
Guang Zhu
Department of Gastroenterology, Affiliated Hospital of Yangzhou University, Yangzhou University, 225000, Yangzhou, China
Hai-bo Wang
Department of Gastroenterology, Affiliated Hospital of Yangzhou University, Yangzhou University, 225000, Yangzhou, China
Feng Jin
Department of Gastroenterology, Affiliated Hospital of Yangzhou University, Yangzhou University, 225000, Yangzhou, China
Li-de Tao
Yangzhou University-Yangzhou Cancer Research Institute, Yangzhou University, Yangzhou, 225009, Jiangsu, People�s Republic of China
Dan Li
Department of Gastroenterology, Affiliated Hospital of Yangzhou University, Yangzhou University, 225000, Yangzhou, China
Teng-yang Ni
Department of Gastroenterology, Affiliated Hospital of Yangzhou University, Yangzhou University, 225000, Yangzhou, China
Wen-yuan Li
Department of Gastroenterology, Affiliated Hospital of Yangzhou University, Yangzhou University, 225000, Yangzhou, China
Bo Pan
Yangzhou University-Yangzhou Cancer Research Institute, Yangzhou University, Yangzhou, 225009, Jiangsu, People�s Republic of China
Wei-ming Xiao
Department of Gastroenterology, Affiliated Hospital of Yangzhou University, Yangzhou University, 225000, Yangzhou, China
Yan-bing Ding
Department of Gastroenterology, Affiliated Hospital of Yangzhou University, Yangzhou University, 225000, Yangzhou, China
Masataka Sunagawa
Department of Physiology, School of Medicine, Showa University, 142 Tokyo, Japan
Yan-qing Liu
Department of Gastroenterology, Affiliated Hospital of Yangzhou University, Yangzhou University, 225000, Yangzhou, China
LiveDNA: 86.23350
International Journal of Pharmacology
Year: 2019 | Volume: 15 | Issue: 1 | Page No.: 40-49







ABSTRACT
Background and Objective: The traditional Chinese medicine herb Celastrus Orbiculatus is a folk medicinal plant that has been used in various diseases. The ethyl acetate extract of Celastrus orbiculatus (COE) was reported to show significant anti-tumor effects. Prohibitin (PHB), an evolutionarily-conserved protein has been found to be over-expressed in Gastric Cancer (GC) and be closely related with tumor malignancy. In this study the effects of COE and small interfering RNA (siRNA)-mediated silencing of PHB gene expression on the migration and invasion of GC was investigated. Materials and Methods: The effects of COE on the expression of PHB in MGC-803 GC cells were evaluated by Western Blots. Small interfering RNA (siRNA) sequences targeting PHB were transfected into the MGC-803 cell line. The effect of COE on PHB silenced GC cell viability was examined using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The effects of COE and siRNA-mediated silencing of PHB gene expression on invasion and migration of GC and protein expression of tumor cells were analyzed by wound-healing assay, trans well assay and Western Blots, respectively. Results: It was found that COE reduced PHB expression in MGC-803 cells. In addition, invasion and migration of GC was significantly decreased upon PHB silencing. Gene silencing of PHB increased E-cadherin expression, decreased N-cadherin, Vimentin expression and inhibited c-Raf/ERK signaling pathway. Moreover, COE effectively inhibited GC invasion and migration, EMT and c-Raf/ERK signaling pathway of PHB silenced MGC-803 cells in a dose-dependent manner. Conclusion: The COE inhibits migration and invasion probably via suppressing PHB expression, which correlates with c-Raf/ERK signal pathway inhibition in MGC-803 cells.
PDF Abstract XML References Citation
Received: February 11, 2018;
Accepted: August 06, 2018;
Published: December 15, 2018
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Guang Zhu, Hai-bo Wang, Feng Jin, Li-de Tao, Dan Li, Teng-yang Ni, Wen-yuan Li, Bo Pan, Wei-ming Xiao, Yan-bing Ding, Masataka Sunagawa and Yan-qing Liu, 2019. Celastrus Orbiculatus Extract Suppresses Migration and Invasion
of Gastric Cancer by Inhibiting Prohibitin and c-Raf/ERK Signaling
Pathway. International Journal of Pharmacology, 15: 40-49.
