
Muhammad Irshad
Department of Biochemistry and Biotechnology, University of Gujrat, Pakistan
Muhammad Asgher
Department of Biochemistry, University of Agriculture Faisalabad, Pakistan
Khizar Hayat Bhatti
Department of Botany, University of Gujrat, Pakistan
Muddassar Zafar
Department of Biochemistry and Biotechnology, University of Gujrat, Pakistan
Zahid Anwar
Department of Biochemistry and Biotechnology, University of Gujrat, Pakistan
International Journal of Pharmacology
Year: 2017 | Volume: 13 | Issue: 7 | Page No.: 808-817







ABSTRACT
In the past few years, both phytase and phytate have gained a special place in the biotechnological sector at large and biomedical sector in particular. Phytase belongs to a group of novel organic phosphorus-containing compounds which are widely distributed in nature. The scientific literature is witnessed of a significant landmark in phytase research with the discovery that not all phytases share identical catalytic mechanisms. In this context, many research scientists or research based organizations, around the globe, have already been or being directed or redirecting their research interests on designing and engineering ideal phytase-based bio-products for targeted applications in various sectors of the modern world. Many efforts have been made on the biotechnological valorization of this potent enzyme. However, the current literature still lacks with its biomedical valorization. Though, the information on medical perspectives of phytase and phytate is very scarce and scattered. Therefore, herein an effort has been made to discuss the anticancer and nutraceutical potentialities of phytase or phytate at one place. Besides from the anticancer potential, phytase/phytate also have numerous health benefits e.g. the physiological presence of phytate in our body, close association of phytate-rich diet with low incidence of numerous diseases and vice versa.
PDF Abstract XML References Citation
How to cite this article
Muhammad Irshad, Muhammad Asgher, Khizar Hayat Bhatti, Muddassar Zafar and Zahid Anwar, 2017. Anticancer and Nutraceutical Potentialities of Phytase/Phytate. International Journal of Pharmacology, 13: 808-817.
DOI: 10.3923/ijp.2017.808.817
URL: https://scialert.net/abstract/?doi=ijp.2017.808.817
DOI: 10.3923/ijp.2017.808.817
URL: https://scialert.net/abstract/?doi=ijp.2017.808.817
INTRODUCTION
Phytase belongs to a group of novel organic phosphorus-containing compounds which are widely distributed in nature. Historically, the first phytase activity was from rice bran and reported in the literature by Suzuki and co-workers in 19071. Specifically, phytase is a mixture of phytic acid (myo-inositol 1, 2, 3, 4, 5, 6-hexakis dihydrogen phosphate) and mixed cations of phytic acid. In recent years, phytases are of supreme interest as phosphorus in terms of bioavailability features among different sources including plants, animals and microbes2-6 and considered as a vital constituent for growth. Phytase (myo-inositol hexakisphosphate phosphohydrolase) is a generic term used to describe a stepwise removal of inorganic orthophosphates from phytic acid (myo-inositol hexakisphosphate) or its salt phytate7,8. Phytase is a complex of different phytases and has been classified as (1) Three-phytases, EC 3.1.3.8, (2) Six-phytases, EC 3.1.3.26 and (3) Five-phytases EC 3.1.3.72. This specific classification is based on the position of specificity of the initial hydrolysis of phytate. Whereas, based on reaction environment, phytases are widely classified into two major groups i.e., (1) Alkaline phytases and (2) Acidic phytases. From the catalytic point of view, phytases have been categorized into various classes i.e., (1) Cysteine phytases or protein tyrosine phytase (PTPhy), (2) Histidine acid phosphatases (HAPhy), (3) β-propeller phytases (BPPhy) and (4) Purple acid phosphatase (PAPhy)5,7-10.
The past many years are witnessed of a significant landmark in phytase research with the discovery that not all phytases share identical catalytic mechanisms. Among all phytases, the foremost and extensively investigated phytase group belongs to the class HAPhy8. Consequently, a commonly accepted phytase nomenclature has been proposed to outline the corresponding three-dimensional structures and catalytic mechanisms for HAPhy, BPPhy, PAPhy, PTPhy, respectively11,12. Ultimately, the proposed phytase nomenclature further expands the potential applications of phytases subject to the distinct catalytic mechanisms5,6,8. The distribution, physicochemical, structural and catalytic characteristics of various phytases have been reviewed elsewhere5,6,8,13, thus only a concise description is given in this review work. The present review main focuses on an area that has not been comprehensively reviewed so far. The main focus has been given to biomedical applications of phytase and phytate with special emphasis to anticancer and biotechnological roles of phytase and phytate with special reference to nutraceutical. As mentioned earlier, up till now, many comprehensive reviews with special emphasis on nutritional or anti-nutritional impacts of phytase or phytate have been published but the recent literature still lacks the medicinal valorization of this potent source. Since the information on medical perspectives of phytase and phytate is very scarce and scattered, thus an effort has been made to discuss such useful aspects at one place, in this review.
