/ear, 100 kernel weight (g) and grain yield per plant (g).
Laboratory trials: Two leaves of eight maize inbred lines were used for SDS-protein analysis. Sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed on water soluble protein fractions (albumin and globulin) according to the method of Laemmli (1970) as modified by Studier (1973). The SDS-protein gel was scanned and analyzed using Gel Doc 2000 Bio-Rad System.
Statistical analysis: An ordinary analysis of variance (p<0.01) for each sowing date and the combined analysis over the two sowing dates was performed according to Snedecor and Cochran (1981) General and specific combining ability variances and effects were obtained by employing (Griffing, 1956) diallel cross analysis method 2 model I at each Planting date (SAS program). Percentage of heterosis over the check variety was estimated according to Wynne et al. (1970).
RESULTS AND DISCUSSION
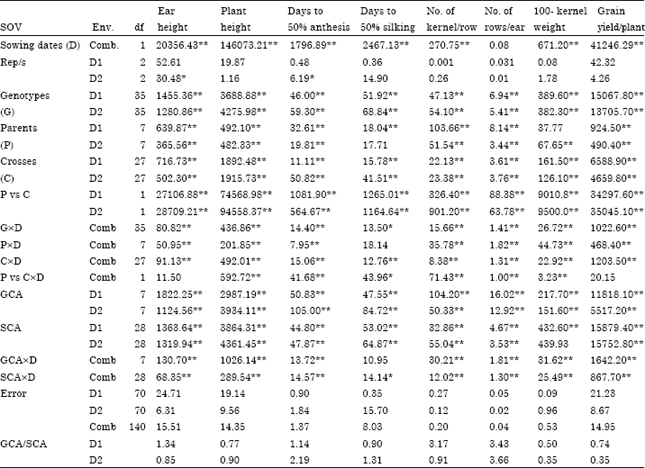
Analysis of variance: Mean square estimates for all studied traits under normal and late sowing dates are presented in Table 1. Highly significant differences (p<0.01) between sowing dates were found for all studied traits except number of rows per ear, indicating that the genotypes performance differed under the two sowing dates.
Mean squares for genotypes, parents and crosses showed significant differences for all recorded traits under both sowing dates except 100 kernel weight and days to 50% silking for parents under normal and late sowing dates, respectively. This indicates that variability exists among these populations may increase the chance of good new recombination that can be isolated in the succeeding generations. Parents vs. crosses mean squares as an indication for average heterosis over all crosses were significant for all studied traits under normal and late sowing dates. The interaction of genotypes, parents and crosses with both sowing dates were highly significant for all studied traits except days to 50% silking for parentsxEnv., indicating inconsistent responses of these populations from normal to late sowing date. Moreover, mean square for Parents vs crossesxEnv. was significant for all recorded traits except ear height and grain yield per plant.
Mean performance and heterosis over check variety: Mean performance for all studied traits under normal and late sowing dates is presented in Table 2.