DOI: 10.3923/ijp.2019.40.49
URL: https://scialert.net/abstract/?doi=ijp.2019.40.49
DOI: 10.3923/ijp.2019.40.49
URL: https://scialert.net/abstract/?doi=ijp.2019.40.49
INTRODUCTION
Gastric Cancer (GC) is one of the most common malignancies and currently the second leading cause of cancer-related death worldwide1. The high mortality of GC is related to the late diagnosis owing to non-specific manifestations of the disease in the early phases, as well as to its aggressive nature2. A key event in promoting stationary tumor cells to migrate and invade is the epithelial-mesenchymal transition (EMT) program3. The EMT is a process in which epithelial cells lose their junctions and polarity to gain a motile mesenchymal phenotype, which can be seen in cancer cells as they leave the primary tumor and disseminate to other parts of the body to colonize distant organs and form metastases4. Prohibitin (PHB) is evolutionarily highly conserved and generally existed in eukaryotes5. The PHB protein was initially found in the mitochondrial inner membrane and plays a central role in maintaining mitochondrial morphology and normal functions, thus preventing apoptosis in malignant cells against metabolic stress6. The PHB was found to be overexpressed in several tumor types and plays crucial roles in cancer development and progression7-9. The PHB might be a potential target for the treatment of malignant tumor growth and metastasis10-13.
Celastrus orbiculatus, a member of the Celastraceae family and the genus Celastrus, is an important folk medicine in China for the treatment of many diseases. Preliminary research indicates that the ethyl acetate extract of Celastrus orbiculatus (COE) possesses anti-tumor effects in vitro and in vivo14-16. However, the underlying molecular mechanisms of COE-induced inhibition of GC invasion and migration has not been elucidated so far. Thus, the aim of the present study was to investigate the effect of COE on the metastasis and invasion of GC through PHB and c-Raf/ERK pathway, which may have significant implications for the development of novel anti-tumor traditional Chinese medicine.
MATERIALS AND METHODS
Drug extraction: Celastrus orbiculatus (Batch No: 070510) were purchased from Guangzhou Zhixin Pharmaceutical Co. Ltd. (Guangzhou, China). The COE was characterized and prepared at the Department of Chinese Materia Medica Analysis, China Pharmaceutical University (Nanjing, China). The detailed preparation procedure of COE has been described previously17,18. Briefly, the dried stem of Celastrus orbiculatus was crushed into powder and extracted three times using 95% ethanol. After filtration, ethanol removal and vacuum freeze-drying at low temperature, the extract was obtained. Finally, the ethyl acetate extract of the Celastrus orbiculatus was condensed, purified and lyophilized into powder and stored at 4°C. The COE powder was dissolved in dimethyl sulfoxide (DMSO) and then diluted to different concentrations with serum-free medium prior to being used. The final concentration of DMSO in the treatment medium did not exceed 0.1%.
Chemical reagents and antibodies: RPMI-1640 medium and Fetal Bovine Serum (FBS) were purchased from Gibco; Thermo Fisher Scientific, Inc. (Waltham, MA, USA). MTT and DMSO were purchased from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). Matrigel was purchased from BD Biosciences (San Jose, CA, USA). Antibodies against E-cadherin (cat # 3195), N-cadherin (cat # 4061), vimentin (cat # 5741), MMP-2 (cat # 4022), MMP-9 (cat # 13667), Prohibitin (cat # 2426), c-Raf (cat # 9422), ERK(cat # 4695), p-ERK (cat # 4370) and β-actin (cat # 3700) were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). Other reagents used of analytical grade were obtained from commercial sources.
Cell culture: The human gastric cancer cell line MGC-803 was obtained from the Cell Bank of Chinese Academy of Sciences, Shanghai Institute of Cell Biology (Shanghai, China). MGC-803 cells were cultured with RPMI-1640 medium containing 10% FBS at 37°C in a humidified 5% CO2 incubator. Cell morphology was observed at ×100 magnification in an optical microscope (IX72; Olympus Corporation, Tokyo, Japan).
RNA interference: Recombinant lentiviruses expressing LV-PHB-RNAi or negative control LV-NC-RNAi were produced by Genechem (Shanghai, China). The MGC-803 cells were infected with concentrated virus according to the manufacturer’s instructions. Cells were then grown for 48 h at 37°C, 5% CO2. The cells were visualized under a fluorescence microscope (Nikon, Chiyoda-Ku, Tokyo, Japan) to detect transfection efficiency and the PHB expression in the infected cells was validated by Western Blotting assay.