Biomedical and biotechnological applications of phytase/phytate: From the application viewpoint, a wider spectrum of phytase sources including plants, animals and microbes is available and following a careful evaluation and characterization, it offers numerous applications in the biomedical and biotechnological sector of the modern world. During the past years, a considerable improvement in many of the processes related to phytase and phytate has appeared and triggered in-depth studies of enzyme-based novel constructs and their vital position in the modern era of biotechnology. Owing to those mentioned above unique structural, physicochemical and diverse catalytic functionalities, phytase has approved as a potential candidate for a range of biomedical, pharmaceutical and nutraceutical applications. In this context, many scientists, around the globe, have already been or being directed or redirecting their research interests on designing and engineering ideal phytase-based bio-products for targeted applications. Figure 1 illustrates various biomedical applications of phytase. Among them, many biomedical and biotechnological applications of phytase have not been reviewed yet, comprehensively.
Biomedical applications: In the past few years, both phytase and phytate have gained a special place in the biomedical sector. Many monogastric animals suffer from no or low phytase activity level in their gastric tracts, thus cannot digest/mineralize the portion of phytate available in the food5,14. Besides other negative impact/influence on human health, the phytate consumption has some favorable aspects too. For example, the phytate containing vegetarian-based diet has shown a lower incidence of cancer, thus act as an anticarcinogenic15,16. The available dietary phytate also support in lowering the blood glucose response by reducing the rate of starch digestion17 and also has considerable potential to regulate insulin secretion18, thus has been found beneficial for diabetic patients. Also, phytate has considerable potential to regulate/reduces blood clots, lowers cholesterol and triglycerides and thus prevents heart diseases19,20.
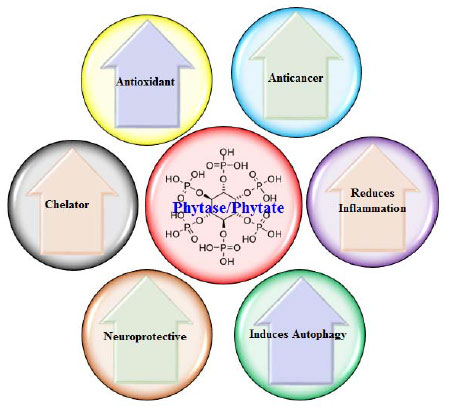 | |
| Fig. 1: | Biomedical and biotechnological applications of phytase/phytate |
Its other biological activities include antioxidant function, reduce/prevents renal stone development, prevent dental cavities and inhibiting the production of hydroxyl radicals, etc21-23. A few decades back, in-vitro studies have indicated that phytic acid incubated with HIV-1 infected T cells inhibits the replication of HIV-124,25.
Anti-cancerous role: The anti-cancerous role of phytate also termed as phytic acid, IP6 or inositol hexaphosphate is considered one of its most significant beneficial activities. Moreover, IP6 (consisting of one inositol and 6 phosphate groups) acts as a broad-spectrum antineoplastic agent and is assimilated by different cells types, including tumor cells16. The antiproliferative potential of IP6 has been demonstrated in various in-vitro studies using various cancer cells. For example, human colon cancer HT-29 cells26, human leukaemic hematopoietic cell lines, such as K-56227, human normal and leukaemic hematopoietic cells27, breast cancer cells28, cervical cancer29, prostate cancer30,31 and HepG2 hepatoma cell lines32 have been treated/inhibited via phytate administration. The action mechanism by which IP6 exerts its anti-cancer activity is not clear yet. One possible exception could be through its antioxidant activity. Besides its direct action on tumor cells, IP6 stimulates the immune system by increasing the activity of natural killer cells. Tumors stimulate the formation of endothelial cells resulting in the formation of blood vessels. These blood vessels provide the necessary supply of nutrients required for tumor growth. Studies have shown that IP6 inhibits the formation of endothelial cells16. Aisde from phytate, phytates have been shown to inhibit the growth of human leukemia cells, colon cancer cells, both estrogen receptor-positive and negative breast cancer cells, voicebox cancer, cervical cancer, prostate cancer, liver tumors, pancreatic, melanoma and muscle cancers. Recently, Soni et al.33 developed a novel phytase enzyme based nanospheres with anticancer potentialities. According to the authors, a controlled self-assembly of phytase enzyme in an Ionic Liquid 1-butyl-3- methylimidazolium tetrafluoroborate led to the formation of therapeutically active phytase nanospheres. The newly developed Pt-coated hybrid bio-macromolecular phytase nanospheres following curcumin loading were subjected to their anticancer efficacy against three different cancer cell lines i.e., MCF-7, Hep-G2 and THP-1 derived human macrophages33. Very recently, Barahuie and co-workers (2017) reported a sustained release of anticancer agent phytic acid from its chitosan-coated magnetic nanoparticles for drug-delivery system. A 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay was used to evaluate the anticancer potential of newly developed phytic acid-chitosan-iron oxide nanocomposite against HT-29 colon cancer cells. Comparative cytotoxic evaluation was also performed with no cytotoxic effect on normal cells (3T3 fibroblast cells)34.
