
Yu. P. Petrov
Department of Cell Culture, Institute of Cytology, Russian Academy of Sciences, St. Petersburg, Russia
T.A. Krylova
Department of Cell Culture, Institute of Cytology, Russian Academy of Sciences, St. Petersburg, Russia
N.V. Tsupkina
Department of Cell Culture, Institute of Cytology, Russian Academy of Sciences, St. Petersburg, Russia
V.P. Pershina
Department of Cell Culture, Institute of Cytology, Russian Academy of Sciences, St. Petersburg, Russia
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 1 | Page No.: 102-112







ABSTRACT
Dynamics of cell spreading in the whole cell population is formed by dynamics of spreading of individual cells. In attempt to reveal regularities of spreading for the whole cell population we have measured Area, Perimeter and spreading-coefficient of the cells. Within 3 h upon plating of the cells on various substrates, cell spreading is a two-phase process, with retardation of spreading between 60 and 90 min after plating. The external factors (in particular, a type of substrate) do not influence on the character of cell spreading but affect only on the spreading rate. We suggest that these phenomena are based on such fundamental properties action as assembly and disassembly of filaments indicating a sharp transition from a globular to polymeric action state.
PDF Abstract XML References Citation
How to cite this article
Yu. P. Petrov, T.A. Krylova, N.V. Tsupkina and V.P. Pershina, 2007. Spreading as a General Attribute of Cell Population. Journal of Biological Sciences, 7: 102-112.
DOI: 10.3923/jbs.2007.102.112
URL: https://scialert.net/abstract/?doi=jbs.2007.102.112
DOI: 10.3923/jbs.2007.102.112
URL: https://scialert.net/abstract/?doi=jbs.2007.102.112
INTRODUCTION
The number of works on the properties and practical use of stem cells raises every year. Significance of these works for development of replacement therapy is evident. For this direction would receive a reliable substantiation for practical purposes, comprehensive experimental work is necessary. In most cases replacement therapy is based on availability of established lines of stem cells. This implies that properties of cell populations rather than those of individual cells should be studied (Dugina et al., 1998; Kapur and Rudolph, 1998; Deasy et al., 2003; Kharitonova and Vasiliev, 2004; Nelson et al., 2004).
Cells placed in a Petri dish begin to behave as independent association (population). Surveying cell culture from this point of view, it is reasonable to use such experimental tests that would characterize the whole population. Such characteristics of cell associations are, for example, growth curves, mitotic index, karyotype, etc.
To choose parameters for testing cell populations we have taken into account the following. The cell placed in culture conditions necessarily contacts the substrate. First of all, interaction of cell with substrate will be determined by mechanisms of adhesion and spreading (Orend and Chiquet-Ehrismann, 2000; Sastry and Burridge, 2000; Chen et al., 2003). It is possible to assert a priori that in terms of these characteristics cell population is heterogeneous. Hence for the given cell population these characteristics should be of stochastic nature. To formalize an extent of cell spreading, we have earlier suggested to measure the area of the substrate occupied by a cell during spreading (Area), the length of the cell circumference (Perimeter) cells and spreading coefficient (Rp/Ra).
Using these parameters in that work we have demonstrated specific heterogeneity of cell populations during spreading. However the data have been analysed only in one fixed time point per experiment. The aim of the present work was to follow dynamics of cell spreading within first hours upon reinoculation. To search for general patterns in behaviour of cells in culture, we used different cell lines (epitelioid and fibroblast-like cells, primary cultures and established cell lines).
MATERIALS AND METHODS
The following cells were used in this work: mouse embryonic fibroblast STO, human larynx epidermoid carcinoma-derived cells HEP-2, human epidermoid carcinoma-derived cells A-431 and the human mesenchymal cells. The established cell lines (STO, HEP-2, A-431) were obtained from the Cell Culture Collection (Institute of Cytology, Russian Academy of Sciences). The mesenchymal cells were obtained from marrow of human embryos (5-6 week of development).
All cells were grown in Petri dishes (35 mm) in DMEM supplemented with 10% calf serum, in a humidified atmosphere of 5% CO2 at 37°C. Concentration of cells at plating was 2-3x106 cell/dish. The cells were allowed to grow during 30, 60, 90, 120 and 180 min, then they were washed out with PBS (pH 7.4), fixed by methanol and stained with haematoxylin and eosin. The preparations were examined under light microscope with teleheader (Nikon Digital Camera COOLPIX4500, USA) at magnification of x200 (objective x20, eyepiece x10).