Cell viability assay: Cell viability was analyzed using the MTT assay. MGC-803 cells were grown until logarithmic phase after infection and then plated in a 96-well plate at a density of 1,000-10,000 cells/well at 37°C for 12 h. After attachment, cells were randomly grouped into the control group and COE groups. Cells in COE groups were incubated with different concentrations of COE ranging between 10 and 320 μg mL–1, with five wells for each concentration. After 24, 48 and 72 h of incubation, the cells were incubated with RPMI-1640 medium containing 0.5 mg mL–1 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) for 4 h at 37°C, followed by dissolving the formazan crystals with 150 μL DMSO. The plates were incubated on an agitator for 15 min at room temperature and the absorbance (A) value was measured at 490 nm by a microplate reader. The inhibition rate of COE was calculated as:
![]()
Cell migration and invasion assays: In the wound-healing assay, cells were seeded into 6 well plates and grown to confluence reached 90%. Wounds were made by scraping confluent cell monolayers with micropipette tips, then washed off the exfoliated cells three times with PBS. Photomicrographs were taken at time points 0 and 24 h after wounding. The percentage (%) change of wound healing was determined via comparison of the differences in wound width. Moreover, cell migration and invasion were evaluated using 8 μm transwell filters (Corning Incorporated, Corning, NY, USA). Briefly, cells were suspended in 200 μL serum-free media and then added into the upper chamber with an uncoated or Matrigel-coated membrane of the transwell, whereas 600 μL culture medium containing 10% FBS was added into the lower chamber. Cells in the COE group were treated with 10, 20 or 40 μg mL–1 COE, with three wells for each concentration. After incubation for 24 h, the cells that migrated or invaded through and adhered to the bottom of the membrane were fixed with 95% methanol for 30 min and stained with 0.1% crystal violet for 20 min at room temperature. Five fields (200× magnification) were randomly captured for each membrane under a microscope (Nikon Corporation, Tokyo, Japan) and the migratory and invasive cells were counted and averaged. Each experiment was repeated 3 times.
Western blot analysis: The expression levels of PHB, EMT-related proteins (E-cadherin, N-cadherin, vimentin), MMPs, c-Raf, ERK and p-ERK were determined by western blot analysis. β-actin was used as the loading control. Briefly, total cellular protein of cells was extracted after cell lysis by Cytoplasmic Extraction Reagent II (Fermentas Thermo Fisher Scientific, Inc., Pittsburgh, PA, USA). Lysate proteins were resolved by 10% SDS-PAGE and then blotted onto the nitrocellulose membranes (Millipore, Bedford, MA, USA). The membranes were blocked with 5% skim milk kept for 2 h at room temperature and incubated overnight at 4°C with appropriate primary antibodies. Afterwards, the secondary antibody was added after washing three times and incubated at room temperature for 2 h, followed by enhanced chemiluminescence (ECL) detection using the Super Signal West Pico Chemiluminescent Substrate (Thermo Fisher Scientific, Inc.) on a Molecular Imager ChemiDoc XRS System (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis: Each experiment was repeated >3 times. Data processing was performed by SPSS 16.0 software (SPSS Inc., Chicago, IL) using one-way analysis of variance. The data are expressed as the Mean±standard deviation and statistical significance was accepted when p<0.05.
RESULTS
Effects of COE on expression of PHB in MGC-803 cells: A previous study has identified that COE inhibited proliferation of MGC-803 cells and the IC50 for MGC-803 cells was 108.9 μg mL–1 19. Therefore, COE concentrations of 20, 40 and 80 μg mL–1 were used in this experiment to exclude the cytotoxic effect of COE. MGC-803 cells treated with COE for 24 h showed decreased expression of PHB in a dose-dependent manner (Fig. 1).
Gene silencing of PHB in MGC-803 cells: Recombinant lentiviruses expressing LV-PHB-RNAi or negative control LV-NC-RNAi were transfected into MGC-803 cells and the cells were selected by flow cytometry for GFP+cells, a transduction efficiency of close to 100% was achieved (Fig. 2a). Western Blot analysis indicated that PHB protein expression in LV-PHB-RNAi group was markedly silenced (p<0.01) compared with the normal MGC-803 cells (NC group) and LV-NC-RNAi group (Fig. 2b, c).
Effects of PHB silencing on invasion and migration of MGC-803 cells: The wound-healing assay indicated that PHB RNA interference for 48 h significantly inhibited the migration of MGC-803 cells (Fig. 3a, b). Moreover, the number of cells in LV-PHB-RNAi group penetrating the membrane was significantly reduced in comparison to the NC and LV-NC-RNAi group (Fig. 3c, d). The results indicated that PHB silencing inhibited cell migration and invasion of MGC-803 cells.