Colon cancer or colorectal cancer (CRC): Colon cancer, a major neoplastic disease, also known as bowel cancer and colorectal cancer (CRC). The CRC is considered one among the major morbidity and mortality issues in western countries. According to a recent report from the American Cancer Society in 2017, 95,520 new cases were registered with colon cancer only, Whereas, up to 50, 260 death incidents occurred collectively because of the colon and rectal cancers in the United States (ACS35). Apart from the very obvious risk factor i.e., increasing age, likewise other most cancers, following risk factor majorly contributes in CRC cancer:
| • | The family history of CRC (especially in the first-degree relative)36 |
| • | The personal history of CRC37-39 |
| • | Hereditary conditions (hereditary nonpolyposis colorectal cancer [HNPCC])40 |
| • | Overdose alcohol consumption41 |
| • | Tobacco smoking42 |
| • | Obesity43 |
| • | Race/ethnicity: American, African, etc44,45 |
Most of the epidemiological and animal-based research have shown an inverse relationship between colon cancer and consumption of high-fibre foods13. Among major dietary fibers constituents, a great deal of scientific investigation has been given to phytate due to its anti-cancerous activity against CRC46. In an in-vitro study, Yang and Shamsuddin47 showed an IP6-induced growth inhibition and differentiation of HT-29 human colon cancer cells.
Prostate cancer (PCA): The PCA is the most common, invasive and frequently diagnosed cancer type among men, after skin cancer. Among the majority of cases, the PCA starts in the prostate (a gland in the male reproductive system) and the condition is known as adenocarcinoma. In the USA, it is the second leading cause of cancer deaths in men48. In another study, Zi et al.49 observed that epidermal growth factor receptor (EGFR or erbB1) endocytosis and associated mitogenic signaling occur in human DU145 prostate cancer cells. It further suggests that erbB1 endocytosis might be involved in advanced and androgen-independent PCA growth. In their study, phytate impaired both receptor-mediated and fluid-phase endocytosis, resulting in the inhibition of mitogenic signals associated with growth and proliferation of human prostate carcinoma DU145 cells. The results obtained further suggest a novel molecular pathway be further explored for the intervention of advanced and androgen-independent human PCA by phytate.
Threatening key facts about PCA50:
| • | After skin cancer, PCA is the most common cancer among men in the US |
| • | PCA is one of the leading causes of cancer death among men of all races and Hispanic origin populations |
| • | In the US around 209,292 men are diagnosed with prostate cancer per year |
| • | Around 27,970 men die from prostate cancer in the US each year |
| • | According to the American Cancer Society, about 1 man in 7 will be diagnosed with prostate cancer during his lifetime |
| • | Prostate cancer mainly occurs in older men-about 6 cases in 10 are diagnosed in men 65 years or older |
| • | Almost all prostate cancers are adenocarcinomas-cancers that begin in cells that make and release mucus and other fluids |
| • | Prostate cancer often has no early symptoms |
| • | Advanced prostate cancer can cause men to urinate more often or have a weaker flow of urine |
| • | Most men diagnosed with prostate cancer do not die from it. More than 2.9 million men in the US diagnosed with prostate cancer at some point are still alive today |
Apart from another obvious risk factor, following risk factor majorly contributes in PCA.
| • | Age (primary risk factor): Prostate cancer is rare among men under the age of 45 but much more common after the age of 50 |
| • | Genetics (definite risk factor): Studies indicate that the two faulty genes-BRCA 1 and BRCA 2-which are important risk factors for breast cancer and ovarian cancer, have also been implicated in prostate cancer risk |
| • | Diet (secondary risk factor): The diet lacks with vitamin D and a diet high in red meat may raise a person’s chances of developing prostate cancer |
| • | Medication (secondary risk factor): Concerning additional medication, there might be a link between the daily use of anti-inflammatory medicines and prostate cancer risk |
| • | Obesity (definite risk factor): There is a clear link between obesity and raised prostate cancer risk, as well as a higher risk of metastasis and death among obese people who develop prostate cancer |
| • | Sexually transmitted disease (STDs) |
| • | Agent Orange (Depending upon exposure concentration, a higher risk factor) |
| • | Enzyme PRSS3. PRSS3, an enzyme, changes the environment of prostate cancer cells, making cancer much more likely to metastasize |
Efficacy against Parkinson’s disease: The phytic acid also termed as IP6 or inositol hexaphosphate is considered one of the most significant beneficial agent that can target Parkinson’s and Alzheimer’s disease. Apart from other useful aspects, the IP6 has following unique characteristics i.e., (1) Strong anticancer activity, (2) Protective effect against neurons from cell death, (3) Chelates iron, (4) It is cheap and safe and (5) Could help in preventing Parkinson’s disease. Evidently, according to the literature, IP6 has been proposed to treat Parkinson’s disease51. Parkinson’s is a second most common neurodegenerative disorder after Alzheimer’s52. This neurodegenerative disorder is specifically characterized by a selective dopamine neurons degeneration in the substantia nigra, resulting in irreversible motor dysfunction53. The substantia nigra part of the mammalian part has substantial quantities of iron which is believed to be a significant contributor to Parkinson’s disease. One possible reason for this dopamine neurons degeneration is closely related to the free iron content in those parts of the brain. The induced cell damage including apoptosis caused by an oxidative stress also plays a critical role in the neurodegeneration associated with the Parkinson’s disease54. It is, therefore, the proficiency of unbound iron to generate free radicals and induce oxidative stress, which is at the center of their deleterious effects. Figure 2 illustrates a comprehensive overview of this relationship. For instance, Xu et al.53 reported the neuroprotective influence of the natural iron chelator i.e., IP6 or phytic acid in a cell culture model of Parkinson’s disease.