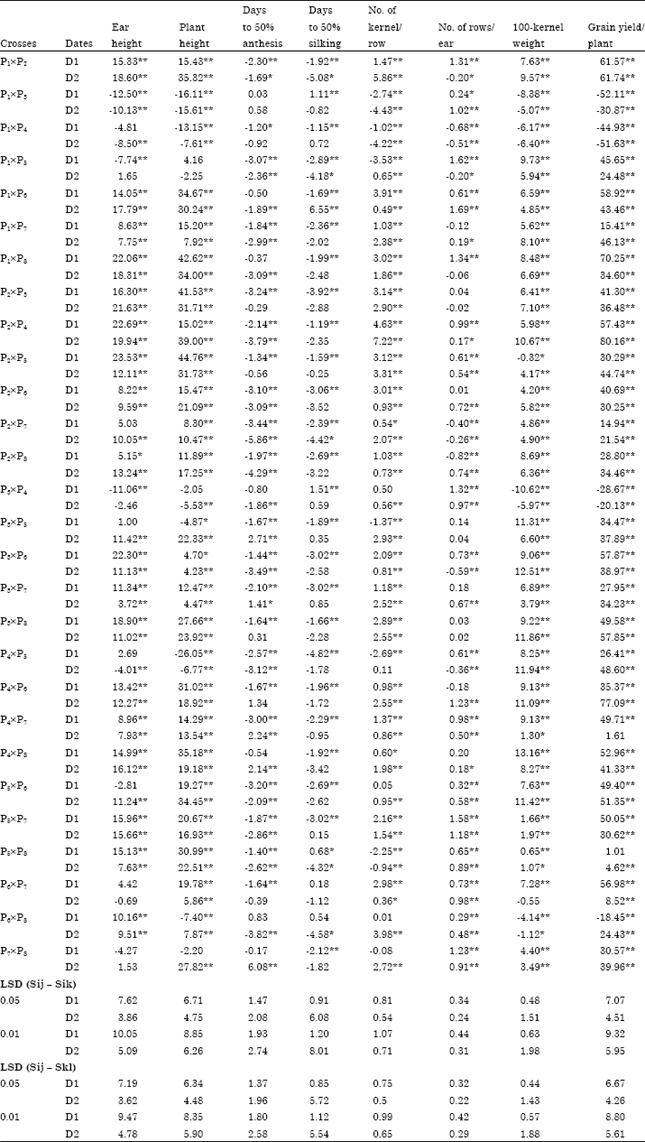
| Table 1: | Mean squares estimates for studied traits in 8x8 white maize diallel crosses under normal (D1) and late (D2) sowing dates |
 | |
| *,**Significance at 0.05 and 0.01 probability level, respectively | |
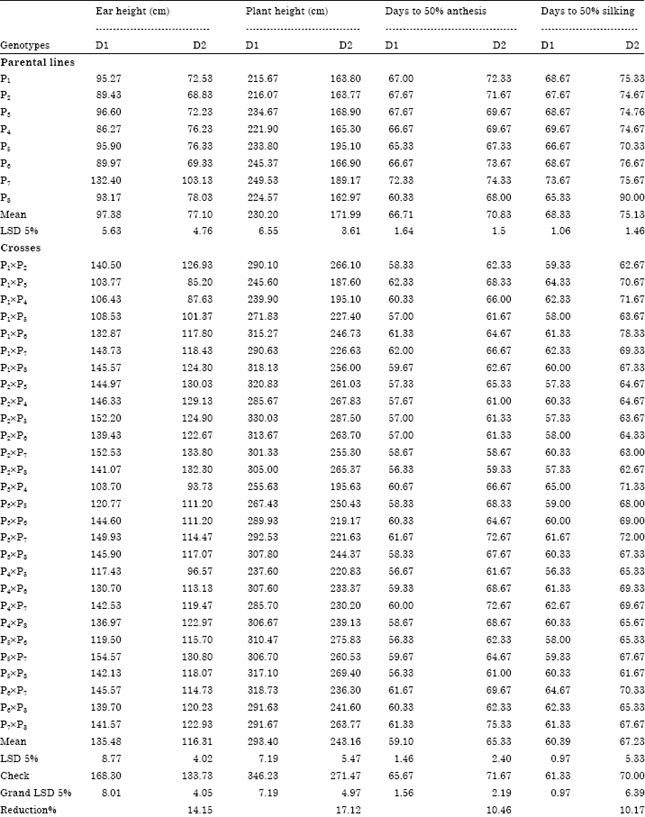
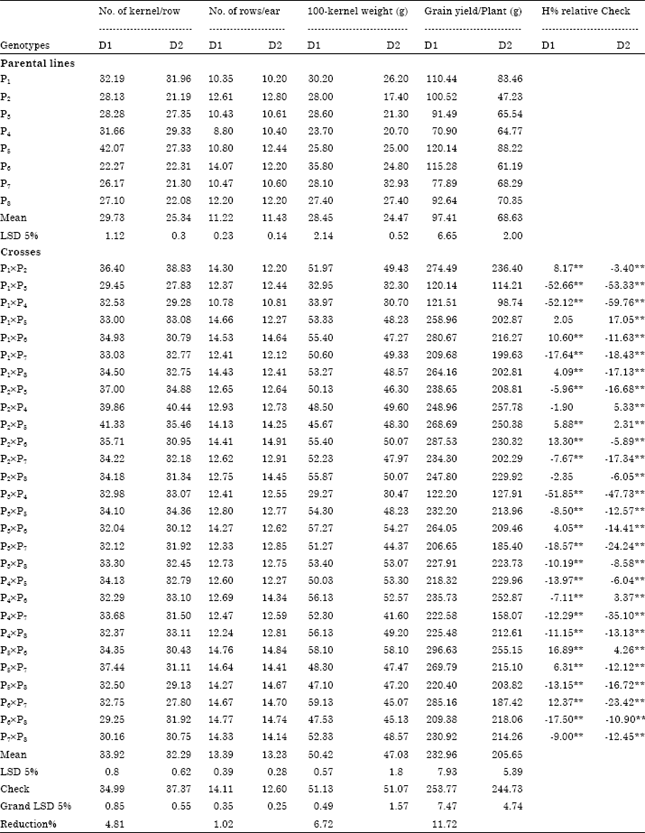
| Table 2: | Mean performance of the eight maize inbred lines and their crosses under normal D1) and late (D2) sowing dates and heterosis relative to check variety S.C.10 for grain yield per plant |
 | |
 | |
| *,**Significance at 0.05 and 0.01 probability levels, respectively | |
Mean values for these traits exhibited the parental diversity and the hybrid differential response from normal to late sowing date. The inbred line P5 ranked the first as the best values under normal and late sowing dates for grain yield per plant (120.14 and 88.22 g, respectively) and most of the studied traits. The inbred line P6 ranked the second under normal sowing date for grain yield per plant (115.28 g) and most of the studied traits. The inbred line P1 ranked the third under normal (110.44 g) and the second under late sowing dates for grain yield per plant (83.46 g) and some of the other studied traits.
The inbred line P7 was the best under both conditions for ear (132.40 and 103.13 cm, respectively) and plant (249.53 and 189.17 cm, respectively) heights. The inbred line P8 was the best under normal sowing date for days to 50% anthesis (60.33 days) and silking (65.33 days).
