
Jung Hun Lee
Department of Biological Sciences, Myongji University, San 38-2 Namdong, Yongin,
Gyeouggido, 449-728, Republic of Korea
Seung Jin Bae
Department of Biological Sciences, Myongji University, San 38-2 Namdong, Yongin,
Gyeouggido, 449-728, Republic of Korea
Jae Jin Lee
Department of Biological Sciences, Myongji University, San 38-2 Namdong, Yongin,
Gyeouggido, 449-728, Republic of Korea
Il Kwon Bae
Department of Laboratory Medicine, College of Medicine, Kosin University,
Busau 602-702, Republic of Korea
Seok Hoon Jeong
Department of Laboratory Medicine, College of Medicine, Kosin University,
Busau 602-702, Republic of Korea
Eui Suk Sohn
Division of Antimicrobial Resistance, Infectious Disease Research Center, National Institute of Health, Korea Center of Disease Control and Prevention, Seoull22-701, Republic of Korea
Yeong Seon Lee
Division of Antimicrobial Resistance, Infectious Disease Research Center, National Institute of Health, Korea Center of Disease Control and Prevention, Seoull22-701, Republic of Korea
Sang Hee Lee
Department of Biological Sciences, Myongji University, San 38-2 Namdong, Yongin,
Gyeouggido, 449-728, Republic of Korea
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 5 | Page No.: 486-490







ABSTRACT
In our proposed nomenclature system including GES-type (Guiana-Extended-Spectrum β-lactamase) or IBC-type (Integron-Borne Cephalosporinase) extended-spectrum β-lactamases (ESBLs), GES-2 has an amino acid substitution of G170N, GES-3 has two M62T and E104K substitutions, GES-4 has three M62T, E104K and G170S substitutions, GES-5 has a G170S substitution, GES-6 has two F104K and G170S substitutions, GES-7 (IBC-1) has a G104K substitution, GES-8 (IBC-2) has a A125L substitution and GES-9 has a G243S substitution, compared to the sequence of GES-1. This proposed nomenclature update would not confuse microbiologists studying GES-type ESBLs, fundamentally preventing misleading nomenclature of these antibiotic resistance genes. The definitive renaming of GES/IBC-type ESBLs can help also some researchers to correctly designate new GES-type ESBLs such as novel enzymes identified from some nationwide surveys. In order to detect rapidly and exactly GES-type ESBLs with different hydrolysis profiles and inhibitor-resistance patterns by pyrosequencing, we also proposed here the new four key amino acid substitutions and four GES-type variants as follows: (i) GES-1 able to hydrolyze ceftazidime, (ii) GES-4 able to hydrolyze ceftazidime, cefoxitin and imipenem and able to show inhibitor-resistance, (iii) GES-5 able to hydrolyze ceftazidime, cefoxitin and imipenem and (iv) GES-9 able to hydrolyze ceftazidime and aztreonam.
PDF Abstract XML References
How to cite this article
Jung Hun Lee, Seung Jin Bae, Jae Jin Lee, Il Kwon Bae, Seok Hoon Jeong, Eui Suk Sohn, Yeong Seon Lee and Sang Hee Lee, 2007. Four Exact GES Variants Used for Identification of GES-type Extended-spectrum β-lactamases with Different Hydrolysis Profiles and Inhibitor-Resistance Patterns by Pyrosequencing. Research Journal of Microbiology, 2: 486-490.