The culture medium in which the cells were grown up to a complete monolayer were used as a conditioned medium. After reaching stationary phase, the cells were removed from the substrate by incubation in 0.02% EDTA for 20-30 min at 37°C. In addition, the Petri dishes with the extracellular matrix preserved after the cells were removed were used. The matrigel has been kindly provided by Dr. L.V. Kukhareva. The matrigel solution (concentration 0,25 mg mL-1) was used to coat plates overnight at 4°C. Non-bound matrigel was removed by washing with DMEM and the plates were used for cell seeding.
The cell images were analyzed using ImageJ v.1.37 (Wayne Rasband, National Institute of Health, Maryland, USA). Each cell was outlined manually. The program counted perimeter and area of the every marked cell automatically. Both parameters were measured in pixels without transforming them into a standard metrical unit; this did not prevent comparison of relative values. At least 1000-2000 cells were analyzed per each dish.
RESULTS
Spreading curves: After re-seeding, cells should adapt to a new environment. Hence the lag phase precedes the process of an intensive spreading. Any real cell population is heterogeneous in terms of adhesive properties of the cells. Therefore all cells cannot begin spreading simultaneously. Besides, cells can not spread infinitely. This suggests the presence of an upper limit for Area. Taking this into account it is reasonable to assume that upon seeding the Area values should increase as a S-shaped curve and be described by the appropriate equation (for example, by Boltzmann equation).
However, in a real cell population the changes of Area deviated from a classical S-curve. The increase of Area for two populations of established cell lines is presented in Fig. 1. It is clearly seen that the presented data cannot be satisfactorily described by a typical S-curve. In this and other cases an additional plateau was revealed in the part of the curve where the logarithmic course of the S-curve should be expected. This part of the curve corresponds to the time interval of 90-120 min for STO cells and 60-90 min for HEP-2. In other cell lines studied the intermediate plateau was revealed to a greater or smaller extent.
We assume that the unusual behaviour of cell populations points to a two-phase process of spreading in which the every stage can be described by an independent S-curve.
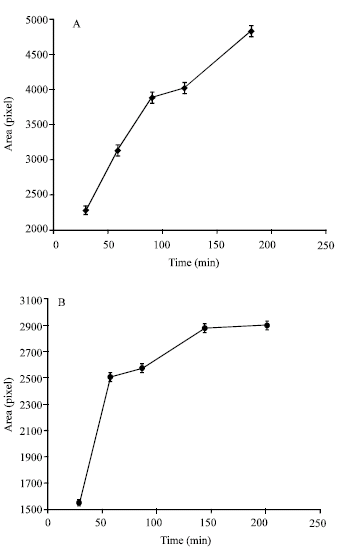 | |
| Fig. 1: | Dynamics of spreading of STO (A) and HEP-2 cells (B) on plastic. The area values are presented as mean±SE |
For example, if one would exclude the 180 min point (Fig. 1A) from the total sample the other values would be easily described by the S-curve (Fig. 2A). The second stage can be deduced in a similar way if one would exclude the 30 min point. Figure 2B shows this result for HEP-2.
Cell spreading always includes both phases, although one of them can be more pronounced than another. For example, when the Area of STO cells were measured in 3 h and later, the corresponding plateau was observed in 5-6 h of cell cultivation (data not presented). Unfortunately, it was not possible to register a lag phase for HEP-2 cells under these conditions because the cells passed the first phase of spreading very quickly.
Effect of environment on dynamics of cell spreading: Because of the increased interest to stem cells special attention has recently been drawn to cell micro environment and its active effect on the fate of the cells.