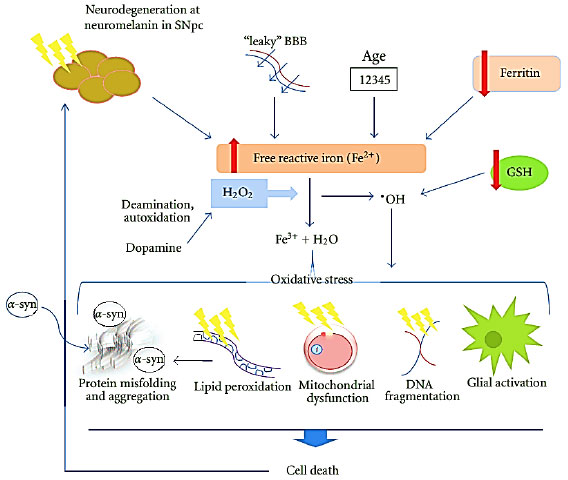 | |
| Fig. 2: | Iron-mediated cell death in PD. Reduced storage capacity in PD due to decreased ferritin expression and degeneration of nigral melatonin-containing neurons causes an increase in the reactive Fe2+ iron pool. Age-related increases in iron and a leaky BBB cause further iron accumulation. The transfer of the free iron to ferric iron, Fe3+, in the hydrogen peroxide-mediated Fenton reaction produces the highly toxic hydroxyl radical. A compromised level of glutathione exacerbates the levels of free radicals, whilst the deamination and autoxidation of dopamine produces further H2O2. The subsequent oxidative stress can then elicit a range of cytotoxic reactions including protein misfolding, lipid peroxidation (which, in turn, can cause α-synuclein aggregation), mitochondrial dysfunction and activation of glial cells. These various insults can induce cell death by apoptosis, causing further degeneration (Reproduced from Ref. Mounsey and Teismann51, an open access article distributed under the Creative Commons Attribution License) |
Authors have reported disrupted iron metabolism and excess iron accumulation in the brains of Parkinson’s disease patients. Besides an excellent chelator particularly for divalent minerals and reduce their absorption, IP6 has significantly considerable ability to alter cell signaling pathways and/or antioxidant catalysts (enzyme) that are responsible for ROS detoxification55. Figure 3 shows an action of iron chelators targeting Parkinson’s disease. A plethora of research studies have shown/reported anticancer role of IP6, however, the data on its effect on neurodegenerative diseases are limited53. Considering the above factors and outcomes from various studies, it could be helpful for Parkinson’s disease patients to consume high IP6/phytic acid containing natural products e.g., cereals and legumes, etc. However, future animal based models or human-related studies are needed to confirm the exact mechanism of action of IP6 under various conditions.
Biotechnological applications
Nutraceutical potential for animals: From the past several decades, the scientific literature is evidenced that the supplementation of phytase and their determinants in different diets for various species56-60. Phytase enzyme hydrolyzes phytic acid from cereals and releases bound Phosphate61,62. The addition of phytase in poultry rations can remove the inorganic P63. The activity of this extracellular phytase breaks down the bound phytate-P as well as releases the micronutrients (Zn, Iron) and reduces the excretion of P in feces64. The use of enzymes in poultry and animal diets improves digestibility and availability of nutrients. There is an economic, environmental and sustainable, for reuse of P65,66. Two types of phytases are 3-phytase (myo-inositol hexakisphosphate-3-phosphohydrolase, E.C. 3.1.3.8) that remove phosphate from the 3-position of phytate and a 6-phytase (myo-inositol hexakisphosphate 6-phosphohydrolase, E.C. 3.1.3.26) that removes phosphate from the 6-position of phytate67. Microbes extracellularly secrete the 3-phytase.
The use of phytase in nutrition supplements decreases the viscosity and increases the absorption of nutrients by ruminant which also reduces the number of feces68,69. Microbes are attracting significant industrial interest for extracellular secretion of phytases both for environmental and economic reasons70.
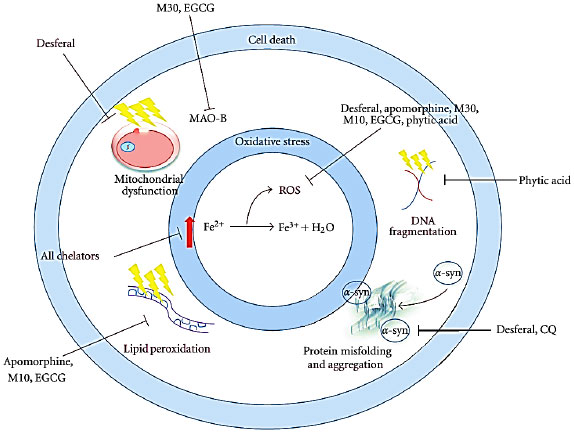 | |
| Fig. 3: | Action of iron chelators targeting PD. All iron chelators mop up excess free, reactive iron, thus reducing the reduction of Fe2+ to Fe3+; a reaction that produces various ROS, such as the hydroxyl radical. Oxidative stress resulting from the generation of ROS produces a range of deleterious insults, which can be targeted with the multiple actions of inhibitors. This can attenuate the cell death that these events induce. Chelators with antioxidant properties inhibit the production of ROS, in an environment of diminished antioxidant activity. The dopamine-oxidising enzyme MAO-B, which resides in the outer membrane of mitochondria, can also be inhibited by some chelators (Reproduced from Ref. Mounsey and Teismann51, an open access article distributed under the Creative Commons Attribution License) |
The net effect of using the enzyme in the diet increased the weight of the animal. Spray food processing enzymes just before feeding provides greater flexibility of management71. Feeding a treatment with enzymes in this manner can improve the digestibility of food through some different mechanisms72,73, including direct hydrolysis, improved palatability, viscosity changes of the intestine69.