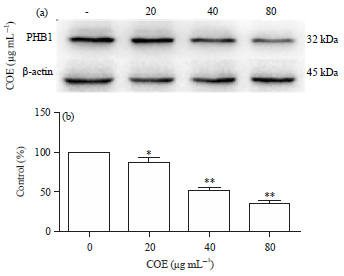 | |
| Fig. 1(a-b): | COE treatment MGC-803 cells for 24 h altered the expression of PHB1 in a dose-dependent manner, (a) PHB1 expression as shown using western blots and (b) Quantification data |
| *p<0.05, **p<0.01, as compared with the untreated control | |
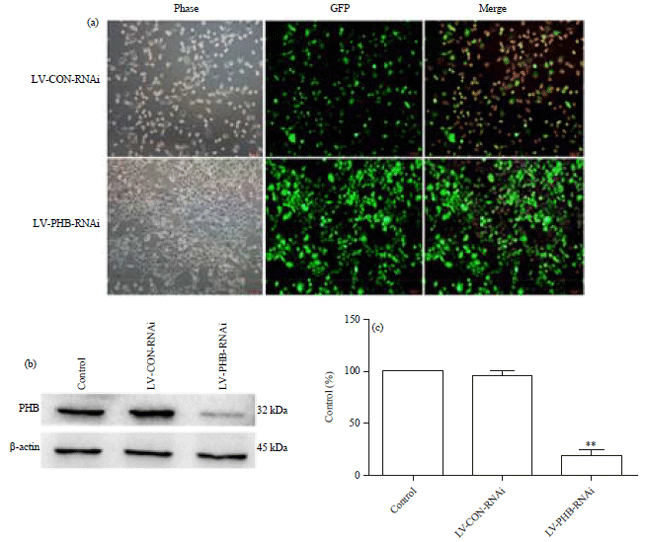 | |
| Fig. 2(a-c): | LV-PHB-RNAi silenced PHB expression (100×), (a) Expression of green fluorescent protein before and after transfection with lentivirus and (b, c) LV-PHB-RNAi clearly down-regulated PHB expression in MGC-803 cells |
| **p<0.01 | |
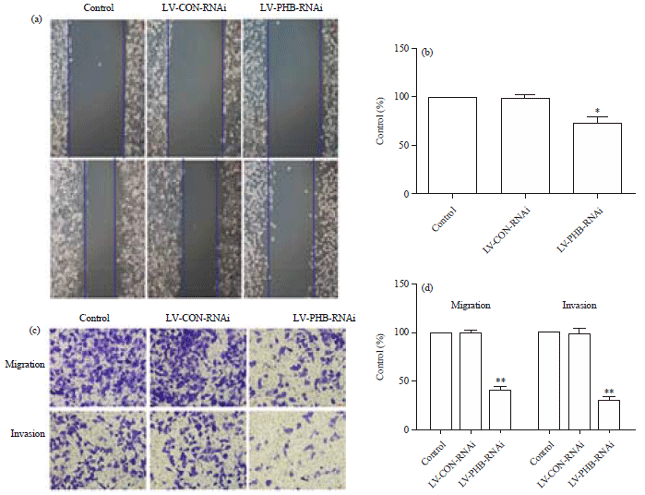 | |
| Fig. 3(a-d): | PHB silencing inhibited the invasion and migration of MGC-803 cells in a dose-dependent manner. Cell invasion and migration were assessed after 24 h incubation by wound-healing assay and transwell assay, (a, c) Cresyl violet staining (200× magnification) and (b, d) Quantification data |
| Values are expressed as Means±SD of three independent experiments, *p<0.05, **p<0.01, as compared with the untreated control | |
Effects of PHB silencing on expression of EMT-related proteins and c-Raf/ERK pathway: Compared with the control groups, the LV-PHB-RNAi group displayed a decrease in the protein expression of Vimentin, N-cadherin, MMP-2 and MMP-9. However, increased expression of E-cadherin was observed (Fig. 4a, b). Furthermore, PHB silencing significantly reduced the levels of c-Raf, ERK and p-ERK (Fig. 4c, d).
Effects of COE on viability of MGC-803/PHB– cells: Control MGC-803/PHB– cells exhibited active growth in vitro, whereas cells treated with various concentrations of COE for 24, 48 or 72 h showed significantly inhibited growth (*p<0.05, **p<0.01) (Fig. 5). The IC50 for MGC-803/PHB– cells was 83.6 μg mL–1. Therefore, the COE concentrations of 10, 20 and 40 μg mL–1 were used in subsequent experiments to exclude the cytotoxic effect of COE on cell invasion and migration.