URL: https://scialert.net/abstract/?doi=jm.2007.486.490
URL: https://scialert.net/abstract/?doi=jm.2007.486.490
INTRODUCTION
The β-lactamases produced by bacterium are known to protect against the lethal effect of penicillins, cephalosporins, carbapenems, or monobactams on cell-wall synthesis. The production of β-lactamase is the single most prevalent mechanism responsible for resistance to β-lactams among clinical isolates of Pseudomonas aeruginosa and the family Enterobacteriaceae and thus, pose therapeutic problems (Sanders and Sanders, 1992). The Antimicrobial Availability Task Force (AATF) of the Infectious Diseases Society of America (IDSA) reported ESBL (extended-spectrum β-lactamases)-producing Enterobacteriaceae as a bacterium of particularly problematic pathogens (George et al., 2006). ESBLs are clavulanate-susceptible enzymes conferring broad resistance to penicillins, aztreonam and cephalosporins (with the exception of cephamycins) (Livermore, 1995). ESBLs are often plasmid mediated and most of them are mutants of the classic TEM and SHV enzymes, with one or more amino-acid substitutions around active site (Paterson et al., 2001). These changes allow hydrolysis of extended-spectrum cephalosporins (e.g., ceftazidime and cefotaxime) and monobactams (e.g., aztreonam), which are stable to classic TEM and SHV enzymes (Bradford, 2001). There are also reports of clinical isolates of Enterobacteriaceae producing various non-TEM, non-SHV ESBLs: CTX-M and GES/IBC-types (Bradford, 2001; Lee et al., 2005b). In particular, GES/IBC-type ESBLs is greatly problematic because these ESBLs can be non-metallo-carbapenemases hydrolyzing carbapenems (e.g., imipenem and meropenem), which have the broadest activity spectra of any β-lactam antibiotics and are often the most appropriate agents for use in the treatment of infections caused by multi-resistant Gram-negative aerobic bacteria (Lee and Lee, 2006). Therefore, rapid identification technique and typing of GES/IBC-type ESBLs are very important for studying the epidemiology and prevalence of carbapenem-hydrolyzing and ESBL-producing pathogens. The exact nomenclature systems of GES/IBC-type ESBLs are also necessary for the typing of these ESBLs and can help also some researchers to correctly designate new GES-type ESBLs such as novel enzymes identified from some nationwide surveys. However, in recently published articles (Jeong et al., 2005; Lee et al., 2005a; Poirel et al., 2005, 2006; Ryoo et al., 2005), the nomenclature systems of GES-type ESBLs still cause confusion. Poirel et al. (2006) described a rapid pyrosequencing technique for identification of GES-type ESBLs because GES variants are the only ESBLs that possess different hydrolysis profiles, including expanded-spectrum cephalosporins, carbapenems, cephamycins and monobactams. However, there are fraught with misleading nomenclature of GES-type ESBLs, faulty definition of the four different hydrolysis spectra of GES enzymes and careless selection of four GES variants that are representatives of the four different hydrolysis profiles. Therefore, the study was aimed to define the exact designation of GES-type ESBLs and four GES variants used for identification of GES-type ESBLs with different hydrolysis profiles and inhibitor-resistance patterns by pyrosequencing.
The Exact Designation of GES/IBC-type ESBLs
The definitive nomenclature system including GES/IBC-type ESBLs was revised by our recent paper (Lee et al., 2005a) and described in a β-lactamase website (http://www.lahey.org/ studies/webt.asp) by our personal communication with George Jacoby (a keeper of the website). In the definitive nomenclature system, we proposed to maintain the current denomination concerning the fully characterized GES-3 and GES-4 enzymes reported by Wachino et al. (2004a and b) from Japan and to rename the variants (GES-3 and GES-4) reported by Vourli et al. (2004) from Greece as GES-5 and GES-6, respectively. However, Poirel et al. (2006) designated incorrectly the variants reported by Wachino et al. (2004a, b) as GES-5 and GES-6 (for GES-3 and GES-4, respectively), although they cited the current GES nomenclature defined in the website. Thus, GES-3, -4, -5 and -6 described by Poirel et al. (2006) should change into GES-5, -6, -3 and -4, respectively. In the exact nomenclature system of the website, GES-2 has an amino acid substitution of G170N, GES-3 has two M62T and E104K substitutions, GES-4 has three M62T, E104K and G170S substitutions, GES-5 has a G170S substitution and GES-6 has two F104K and G170S substitutions, compared to the sequence of GES-1. In actuality, GES-3 emerged in Korea is GES-5 in the website, which has been confirmed by our personal communication with Ryoo et al. (2005). We have also recently reported (Jeong et al., 2005) that the GES-5 gene which has broken out in Korea differed by just one silent mutation (G-A) at position 54 from the variant (GenBank accession No. AY494717) reported by Vourli et al. (2004). Poirel et al. (2005) have also applied the misleading nomenclature system to their article describing GES-9. This misleading nomenclature system confuse microbiologists studying GES-type ESBLs and can not help some researchers to designate correctly new GES-type ESBLs such as novel enzymes identified from some nationwide surveys. The novel GES-type ESBLs can be identified by not the pyrosequencing technique but the whole direct sequencing technique.