Supplementation of diets with exogenous enzymes enhances the efficiency of poultry by increasing digestion of low-quality products and reducing nutrient losses with possible economic benefits. Enzymes increase digestibility, remove anti-nutritional factors and increase nutrient availability. In the current scenario, there is a need to develop versatile enzymes as a sustainable solution to problems of poor nutrient digestibility and bioavailability. The sustainability of phosphorus is emerging as a major societal goal to secure future food and water security for a growing population. The production systems can be redesigned to facilitate recovery and recycling of P. Phytase added in crops, which improve phytate phosphorous uses, reduces environmental pollution, making poultry husbandry ecologically more attractive. Looking at the big picture, it will help meet the new global targets for improving people’s lives around the world as an unprecedented push to tackle the root causes of poverty. It embraces the need for economic development that leaves no one behind and gives every human being a fair chance of leading a decent life thus to protect future generations by limiting climate change, adopting renewable energy and managing resources sustainably.
Concluding remarks: In summary, many research scientists, entrepreneurs and research-based organizations have been focused or be focusing their attention on phytase or phytate and their considerable exploitation for various fields including human health from biomedical and human and animal nutrition from nutraceutical. The above-discussed data suggest that a noteworthy potential exists for the use of phytase in various biotechnological applications at large and biomedical in particular. However, up to now, no single phytase based product for biomedical application has found its way to the market. Besides its antinutrient activity, dietary phytate displays advantageous health-related aspects, for example, higher protection efficacy against various cancer types and health-related diseases including Parkinson’s, Alzheimer’s, diabetes mellitus and renal stones. Considering the above-mentioned useful health effects, the phytase/phytate can play a highly significant role for populations in developed countries, because of the greater frequency of cancer particularly colon cancer which is related to the higher fat and lower fibre-rich food intakes.
SIGNIFICANCE STATEMENT
Phytase/phytate containing natural products have some inherent characteristics that make them potent candidate for biomedical and pharmaceutical applications. Within the last 15 years, the phytate has shown extraordinary potentialities though at laboratory level studies e.g., (1) To halt abnormal cell proliferation, (2) To treat various cancer types, (3) To alter cell signaling pathways and/or antioxidant catalysts (enzyme) that are responsible for ROS detoxification. Considering all the normal physiological facts present in our body, the phytase/phtate supplementation strongly argue in favor of its inclusion as an essential nutrient.
ACKNOWLEDGMENTS
This review work was done as a part of the project entitled "Development of immobilized fungal phytase to improve nutrient utilization of agro-based residues in poultry feed". The authors would like to thank Higher Education Commission (HEC), Islamabad, Pakistan for funding the project #6490/Punjab/NRPU/R&D/HEC/2016. The authors are also grateful to the Department of Biochemistry and Biotechnology of the University of Gujrat, Pakistan for providing the literature facilities.
REFERENCES
- Suzuki, U., K. Yoshimura and M. Takaishi, 1907. Ueber ein Enzym “Phytase” das “Anhydro-oxy-methylen diphosphorsaure” spaltet. Tokyo Imper. Univ. Coll. Agric. Bull., 7: 503-512.
Direct Link - George, T.S., P.J. Gregory, M. Wood, D. Read and R.J. Buresh, 2002. Phosphatase activity and organic acids in the rhizosphere of potential agroforestry species and maize. Soil Biol. Biochem., 34: 1487-1494.
CrossRefDirect Link - Konietzny, U. and R. Greiner, 2002. Molecular and catalytic properties of phytate-degrading enzymes (phytases). Int. J. Food Sci. Technol., 37: 791-812.
CrossRefDirect Link - Wang, X., S. Upatham, W. Panbangred, D. Isarangkul, P. Summpunn, S. Wiyakrutta and V. Meevootisom, 2004. Purification, characterization, gene cloning and sequence analysis of a phytase from Klebsiella pneumoniae subsp. pneumoniae XY-5. ScienceAsia, 30: 383-390.
Direct Link - Jorquera, M., O. Martinez, F. Maruyama, P. Marschner and M. de la Luz Mora, 2008. Current and future biotechnological applications of bacterial phytases and phytase-producing bacteria. Microb. Environ., 23: 182-191.
CrossRefDirect Link - Balaban, N.P., A.D. Suleimanova, L.R. Valeeva, I.B. Chastukhina, N.L. Rudakova, M.R. Sharipova and E.V. Shakirov, 2016. Microbial phytases and phytate: Exploring opportunities for sustainable phosphorus management in agriculture. Am. J. Mol. Biol., 7: 11-29.