The studied traits exhibited the best performance at early planting date and then began to decrease with delay in planting date. Reduction on late planting was as much as in plant height which reached 17.12% relative to early planting date. Ear height, grain yield per plant, days to 50% anthesis and days to 50% silking were most affected by delaying planting recording 14.15%, 11.72%, 10.46% and 10.17% reduction, respectively. The least affected by delaying of planting were 100 kernel weight recording 6.72% reduction followed by No. of kernel per row (4.81% reduction) and No. of rows per ear (1.02% reduction), suggesting that these traits are less sensitive to delay in planting date. The hybrid P5xP6 was the best in grain yield per plant and most of the studied traits under both conditions. The hybrids P1xP2, P1xP6, P1xP8, P2xP5, P2xP6, P3xP6, P5xP6, P5xP7 and P6xP7 under normal sowing date and the hybrids P2xP4, P2xP5, P4xP6 and P5xP6 under late sowing date exceeded the check variety S.C.10 for grain yield per plant. The hybrids P4xP6, P4xP8 and P6xP7 under normal conditions and the hybrids P3xP6, P3xP8 and P4xP5 under late sowing date exceeded the check variety S.C.10 for 100 kernel weight. The hybrids P2xP8, P5xP6 and P5xP8 for days to 50% anthesis, P2xP3, P2xP5, P2xP8 and P4xP5 for days to 50% silking, P2xP3, P2xP4 and P2xP5 for number of kernel per row and P1xP6, P5xP6 and P6xP8 for number of rows per ear under normal conditions exceeded the check variety S.C.10. The hybrids P2xP7 and P2xP8 for days to 50% anthesis, P1xP2, P2xP8 and P5xP8 for days to 50% silking, P1xP2 and P2xP4 for number of kernels per row and P2xP6, P5xP6, P5xP7 and P6xP8 for number of rows per ear under late sowing date exceeded the check variety S.C.10 (Table 2).