Effects of COE on invasion and migration of MGC-803/PHB– cells: In the wound-healing assay, COE treatment for 24 h significantly inhibited the migration of MGC-803/PHB– cells in a dose-dependent manner (Fig. 6a, b). Furthermore, in the transwell migration and invasion assay, COE treatment significantly and dose-dependently decreased the number of transmembrane cells (Fig. 6c, d), indicating that COE inhibited cell migration and invasion of MGC-803/PHB– cells.
Effects of COE on expression of EMT-related proteins and c-Raf/ERK pathway: MGC-803/PHB– cells treated with COE for 24 h showed increased E-cadherin expression and decreased the expression of N-cadherin, vimentin, MMP-2 and MMP-9 in a dose-dependent manner (Fig. 7a, b).
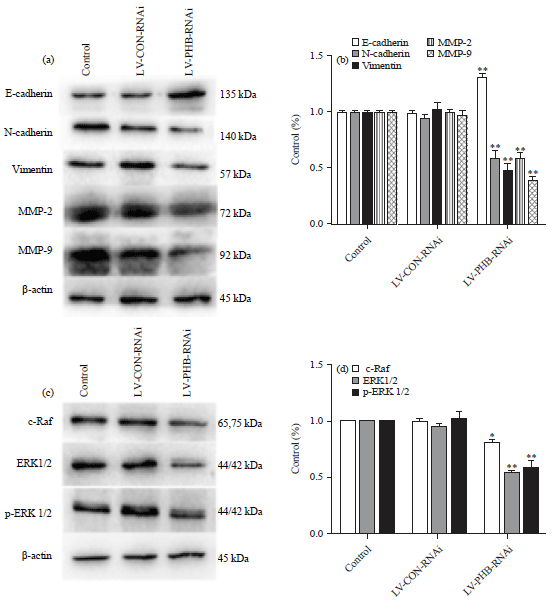 | |
| Fig. 4(a-d): | PHB silencing altered the expression of EMT-related proteins, enzymes, c-Raf/ERK pathway in a dose-dependent manner, (a, c) Protein expression as shown using western blots and (b, d) Quantification data |
| *p<0.05, **p<0.01, as compared with the untreated control | |
Furthermore, COE treatment dose-dependently decreased c-Raf, ERK and p-ERK expression (Fig. 7c, d).
DISCUSSION
In the present study, Western Blot analysis revealed that treatment with COE for 24 h down-regulated the expression level of PHB in a dose-dependent manner. RNA interference were used to produce PHB gene silencing in MGC-803 GC cells. After PHB was silenced by LV-PHB-RNAi, the invasion and migration of MGC-803 cells were inhibited in various migration and invasion assays. Moreover, the MTT assay was used to assess cell viability and MGC-803/PHB– cells showed decreased growth depending on the concentration and duration of COE treatment. When MGC-803/PHB– cells were treated with COE under low-cytotoxic concentrations, the invasion and migration and EMT process were inhibited and COE also had a significant effect on the blockage of c-Raf/ERK pathway. These results indicated that the mechanism of COE to inhibit the invasion and migration of GC may involve the suppression of PHB/c-Raf/ERK signal pathway and PHB may be one of the potential drug targets for gastric cancer therapies.
The COE is a new preparation extracted from the Chinese medicinal herb Celastrus orbiculatus. Previous studies have confirmed that COE inhibits cell proliferation, induces cell apoptosis, suppresses tumor angiogenesis and inhibits tumor metastasis14-16,19-21.