The Exact Definition of the Four Different Hydrolysis Spectra of GES-type ESBLs for Pyrosequencing Technique
Poirel et al. (2006) stated that four GES variants which has used for pyrosequencing technique were as follows: (i) GES-1 able to hydrolyze ceftazidime, (ii) GES-2 able to hydrolyze ceftazidime and imipenem, (iii) GES-3 (actually GES-5) able to hydrolyze ceftazidime, cefoxitin and imipenem and (iv) GES-9 able to hydrolyze ceftazidime and aztreonam. The cefoxitin-hydrolyzing ability of GES-3 (actually GES-5) identified in Greece has been unknown since Vourli et al. (2004) did not perform the biochemical characterization and susceptibility testing of the GES variant on cefoxitin. Without the presentation of kinetic parameters, Poirel et al. (2006) stated that GES-3 (actually GES-5) could hydrolyze cefoxitin. The four different hydrolysis spectra defined by Poirel et al. (2006) do not include also capability to be resistant to inhibitors (e.g., clavulanic acid, sulbactam and tazobactam). In accordance with our biochemical characterization of GES-5 enzyme identified in Greece and Korea (not Japan), GES-5 hydrolyzed imipenem more efficiently than GES-2 and GES-4 and GES-4 hydrolyzed cefoxitin more efficiently than GES-5. That is, the hydrolysis efficiency (kcat/Km) of GES-5 for imipenem was three- and thirty-fold higher than that of GES-4 and GES-2, respectively due to a three- and thirty-fold-higher kcat values (personal data). GES-4 hydrolyzed cefoxitin most efficiently among GES variants and was resistant to inhibitors (Wachino et al., 2004a). The hydrolysis efficiency of GES-4 for cefoxitin was seven-fold higher than that of GES-5 (personal data). Thus, it is reasonable that GES-2 can be replaced by GES-4 among four GES variants used for pyrosequencing technique. Taking this change into account, new PCR primer sets and pyrosequencing primers for the analyses of codons 62 and 104 have to be designed for identification of GES-type ESBLs with different hydrolysis profiles and inhibitor-resistance patterns. Therefore, we propose here the new four key amino acid substitutions and four GES variants for identification of GES-type ESBLs with different hydrolysis profiles and inhibitor-resistance patterns by pyrosequencing (Table 1). Poirel et al. (2005) stated that GES-4, GES-5 and GES-6 variants hydrolyze carbapenems at a level as low as that for GES-2, which also hydrolyzes cephamycins. However, GES-2 could not hydrolyze cephamycins, whereas GES-4 could hydrolyze them (Wachino et al., 2004a).
The Drawback of Pyrosequencing Technique
Pyrosequencing is a gel-free, sequencing-by-synthesis technique that allows fast identification of short DNA sequences and has already been used a rapid and convenient approach for detection of antibiotic resistances (Poirel et al., 2006).