Heterosis expressed as the percentage deviation of F1 mean performance from check variety S.C.10 values for grain yield per plant are presented in Table 2. Concerning grain yield per plant the hybrids P1xP2, P1xP6, P1xP8, P2xP5, P2xP6, P3xP6, P5xP6, P5xP7 and P6xP7 under normal sowing date and the hybrids P2xP4, P2xP5, P4xP6 and P5xP6 under late sowing date out yielded the check hybrid S.C.10. Hence, it could be concluded that these crosses offer possibility for improving grain yield in maize. Many investigators reported high heterosis for yield of maize; i.e., Shafey et al. (2003), Soliman et al. (2005), El-Hosary et al. (2006) and Aliu et al. (2008).
Combining ability: The analysis of variance for combining ability under normal and late sowing dates for all the studied traits is presented in Table 1. The variance of general (GCA) combining ability includes the additive and additivexadditive genetic portion, while specific (SCA) combining ability represents the non-additive genetic portion of the total variance arising largely from dominance and epistatic deviation. The mean squares due to GCA and SCA were significant for all the studied traits under both planting dates except 100 kernel weight under late planting date for SCA. Indicating that both additive and non-additive genetic effects were involved in the inheritance of these traits under both planting dates. When both general and specific combining ability mean squares are significant, one may ask which type and or types of gene action are important in determining the performance of single-cross progeny. To overcome such situation the size of mean squares can be used to assume the relative importance of general and specific combining ability mean squares which were highly significant. Hence, GCA/SCA ratio was used as measure to reveal the nature of genetic variance involved. For days to 50% anthesis and No. of rows per ear under both environments, ear height and number of kernel per row under normal planting date and days to 50% silking under late planting date, high ratios which largely exceeded the unity were obtained, indicating that a large part of the total genetic variability associated with these traits was a result of additive and additive by additive gene action. Al-Ahmad et al. (2004), Soliman et al. (2005) and El-Shouny et al. (2011) came to the same conclusion. Plant height, days to 50% silking, 100 kernel weight and grain yield per plant under normal sowing date and ear height, plant height, number of kernel per row, 100 kernel weight and grain yield per plant under late sowing date exhibited GCA/SCA ratios less than unity. Therefore, it could be concluded that the large portion of the total genetic variability for these traits was due to non-additive gene action. The genetic variance was previously reported by Abdel-Sattar and Ahmed (2004), El-Hosary et al. (2006), El-Shouny et al. (2008) and Younis et al. (2010).
The mean squares of interaction between planting dates and both types of combining ability were significant for all studied traits except days to 50% silking for GCAxD indicating that both additive and non-additive gene effects were influenced by seasonal changes.