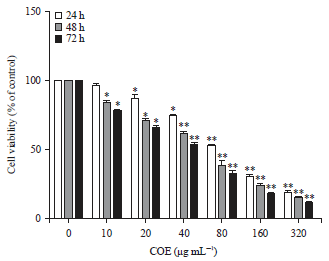 | |
| Fig. 5: | COE treatment inhibited the viability of MGC-803/PHB– cells by the MTT assay. Cells were incubated with the indicated concentration of COE for 24 h. Dose and time-dependent curve of inhibition rate of COE on MGC-803/PHB– cells by the MTT assay |
| Date were presented as the Means±SD of three independent experiments performed in quintuplicate. *p<0.05, **p<0.01, as compared with the untreated control | |
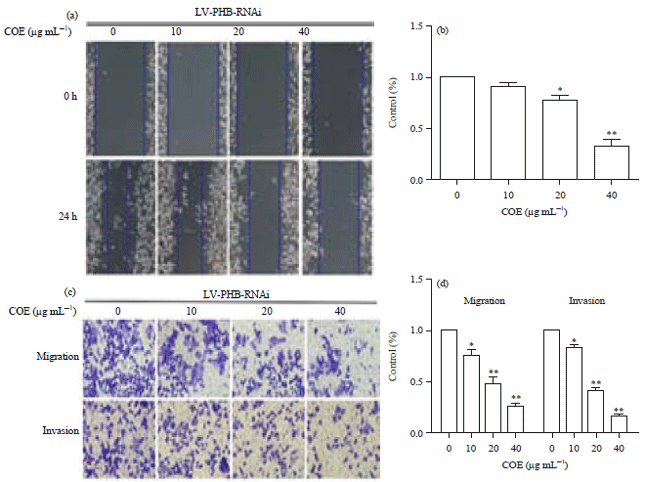 | |
| Fig. 6(a-d): | COE treatment for 24 h inhibited the invasion and migration of MGC-803 cells in a dose-dependent manner. Cell invasion and migration were assessed after 24 h incubation by wound-healing assay and transwell assay, (a, c) Cresyl violet staining (200×magnification) and (b, d) Quantification data |
| Values are expressed as Means±SD of three independent experiments. *p<0.05, **p<0.01, as compared with the untreated control | |
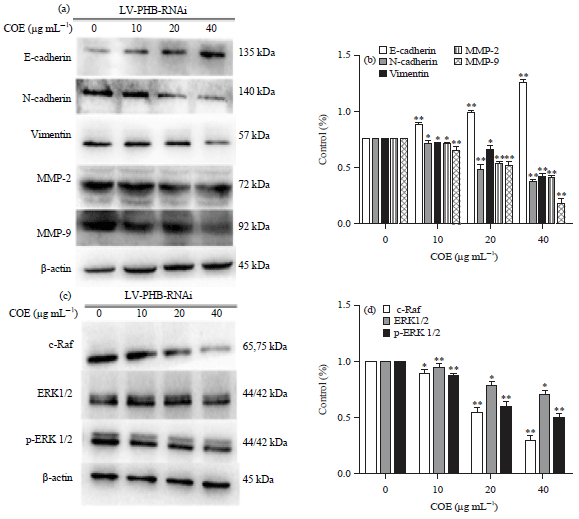 | |
| Fig. 7(a-d): | COE treatment MGC-803/PHB– cells for 24 h altered the expression of EMT-related proteins, enzymes, c-Raf/ERK pathway in a dose-dependent manner, (a, c) Protein expression as shown using western blots and (b, d) Quantification data |
| *p<0.05, **p<0.01, as compared with the untreated control | |
Proteomic expression profiling was applied for the potential molecular targets of COE inhibits TGF-β1 mediated EMT and PHB was identified as one of the most dramatically altered protein22. Therefore, PHB was selected for further study. Although, several reports have suggested that the function of COE are related with tumor metastasis but the specific molecular mechanism remains unclear. In the present study, it demonstrated that COE had a satisfactory anti-tumor activity through inhibition of PHB and c-Raf/ERK signal pathway.
Malignant tumor cells reactivate EMT to acquire the ability to migrate and invade as single cells3. EMT is a developmental program that enables stationary epithelial cells to undergo from phenotypic conversion to mesenchymal cells4. The loss of E-cadherin and the acquisition of N-cadherin, Vimentin are representative characteristics of EMT process23. In addition, Matrix metalloproteinases (MMPs) is a group of zinc-dependent enzymes capable of cleaving components of the extracellular matrix (ECM) and inducing tumor cell proliferation and metastasis24. There into, MMP-2 and MMP-9 are closely correlated with invasion and migration of GC25 and the increased expression of MMP-2 and MMP-9 appears to be a concomitant event in the EMT process of tumor cells26. In the study, Silencing of PHB upregulated the epithelial cell marker (E-cadherin) expression and down-regulated mesenchymal cell marker (N-cadherin, Vimentin) expression. Furthermore, the expression levels of MMP-2 and MMP-9 were decreased after PHB silencing. These data indicated that silencing PHB may suppress cell invasion and migration and inhibit the process of EMT in GC.
PHB is required for Ras-induced Raf-MEK-ERK activation and epithelial cell migration27, supporting the pro-tumorigenic role of PHB in cancer progression. Specifically, the direct interaction of PHB with c-Raf is required for the localization and phosphorylation of c-Raf and results in the activation of Raf and the downstream activation of the ERK pathway10,27. The c-Raf/ERK signaling pathway is a highly conserved signaling module, which regulates multitude of cellular functions such as proliferation, differentiation, cell death, migration, etc. and is involved in the formation of cancer cells distant metastases28. Moreover, this pathway play a role in promotion of mesenchymal protein expression and its downstream effectors induce expression of EMT transcription factors that promote repression of epithelial and induction of mesenchymal character26. In this study, upon depletion of PHB in GC cells, a dramatic reduction in the c-Raf/ERK pathway activation was observed, as reflected by the decreased c-Raf, ERK and p-ERK levels. It seems likely that regulation of PHB on c-Raf/ERK signaling pathway is involved in the EMT process.