CrossRefDirect Link - Mullaney, E.J. and A.H.J. Ullah, 2003. The term phytase comprises several different classes of enzymes. Biochem. Biophys. Res. Commun., 312: 179-184.
CrossRefPubMedDirect Link - Lei, X.G., J.D. Weaver, E. Mullaney, A.H. Ullah and M.J. Azain, 2013. Phytase, a new life for an “old” enzyme. Annu. Rev. Anim. Biosci., 1: 283-309.
CrossRefDirect Link - Chu, H.M., R.T. Guo, T.W. Lin, C.C. Chou and H.L. Shr et al., 2004. Structures of Selenomonas ruminantium phytase in complex with persulfated phytate: DSP phytase fold and mechanism for sequential substrate hydrolysis. Structure, 12: 2015-2024.
CrossRefDirect Link - Vats, P. and U.C. Banerjee, 2004. Production studies and catalytic properties of phytases (myo-inositolhexakisphosphate phosphohydrolases): An overview. Enzyme Microb. Technol., 35: 3-14.
CrossRefDirect Link - Oh, B.C., W.C. Choi, S. Park, Y.O. Kim and T.K. Oh, 2004. Biochemical properties and substrate specificities of alkaline and histidine acid phytases. Applied Microbiol. Biotechnol., 63: 362-372.
CrossRefPubMedDirect Link - Kumar, V., A.K. Sinha, H.P.S. Makkar and K. Becker, 2010. Dietary roles of phytate and phytase in human nutrition: A review. Food Chem., 120: 945-959.
CrossRefDirect Link - Schroder, B., G. Breves and M. Rodehutscord, 1996. Mechanisms of intestinal phosphorus absorption and availability of dietary phosphorus in pigs. Deutsche Tierarztliche Wochenschrift, 103: 209-214.
PubMedDirect Link - Shamsuddin, A.M., 2002. Anti-cancer function of phytic acid. Int. J. Food Sci. Technol., 37: 769-782.
CrossRefDirect Link - Vucenik, I. and A.M. Shamsuddin, 2003. Cancer inhibition by inositol hexaphosphate (IP6) and inositol: From laboratory to clinic. J. Nutr., 33: 3778S-3784S.
PubMedDirect Link - Thompson, L.U., 1993. Potential health benefits and problems associated with antinutrients in foods. Food Res. Int., 26: 131-149.
CrossRefDirect Link - Barker, C.J. and P.O. Berggren, 1999. Inositol hexakisphosphate and beta-cell stimulus-secretion coupling. Anticancer Res., 19: 3737-3741.
PubMed - Onomi, S., Y. Okazaki and T. Katayama, 2004. Effect of dietary level of phytic acid on hepatic and serum lipid status in rats fed a high-sucrose diet. Biosci. Biotechnol. Biochem., 68: 1379-1381.
CrossRefPubMedDirect Link - Grases, F., R.M. Prieto, B.M. Simonet and J.G. March, 2000. Phytate prevents tissue calcifications in female rats. BioFactors, 11: 171-177.
CrossRefDirect Link - Minihane, A.M. and G. Rimbach, 2002. Iron absorption and the iron binding and anti-oxidant properties of phytic acid. Intl. J. Food Sci. Technol., 37: 741-748.
Direct Link - Selvam, R., 2002. Calcium oxalate stone disease: Role of lipid peroxidation and antioxidants. Urol. Res., 30: 35-47.
CrossRefDirect Link - Otake, T., H. Shimonaka, M. Kanai, K. Miyano, N. Ueba, N. Kunita and T. Kurimura, 1989. [Inhibitory effect of inositol hexasulfate and inositol hexaphosphoric acid (phytic acid) on the proliferation of the Human Immunodeficiency Virus (HIV) in vitro]. J. Jpn. Assoc. Infect. Dis., 63: 676-683, (In Japanese).
PubMed - Otake, T., H. Mori, M. Morimoto, K. Miyano and N. Ueba et al., 1999. Anti-HIV-1 activity of myo-inositol hexaphosphoric acid (IP6) and myo-inositol hexasulfate (IS6). Anticancer Res., 19: 3723-3726.
PubMed - Sakamoto, K., G. Venkatraman and A.M. Shamsuddin, 1993. Growth inhibition and differentiation of HT-29 cells in vitro by inositol hexaphosphate (phytic acid). Carcinogenesis, 14: 1815-1819.
CrossRefDirect Link - Deliliers, G.L., F. Servida, N.S. Fracchiolla, C. Ricci, C. Borsotti, G. Colombo and D. Soligo, 2002. Effect of inositol hexaphosphate (IP6) on human normal and leukaemic haematopoietic cells. Br. J. Haematol., 117: 577-587.
CrossRefDirect Link - Shamsuddin, A.M., G.Y. Yang and I. Vucenik,. 1996. Novel anti-cancer functions of IP6: growth inhibition and differentiation of human mammary cancer cell lines in vitro. Anticancer Res., 16: 3287-3292.
PubMedDirect Link - Sandra, F., M. Matsuda, H. Yoshida and M. Hirata, 2002. Inositol hexakisphosphate blocks tumor cell growth by activating apoptotic machinery as well as by inhibiting the Akt/NFκB-mediated cell survival pathway. Carcinogenesis, 23: 2031-2041.