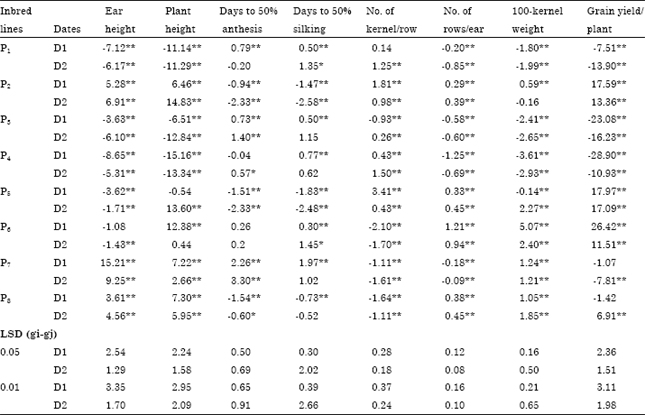
General combining ability effects: Estimates of General Combining Ability (GCA) effects for each parental inbred line for each trait are illustrated in Table 3. High positive GCA values would be of interest in all studied traits except days to 50% anthesis and silking where high negative values would be useful from the breeder’s point of view. The parental lines P2, P5 and P6 seemed to be the best combiners for grain yield per plant (17.59, 17.97 and 26.42 under normal and 13.36, 17.09 and 11.51 under late sowing dates, respectively) and most of the studied traits under normal and late planting dates. The inbred line P8 is proposed to be the best combiner for all studied traits under both conditions except No. of kernel per row and grain yield per plant under normal sowing date and days to 50% silking and No. of kernel per row under late sowing dates. Indicating that these inbred lines could be considered as good combiners for developing early maturity and high yielding genotypes.
The inbred lines P1, P3 and P4 seemed to be poor combiners for all studied traits under normal and late planting dates except number of kernel per row.
It is worth noting that the inbred line which possessed high GCA effects for grain yield per plant might show the same for one or more of the traits contributing grain yield. In most traits, the values of GCA effects was mostly differed from sowing date to another. This finding coincides with that reached above where significant GCA by planting date mean squares were detected (Table 1).
Specific combining ability effects: Specific combining ability effects for all studied traits under normal and late planting dates are presented in Table 4.
| Table 3: | Estimates of general combining ability effects for eight maize inbred lines evaluated under normal (D1) and late (D2) sowing dates |
 | |
| *,**Significance at 0.05 and 0.01 probability levels, respectively | |
Twenty one out of the twenty eight crosses showed desirable significant specific combining ability effects under normal and late sowing dates for grain yield per plant and most of the other studied traits. Two hybrids (P5xP8 and P6xP8) under late sowing date exhibited desirable significant specific combining ability effects for grain yield per plant and most of the other studied traits. All these crosses are considered to be promising in breeding programs either towards hybrid maize production or synthetic varieties composed of hybrids which involved the good combiners for the traits in view. The remaining crosses (P1xP3, P1xP4 and P3xP4) under both conditions, P5xP8 and P6xP8 under normal sowing date and P4xP7 under late planting date) exhibited insignificant specific combining ability effects for grain yield per plant and some of the other studied traits (Table 4).
Protein electrophoresis: The electrophoretic patterns for water soluble proteins (albumin and globulin) of the eight maize inbred lines are illustrated in Fig. 1 and Table 5. From the SDS PAGE (Sodium dodecyl sulphate-polyacrylamide gel electrophoresis) analysis, fifty bands were observed with different Molecular Weights (MW) and Relative Mobilities (RM). Three universal bands were commonly present in all eight inbred lines of MW 272.2, 246.3 and 58.8 kDa. Substantial differences among the studied inbred lines in their molecular weights and relative mobility were recorded. These parental lines were discriminated from each other by some unique bands, where the parental lines P1, P3 and P4 showed one unique band for each parent at MW 198.9, 144.1 and 140.8 kDa, respectively. Three unique bands characterized the parental lines P2 and P7 at MW 233.3, 152.1 and 111.8 kDa for P2 and 242.8, 138.8 and 114.3 kDa for P7.