CONCLUSION
The present study suggests that COE is a potential inhibitor of PHB in human gastric cancer. COE may inhibit invasion and migration and EMT process by suppressing PHB expression, together with inhibiting c-Raf/ERK signal pathway. These findings provide novel evidence that COE may be a potential anti-cancer therapy all over the world, particularly in the cancer with over-expressed PHB.
SIGNIFICANCE STATEMENT
This study discovered the effect of COE that can inhibit the invasion and migration of GC through the suppression of PHB/c-Raf/ERK signaling pathway. The results may be beneficial for the development of new strategies for the formulation of effective therapeutic drugs that would selectively target cancer cells and improve overall survival in patients with cancer. This study help the researcher to uncover the critical areas of targeted cancer therapy with traditional Chinese medicines and on which future studies in this research area can be premised.
ACKNOWLEDGMENT
This study was supported by grants from the National Natural Science Foundation of China (NO. 81773944 and 81573656) and the Natural Science Foundation of Jiangsu Province of China (NO. BK 20141280).
REFERENCES
- Van Cutsem, E., X. Sagaert, B. Topal, K. Haustermans and H. Prenen, 2016. Gastric cancer. Lancet, 388: 2654-2664.
CrossRefPubMedDirect Link - Macedo, F., K. Ladeira, A. Longatto-Filho and S.F. Martins, 2017. Gastric cancer and angiogenesis: Is VEGF a useful biomarker to assess progression and remission? J. Gast. Cancer, 17: 1-10.
CrossRefDirect Link - Yeung, K.T. and J. Yang, 2017. Epithelial-mesenchymal transition in tumor metastasis. Mol. Oncol., 11: 28-39.
CrossRefDirect Link - Nieto, M.A., 2013. Epithelial plasticity: A common theme in embryonic and cancer cells. Science, Vol. 342.
CrossRefDirect Link - Lv, Z., X. Zhang, L. Liu, J. Chen and Z. Nie et al., 2012. Characterization of a gene encoding prohibitin in silkworm, Bombyx mori. Gene, 502: 118-124.
CrossRefDirect Link - Koushyar, S., W.G. Jiang and D.A. Dart, 2015. Unveiling the potential of prohibitin in cancer. Cancer Lett., 369: 315-322.
CrossRefDirect Link - Ko, K.S., M.L. Tomasi, A. Iglesias‐Ara, B.A. French and S.W. French et al., 2010. Liver-specific deletion of prohibitin 1 results in spontaneous liver injury, fibrosis and hepatocellular carcinoma in mice. Hepatology, 52: 2096-2108.
CrossRefDirect Link - Guo, F., K. Hiroshima, D. Wu, M. Satoh and M. Abulazi et al., 2012. Prohibitin in squamous cell carcinoma of the lung: Its expression and possible clinical significance. Hum. Pathol., 43: 1282-1288.
CrossRefDirect Link - Cheng, J., F. Gao, X. Chen, J. Wu and C. Xing et al., 2014. Prohibitin-2 promotes hepatocellular carcinoma malignancy progression in hypoxia based on a label-free quantitative proteomics strategy. Mol. Carcinog., 53: 820-832.
CrossRefDirect Link - Luan, Z., Y. He, M. Alattar, Z. Chen and F. He, 2014. Targeting the prohibitin scaffold-CRAF kinase interaction in RAS-ERK-driven pancreatic ductal adenocarcinoma. Mol. Cancer, Vol. 13.
CrossRefDirect Link - Kang, X., L. Zhang, J. Sun, Z. Ni and Y. Ma et al., 2008. Prohibitin: A potential biomarker for tissue-based detection of gastric cancer. J. Gastroenterol., 43: 618-625.
CrossRefDirect Link - Cao, Y., H. Liang, F. Zhang, Z. Luan and S. Zhao et al., 2016. Prohibitin overexpression predicts poor prognosis and promotes cell proliferation and invasion through ERK pathway activation in gallbladder cancer. J. Exp. Clin. Cancer Res., Vol. 35.
CrossRefDirect Link - Canevari, R.A., F.A. Marchi, M.A. Domingues, V.P. de Andrade and J.R. Caldeira et al., 2016. Identification of novel biomarkers associated with poor patient outcomes in invasive breast carcinoma. Tumor Biol., 37: 13855-13870.