CrossRefDirect Link - Singh, R.P., C. Agarwal and R. Agarwal, 2003. Inositol hexaphosphate inhibits growth and induces G1 arrest and apoptotic death of prostate carcinoma DU145 cells: Modulation of CDKI-CDK-cyclin and pRb-related protein-E2F complexes. Carcinogenesis, 24: 555-563.
CrossRefDirect Link - Agarwal, C., S. Dhanalakshmi, R.P. Singh and R. Agarwal, 2004. Inositol hexaphosphate inhibits growth and induces G1 arrest and apoptotic death of androgen-dependent human prostate carcinoma LNCaP cells. Neoplasia, 6: 646-659.
CrossRefDirect Link - Vucenik, I., K. Tantivejkul, Z.S. Zhang, K.E. Cole, I. Saied and A.M. Shamsuddin, 1998. IP6 in treatment of liver cancer. I. IP6 inhibits growth and reverses transformed phenotype in HepG2 human liver cancer cell line. Anticancer Res., 18: 4083-4090.
PubMedDirect Link - Soni, S.K., S. Sarkar, P.R. Selvakannan, D. Sarkar and S.K. Bhargava, 2015. Intrinsic therapeutic and biocatalytic roles of ionic liquid mediated self-assembled platinum-phytase nanospheres. RSC Adv., 5: 62871-62881.
CrossRefDirect Link - Barahuie, F., D. Dorniani, B. Saifullah, S. Gothai and M.Z. Hussein et al., 2017. Sustained release of anticancer agent phytic acid from its chitosan-coated magnetic nanoparticles for drug-delivery system. Int. J. Nanomed., 12: 2361-2372.
CrossRefDirect Link - Johns, L.E. and R.S. Houlston, 2001. A systematic review and meta-analysis of familial colorectal cancer risk. Am. J. Gastroenterol., 96: 2992-3003.
CrossRefDirect Link - Srinivasan, R., Y.X. Yang, S.C. Rubin, M.A. Morgan and J.D. Lewis, 2007. Risk of colorectal cancer in women with a prior diagnosis of gynecologic malignancy. J. Clin. Gastroenterol., 41: 291-296.
CrossRefDirect Link - Singh, H., Z. Nugent, A. Demers, P.M. Czaykowski and S.M. Mahmud, 2013. Risk of colorectal cancer after diagnosis of endometrial cancer: A population-based study. J. Clin. Oncol., 31: 2010-2015.
CrossRefDirect Link - Imperiale, T.F., R. Juluri, E.A. Sherer, E.A. Glowinski, C.S. Johnson and M.S. Morelli, 2014. A risk index for advanced neoplasia on the second surveillance colonoscopy in patients with previous adenomatous polyps. Gastrointest. Endosc., 80: 471-478.
CrossRefDirect Link - Mork, M.E., Y.N. You, J. Ying, S.A. Bannon, P.M. Lynch, M.A. Rodriguez-Bigas and E. Vilar, 2015. High prevalence of hereditary cancer syndromes in adolescents and young adults with colorectal cancer. J. Clin. Oncol., 33: 3544-3549.
CrossRefDirect Link - Fedirko, V., I. Tramacere, V. Bagnardi, M. Rota and L. Scotti et al., 2011. Alcohol drinking and colorectal cancer risk: An overall and dose-response meta-analysis of published studies. Ann. Oncol., 22: 1958-1972.
CrossRefDirect Link - Liang, P.S., T.Y. Chen and E. Giovannucci, 2009. Cigarette smoking and colorectal cancer incidence and mortality: Systematic review and meta-analysis. Int. J. Cancer, 124: 2406-2415.
CrossRefDirect Link - Ma, Y., Y. Yang, F. Wang, P. Zhang, C. Shi, Y. Zou and H. Qin, 2013. Obesity and risk of colorectal cancer: A systematic review of prospective studies. PLoS ONE, Vol. 8.
CrossRefDirect Link - Laiyemo, A.O., C. Doubeni, P.F. Pinsky, V.P. Doria-Rose and R. Bresalier et al., 2010. Race and colorectal cancer disparities: Health-care utilization vs different cancer susceptibilities. J. Natl. Cancer Inst., 102: 538-546.
CrossRefDirect Link - Lansdorp-Vogelaar, I., K.M. Kuntz, A.B. Knudsen, M. van Ballegooijen, A.G. Zauber and A. Jemal, 2012. Contribution of screening and survival differences to racial disparities in colorectal cancer rates. Cancer Epidemiol. Prev. Biomarkers, 21: 728-736.
CrossRefDirect Link - Shamsuddin, A.M. and I. Vucenik, 1999. Mammary tumor inhibition by IP6: A review. Anticancer Res., 19: 3671-3674.
PubMedDirect Link - Yang, G. and A.M. Shamsuddin, 1995. IP6-induced growth inhibition and differentiation of HT-29 human colon cancer cells: Involvement of intracellular inositol phosphates. Anticancer Res., 15: 2479-2488.
Direct Link - Godley, P.A., M.K. Campbell, P. Gallagher, F.E. Martinson, J.L. Mohler and R.S. Sandler, 1996. Biomarkers of essential fatty acid consumption and risk of prostatic carcinoma. Cancer Epidemiol. Prev. Biomarkers, 5: 889-895.