| Table 4: | Estimates of specific combining ability effects for twenty eight maize crosses under normal and late planting dates |
 | |
| *,**Significance at 0.05 and 0.01 probability levels, respectively | |
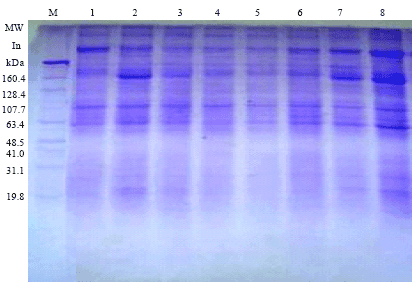 | |
| Fig. 1: | SDS electrophoresis patterns of water soluble protein in eight maize inberd lines |
Two unique bands distinguished the parental lines P5, P6 and P6 at MW 235.3 and 109.2 kDa for P5, 154.3 and 115.3 kDa for P6 and 217.9 and 118 kDa for P8. From these results it is concluded that the analysis of water soluble protein electrophoretic bands could be a useful tool for the identification and characterization of the eight maize inbred lines. These results are in line with the results of Abdel-Tawab et al. (1989), Abdel-Sattar and Ahmed (2004) and El-Shouny et al. (2011).
| Table 5: | Densitometer analysis of water soluble proteins (SDS-PAGE) showing number of bands (B. No.), Relative mobilities (RM) and molecular weight (MW) for eight Maize inbred lines |
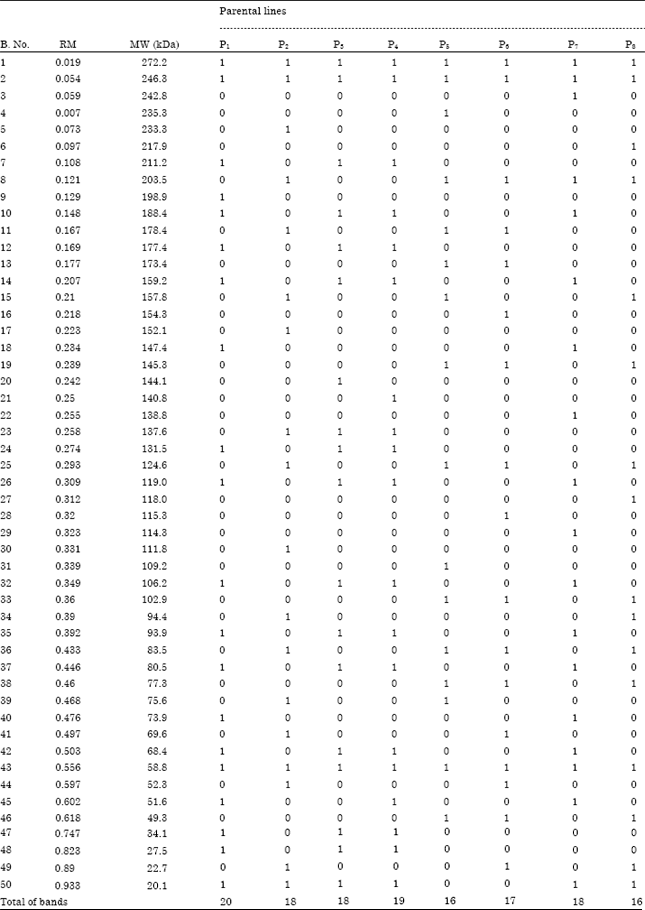 | |
| 1: Present of band, 0: Absent of band | |
In a trail to predict of heterosis and specific (SCA) combining ability via protein electrophoresis as genetic diversity among the eight maize inbred lines. The ten crosses (P1xP2, P1xP5, P1xP6, P1xP8, P2xP5, P2xP6, P3xP6, P5xP6, P5xP7 and P6xP7) exhibited positive significant SCA effects and useful heterosis relative to the commercial variety S.C.10 (Table 2, 4) and in the same time showed wide of genetic variation between the parents of these crosses through protein electrophoretic bands. For example, the inbred line P1 exhibit twenty bands with different MW and Rm from the inbred line P2 (exhibit eighteen bands). These inbred lines (P1 and P2) were only similar at four bands at MW 272.2, 246.3, 58.8 and 20.1 kDa and differed between them in the other bands. The nine crosses (P1xP3, P1xP4, P1xP7, P2xP8, P3xP4, P3xP7, P4xP7, P5xP8 and P6xP8) showed negative significant heterosis and four of them (P1xP3, P1xP4, P3xP4 and P6xP8) showed negative significant SCA effects and the other crosses showed positive significant SCA effects in the same time do not exhibit any genetic diversity between their parents. The parents of these nine crosses were similar in most of their bands. The other nine crosses (P2xP3, P2xP4, P2xP7, P3xP5, P3xP8, P4xP5, P4xP6, P4xP8 and P7xP8) exhibited negative significant heterosis and positive significant SCA effects but in the same time showed wide of genetic variation between their parents through electrophoretic bands (Table 2, 4 and 5).