CrossRefDirect Link - Qian, Y.Y., H. Zhang, Y. Hou, L. Yuan and G.Q. Li et al., 2012. Celastrus orbiculatus extract inhibits tumor angiogenesis by targeting vascular endothelial growth factor signaling pathway and shows potent antitumor activity in hepatocarcinomas in vitro and in vivo. Chinese J. Integr. Med., 18: 752-760.
CrossRefDirect Link - Ma, H., Y. Qian, H. Zhang, X. Ji, Y. Zhu, P. Cui and Y. Liu, 2013. Celastrus orbiculatus extract could inhibit human colorectal carcinoma HT-29 cells metastasis via suppression of the mTOR signaling pathway. Life Sci. J., 10: 1704-1710.
Direct Link - Zhu, Y., Y. Liu, Y. Qian, X. Dai and L. Yang et al., 2015. Antimetastatic effects of Celastrus orbiculatus on human gastric adenocarcinoma by inhibiting epithelial–mesenchymal transition and NF-κB/Snail signaling pathway. Integr. Cancer Ther., 14: 271-281.
Direct Link - Zan, K., X.Q. Chen, Q. Wang and L. Cao, 2007. Chemical constituents in stem of Celastrus orbiculatus. Chinese Trad. Herbal Drugs, Vol. 38.
Direct Link - Li, J.J., J. Yang, F. Lü, Y.T. Qi, Y.Q. Liu, Y. Sun and Q. Wang, 2102. Chemical constituents from the stems of Celastrus orbiculatus. Chinese J. Natural Med., 10: 279-283.
CrossRefDirect Link - Zhu, Y.D., Y.Q. Liu, Y.Y. Qian, H. Zhang, G.Q. Li and L. Yang, 2014. Extracts of Celastrus orbiculatus exhibit anti-proliferative and anti-invasive effects on human gastric adenocarcinoma cells. Chinese J. Integr. Med.
CrossRefDirect Link - Yang, L., Y. Liu, M. Wang, Y. Qian and X. Dai et al., 2016. Celastrus orbiculatus extract triggers apoptosis and autophagy via PI3K/Akt/mTOR inhibition in human colorectal cancer cells. Oncol. Lett., 12: 3771-3778.
CrossRefDirect Link - Zhang, H., Y. Qian, Y. Liu, G. Li and P. Cui et al., 2012. Celastrus orbiculatus extract induces mitochondrial-mediated apoptosis in human hepatocellular carcinoma cells. J. Trad. Chinese Med., 32: 621-626.
CrossRefDirect Link - Zhu, Y., Y. Liu, Y. Qian, X. Dai and L. Yang et al., 2014. Research on the efficacy of Celastrus orbiculatus in suppressing TGF-β1-induced epithelial-mesenchymal transition by inhibiting HSP27 and TNF-α-induced NF-κB/Snail signaling pathway in human gastric adenocarcinoma. BMC Complement. Alternat. Med., Vol. 14.
CrossRefDirect Link - Wang, H., L. Tao, F. Jin, H. Gu and X. Dai et al., 2017. Cofilin 1 induces the epithelial-mesenchymal transition of gastric cancer cells by promoting cytoskeletal rearrangement. Oncotarget, 8: 3919-3942.
Direct Link - Shay, G., C.C. Lynch and B. Fingleton, 2015. Moving targets: Emerging roles for MMPs in cancer progression and metastasis. Matrix Biol., 44: 200-206.
CrossRefDirect Link - Hua, H., M. Li, T. Luo, Y. Yin and Y. Jiang, 2011. Matrix metalloproteinases in tumorigenesis: An evolving paradigm. Cell. Mol. Life Sci., 68: 3853-3868.
CrossRefDirect Link - Pearlman, R.L., M.K.M. de Oca, H.C. Pal and F. Afaq, 2017. Potential therapeutic targets of epithelial-mesenchymal transition in melanoma. Cancer Lett., 391: 125-140.
CrossRefDirect Link - Rajalingam, K., C. Wunder, V. Brinkmann, Y. Churin and M. Hekman et al., 2005. Prohibitin is required for Ras-induced Raf-MEK-ERK activation and epithelial cell migration. Nature Cell Biol., 7: 837-843.
CrossRefDirect Link - Pachmayr, E., C. Treese and U. Stein, 2017. Underlying mechanisms for distant metastasis-molecular biology. Visceral Med., 33: 11-20.
CrossRefDirect Link