Direct Link - Zi, X., R.P. Singh and R. Agarwal, 2000. Impairment of erbB1 receptor and fluid-phase endocytosis and associated mitogenic signaling by inositol hexaphosphate in human prostate carcinoma DU145 cells. Carcinogenesis, 21: 2225-2235.
CrossRefDirect Link - Mounsey, R.B. and P. Teismann, 2012. Chelators in the treatment of iron accumulation in Parkinson's disease. Int. J. Cell Biol.
CrossRefDirect Link - De Lau, L.M. and M.M. Breteler, 2006. Epidemiology of Parkinson's disease. Lancet Neurol., 5: 525-535.
CrossRefDirect Link - Xu, Q., A.G. Kanthasamy and M.B. Reddy, 2008. Neuroprotective effect of the natural iron chelator, phytic acid in a cell culture model of Parkinson's disease. Toxicology, 245: 101-108.
CrossRefPubMedDirect Link - Burke, R.E. and N.G. Kholodilov, 1998. Programmed cell death: Does it play a role in Parkinson's disease? Ann. Neurol., 44: S126-S133.
CrossRefDirect Link - Shamsuddin, A.M., I. Vucenik and K.E. Cole, 1997. IP6: A novel anti-cancer agent. Life Sci., 61: 343-354.
CrossRefDirect Link - Lei, X.G. and J.M. Porres, 2003. Phytase enzymology, applications and biotechnology. Biotechnol. Lett., 25: 1787-1794.
CrossRefDirect Link - Selle, P.H. and V. Ravindran, 2008. Phytate-degrading enzymes in pig nutrition. Livest. Sci., 113: 99-122.
CrossRefDirect Link - Adeola, O. and A.J. Cowieson, 2011. Board-invited review: Opportunities and challenges in using exogenous enzymes to improve nonruminant animal production. J. Anim. Sci., 89: 3189-3218.
CrossRefDirect Link - Kumar, V., A.K. Sinha, H.P.S. Makkar, G. De Boeck and K. Becker, 2012. Phytate and phytase in fish nutrition. J. Anim. Physiol. Anim. Nutr., 96: 335-364.
CrossRefDirect Link - Bilal, T., S. Atis and O. Keser, 2015. The effects of microbial phytase on serum calcium and phosphorus levels and alkaline phosphatase activities in broilers fed diets containing different levels of phosphorus. Acta Scientiae Veterinariae, Vol. 43.
Direct Link - Tu, S., L. Ma and B. Rathinasabapathi, 2011. Characterization of phytase from three ferns with differing arsenic tolerance. Plant Physiol. Biochem., 49: 146-150.
CrossRefDirect Link - Joseph, I. and R.P. Raj, 2007. Isolation and characterization of phytase producing Bacillus strains from mangrove ecosystem. J. Mar. Biol. Assoc. India, 49: 177-182.
Direct Link - Butani, J.B. and S. Parnerkar, 2015. Role of microbial phytase in broiler nutrition-A review. J. Livest. Sci., 6: 113-118.
Direct Link - Schipanski, M.E. and E.M. Bennett, 2012. The influence of agricultural trade and livestock production on the global phosphorus cycle. Ecosystems, 15: 256-268.
CrossRefDirect Link - Withers, P.J., J.J. Elser, J. Hilton, H. Ohtake, W.J. Schipper and K.C. van Dijk, 2015. Greening the global phosphorus cycle: How green chemistry can help achieve planetary P sustainability. Green Chem., 17: 2087-2099.
CrossRefDirect Link - Joshi, J.B., 2014. Phytase-a key to unlock phytate complex. Int. J. Pure Applied Biosci., 2: 304-313.
Direct Link - Kumar, G.P. and V. Suneetha, 2014. A cocktail enzyme: Pectinase from fruit industrial dump sites: A review. Res. J. Pharm. Biol. Chem. Sci., 5: 1252-1258.
Direct Link - Garg, G., A. Singh, A. Kaur, R. Singh, J. Kaur and R. Mahajan, 2016. Microbial pectinases: An ecofriendly tool of nature for industries. 3 Biotech., Vol. 6, No. 1.
CrossRefDirect Link - Sumengen, M., S. Dincer and A. Kaya, 2013. Production and characterization of phytase from Lactobacillus plantarum. Food Biotechnol., 27: 105-118.
CrossRefPubMedDirect Link - Sato, V.S., J.A. Jorge, W.P. Oliveira, C.R.F. Souza and L.H.S. Guimaraes, 2014. Phytase production by Rhizopus microsporus var. microsporus biofilm: Characterization of enzymatic activity after spray drying in presence of carbohydrates and nonconventional adjuvants. J. Microbiol. Biotechnol., 24: 177-187.
CrossRefDirect Link - Nielsen, A.V.F., I. Tetens, S. Anne and A.S. Meyer, 2013. Potential of phytase-mediated iron release from cereal-based foods: A quantitative view. Nutrients, 5: 3074-3098.
CrossRefDirect Link - Sasirekha, B., T. Bedashree and K.L. Champa, 2012. Optimization and partial purification of extracellular phytase from Pseudomonas aeruginosa p6. Eur. J. Exp. Biol., 2: 95-104.
Direct Link