We could depend on the genetic diversity among maize inbred lines via protein electrophoretic bands in prediction of heterosis and specific combining ability effects because about 67% of these crosses (nineteen out of the twenty eight crosses) valid in prediction of heterosis and combining ability. These results indicated to some extent the effectiveness of using soluble protein electrophoresis in the identification of the highly heterotic hybrids and high specific combining ability as biochemical genetic markers associated with hybrid vigor and specific combining ability in maize crosses. Some studies detected positive association between parental genetic distance based on protein electrophoretic bands and hybrid field performance (Abdel-Tawab et al., 1989; Esmail et al., 1999; Abdel-Sattar and Ahmed, 2004; El-Shouny et al., 2011).
CONCLUSIONS
Results of this study indicated that the three maize parental lines P2, P5 and P6 could be considered as good combiners for developing early maturity and high yielding genotypes under different sowing dates. Nine hybrids, P1xP2, P1xP6, P1xP8, P2xP5, P2xP6, P3xP6, P5xP6, P5xP7 and P6xP7, under normal sowing date and four hybrids, P2xP4, P2xP5, P4xP6 and P5xP6, under late sowing date offer possibility for improving grain yield in maize. The electrophoresis patterns could be useful tools for the identification and characterization of maize inbred lines and could be used as biochemical genetic markers associated with hybrid vigor and SCA in maize.
REFERENCES
- Aliu, S., S. Fetahu and A. Salillari, 2008. Estimation of heterosis and combining ability in maize (Zea mays L.) for ear weight using the diallel cross method. Latvian J. Agron., 11: 7-12.
Direct Link - Beiragi, M.A., S.K. Khorasani, S.H. Shojaei, M. Dadresan, K. Mostafavi and M. Golbashy, 2011. A study on effects of planting dates on growth and yield of 18 corn hybrids (Zea mays L.). Am. J. Exp. Agric., 1: 110-120.
Direct Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lauer, J.G., P.R. Carter, T.M. Wood, G. Diezel, D.W. Wiersma, R.E. Rand and M.J. Mlynarek, 1999. Corn hybrid response to planting date in the northern corn belt. Agron. J., 91: 834-839.
CrossRefDirect Link - Nielson, R.L., P.R. Thomison, G.A. Brown, A.L. Halter, J. Wells and K.L. Wuethrich, 2002. Delayed planting date effects on flowering and grain maturation of corn. Agron. J., 94: 549-558.
Direct Link - Sedhom, S.A., 1994. Estimation of general and specific combining ability In maize under two different planting dates. Ann. Agric. Sci. Moshtohor., 32: 119-130.
Direct Link - Soliman, M.S.M., F.A.E. Nofal and M.E.M.A. El-Azeem, 2005. Combining ability for yield and other attributes in diallel cross of some yellow maize inbred lines. Minufia J. Agric. Res., 30: 1767-1781.
Direct Link - Studier, F.W., 1973. Analysis of bacteriophage T7 early RNAs and proteins on slab gels. J. Mol. Biol., 79: 237-248.
CrossRefPubMedDirect Link - Wynne, J.C., D.A. Emery and P.W. Rice, 1970. Combining ability estimates in Arachis hypogaea L. II. Field performance of F1 hybrids. Crop Sci., 10: 713-715.
CrossRefDirect Link