| Table 1: | Key amino acid substitutions (bold type) and GES variants for identification of GES-type ESBLs with different hydrolysis profiles and inhibitor-resistance patterns by pyrosequencing |
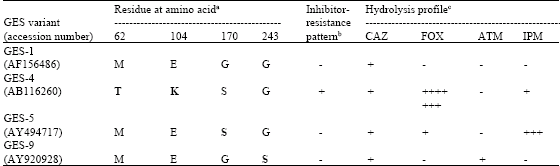 | |
| aAmber’s position (1980). bInhibitors are clavulanic acid, sulbactam and tazobactam. +, resistant; -, sensitive. c+++++++, seven-fold more efficient hydrolysis than GES-5; +++, three-fold more efficient hydrolysis than GES-4; +, hydrolysis; -, no hydrolysis; CAZ, Ceftazidime; FOX, Cefoxitin; ATM, Aztreonam; IPM, Imipenem | |
The overall experiment of the pyrosequencing technique is based on a protocol detailed within the AB Biotage website (http://www.pyrosequencing.com). The pyrosequencing technique has been coupled with PCR-based technique. The major drawback of PCR-based techniques is that they distinguish between just a few variants due to their limited multiplexing capabilities. The DNA microarray technology for the genotypic screening of 102 TEM (named after the patient [Temoneira] providing the first sample) β-lactamase variants was recently described (Grimm et al., 2004). This method has been shown to be a reproducible and sensitive means for the identification of ESBL variants which cannot be identified by simple phenotypic screening. With an assay time of less than 3.5 h, this method can be a rapid method for the detection of ESBLs and IRTs (Inhibitor resistant TEM beta-lactamases) and offers a valuable tool that can be used to monitor the spread of these resistance genes in the routine clinical diagnostic laboratory (Grimm et al., 2004). The length of detection time may be reduced further by replacing 102 TEM variants by nine GES variants. The pyrosequencing technique allows identification in additional more than 2 h of 100 β-lactamase variants. From extraction to sequencing results, the pyrosequencing technique takes more than 5 h in case of 100 variants. Although the pyrosequencing technique would be suitable to distinguish GES-1 ESBL from GES variants with different hydrolysis profiles and inhibitor-resistance patterns, the microarray technique could be suitable in case of too many variants.
CONCLUSIONS
GES variants are the only ESBLs that possess different hydrolysis profiles, including expanded-spectrum cephalosporins, carbapenems, cephamycins and monobactams. Therefore, the rapid identification technique and the exact nomenclature systems of GES/IBC-type ESBLs are very important for studying the epidemiology and prevalence of carbapenem-hydrolyzing and ESBL-producing pathogens. GES-type ESBLs should be correctly designated in accordance with our proposed nomenclature system including GES/IBC-type ESBLs. In our proposed nomenclature system, GES-2 has an amino acid substitution of G170N, GES-3 has two M62T and E104K substitutions, GES-4 has three M62T, E104K and G170S substitutions, GES-5 has a G170S substitution, GES-6 has two F104K and G170S substitutions, GES-7 (IBC-1) has a G104K substitution, GES-8 (IBC-2) has a A125L substitution and GES-9 has a G243S substitution, compared to the sequence of GES-1. Furthermore, we proposed that four GES variants which has used for pyrosequencing technique were as follows: (i) GES-1 able to hydrolyze ceftazidime, (ii) GES-4 able to hydrolyze ceftazidime, cefoxitin and imipenem and able to show inhibitor-resistance, (iii) GES-5 able to hydrolyze ceftazidime, cefoxitin and imipenem and (iv) GES-9 able to hydrolyze ceftazidime and aztreonam. In order to distinguish too many ESBL variants, the pyrosequencing technique would be less suitable than the microarray technique because of limited multiplexing capabilities of the PCR-based technique coupled with pyrosequencing.
ACKNOWLEDGMENTS
This work was supported by the National Institute of Health of KCDC in Republic of Korea, BioGreen 21 Program (20050301034479) of Rural Development Administration in Republic of Korea, the Driving Force Project for the Next Generation of Gyeonggi Provincial Government in Republic of Korea and the Second-Phase of Brain Korea 21 Project.
REFERENCES
- Ambler, R.P., 1980. The structure of beta-lactamases. Philos. Trans. Royal Soc. B Biol. Sci., 289: 321-331.
PubMed - Bradford, P.A., 2001. Extended-spectrum β-lactamases in the 21st century: Characterization, epidemiology and detection of this important resistance threat. Clin. Microbiol. Rev., 14: 933-951.
CrossRefPubMedDirect Link - George, H.T., J. Bradley, J.E. Edwards, Jr., D. Gilbert, M. Scheld and J.G. Bartlett, 2006. Bad bugs need drugs an update on the development pipeline from the antimicrobial availability task force of the Infectious diseases society of america. Clin. Infect. Dis., 42: 657-668.
Direct Link - Grimm, V., S. Ezaki, M. Susa, C. Knabbe, R.D. Schmid and T.T. Bachmann, 2004. Use of dna microarrays for rapid genotyping of tem β-lactamases that confer resistance. J. Clin. Microbiol., 42: 3766-3774.
CrossRef - Jeong, S.H., I.K. Bae, D. Kim, S.G. Hong, J.S. Song, J.H. Lee and S.H. Lee, 2005. First outbreak of Klebsiella pneumoniae clinical isolates producing GES-5 and SHV-12 extended-spectrum β-lactamases in Korea. Antimicrob. Agents Chemother., 49: 4809-4810.
CrossRef - Lee, S.H., J.Y. Kim, G.S. Lee, S.H. Cheon, Y.J. An, S.H. Jeong and K.J. Lee, 2002. Characterization of blaCMY-11, an AmpC-type plasmid-mediated β-lactamase gene in a Korean clinical isolate of Escherichia coli. J. Antimicrob. Chemother., 49: 269-273.
CrossRefDirect Link - Lee, S.H., S.H. Jeong, J.-I. Wachino, T. Arakawa, L. Poirel and P. Nordmann, 2005. Nomenclature of GES-type extended-spectrum β-lactamases. Antimicrob. Agents Chemother., 49: 2148-2150.
CrossRef - Lee, S.H., S.H. Jeong and S.S. Cha, 2005. Minimising antibiotic resistance. Lancet Infect. Dis., 5: 668-670.
CrossRef - Lee, J.H. and S.H. Lee, 2006. Carbapenem resistance in gram-negative pathogens: Emerging Non-metallo-carbapenemases. Res. J. Microbiol., 1: 1-22.
CrossRefDirect Link - Livermore, D.M., 1995. Β-lactamases in laboratory and clinical resistance. Clin. Microbiol. Rev., 8: 557-584.
Direct Link - Paterson, D.L., W.C. Ko, A. von Gottberg, J.M. Casellas and L. Mulazimoglu et al., 2001. Outcome of cephalosporin treatment for serious infections due to apparently susceptible organisms producing extended-spectrum β-lactamases implications for the clinical microbiology laboratory. J. Clin. Microbiol., 39: 2206-2212.
CrossRef - Poirel, L., L. Brinas, N. Fortineau and P. Nordmann, 2005. Integron-encoded ges-type extended-spectrum β-lactamase with increased activity toward aztreonam in Pseudomonas aeruginosa. Antimicrob. Agents Chemother., 49: 3593-3597.
CrossRef - Poirel, L., T. Naas and P. Nordmann, 2006. Pyrosequencing as a rapid tool for identification of GES-type extended-spectrum β-lactamases. J. Clin. Microbiol., 44: 3008-3011.
CrossRef - Ryoo, N.H., E.C. Kim, S.G. Hong, Y.J. Park and K. Lee et al., 2005. Dissemination of SHV-12 and CTX-M-type extended-spectrum β-lactamases among clinical isolates of Escherichia coli and Klebsiella pneumoniae and emergence of GES-3 in Korea. J. Antimicrob. Chemother., 56: 698-702.
CrossRef - Sanders, C.C. and W.E. Jr, Sanders, 1992. Β-lactamase in gram-negative bacteria global trends and clinical impact. Clin. Infect. Dis., 15: 824-839.
Direct Link - Vourli, S., P. Giakkoupi, V. Miriagou, E. Tzelepi, A.C. Vatopoulos and L.S. Tzouvelekis, 2004. Novel GES/IBC extended-spectrum β-lactamase variants with carbapenemase activity in clinical enterobacteria. FEMS Microbiol. Lett., 234: 209-213.
PubMedDirect Link - Wachino, J.I., Y. Doi, K. Yamane, N. Shibata, T. Yagi, T. Kubota and Y. Arakawa, 2004. Molecular characterization of a cephamycin-hydrolyzing and inhibitor-resistant class A β-lactamase, GES-4, possessing a single G170S substitution in the Ω-loop. Antimicrob. Agents Chemother., 48: 2905-2910.
CrossRef - Wachino, J.I., Y. Doi, K. Yamane, N. Shibata, T. Yagi, T. Kubota, H. Ito and Y. Arakawa, 2004. Nosocomial spread of ceftazidime-resistant Klebsiella pneumoniae strains producing a novel class A β-lactamase, GES-3, in a neonatal intensive care unit in Japan. Antimicrob. Agents Chemother., 48: 1960-1967.
CrossRef