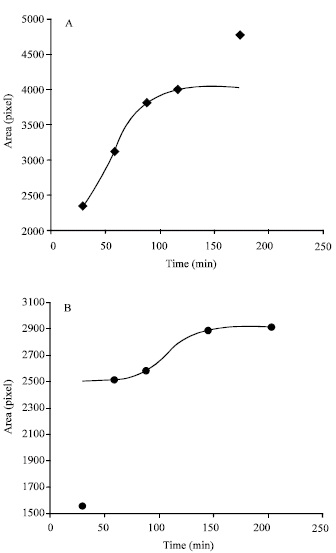 | |
| Fig. 2: | (A) - Reconstruction of the first phase of the spreading (STO cells). (B) - Reconstruction of the second phase of spreading (HEP-2 cells). Markers are the experimental values. A solid line is a theoretical S-curve. The large markers are the experimental points that were not used for fitting |
Many works have been performed to separate components of extracellular matrix (ECM), in particular collagens, laminins and fibronectins. It is believed that ECM proteins can play a main role in cell differentiation or transdifferentiation. In this study we elucidated general response of the cells on their surrounding. As the components potentially influencing cell spreading we used plastic (Petri dishes), matrigel (as a substance consisting of the major proteins of ECM), the own ECM of the cells and a conditioned medium.
Rp/Ra as the spreading-coefficient: Cell populations can be heterogeneous either because of the presence of differences of the Area stable in time (true cell subpopulations) or because of different types of the functional cell state (transient modifications of Area). In addition to parameter Area it is reasonable to use parameter Perimeter. Perimeter is not of a great importance if the cells have a rounded form as it is the case, for example, for epithelial cells. In this case, the S-curves for Area and Perimeter practically coincide. Importance of Perimeter as an independent parameter is increased if the cells have an expressed tendency to spreading. Then, the cells deviate from a correct circle to a great degree.
Although with spreading the Perimeter values grow faster than Area, both parameters are connected closely enough. To evaluate deviation of cell shape from a correct circle (polarization or stellation), we have earlier proposed to use Rp/Ra (Kuz'minykh and Petrov, 2004) that is called here spreading coefficient (Ks). Ks represents an interrelation between the radii calculated from experimental values of Perimeter and Area (Rp and Ra, respectively). For the clear reasons, the spreading coefficient cannot be less than 1. The coefficient grows up when the cell is either polarized or shapes some stellate form. Therefore Ks is less important at the initial stages of cell spreading and for epithelial cells resistantly retaining the rounded shape during cultivation.
Analysis of population of mesenchymal cells: Relevance of spreading parameters (Area, Perimeter and Rp/Ra) can be demonstrated for the cells that have tendency to polarize and form stellate shape. For this reason mesenchymal cells were chosen.
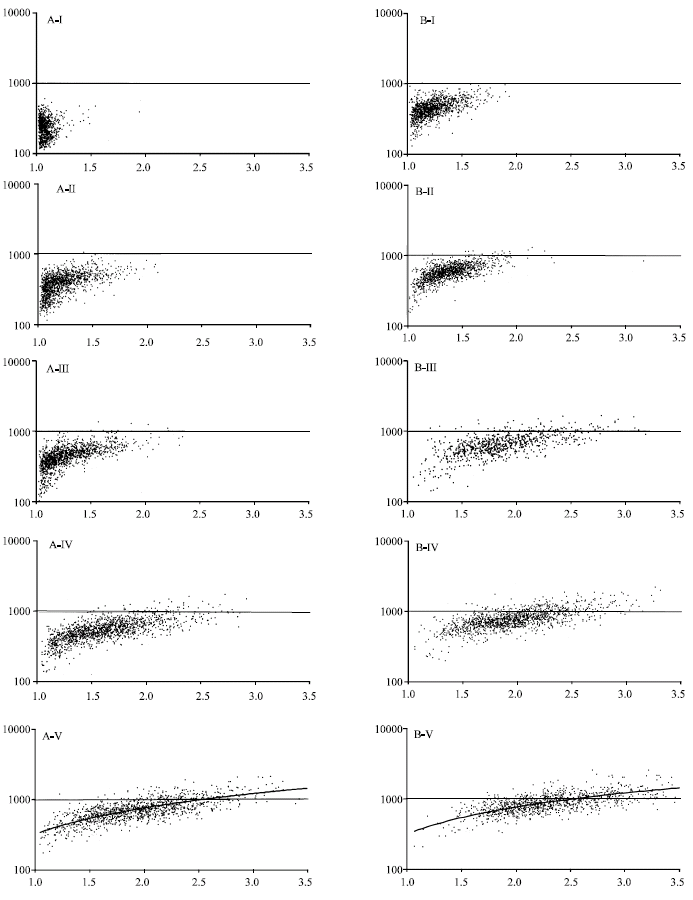
The data presented in Fig. 3 show that all three parameters of spreading can be relatively independent. The Area of mesenchymal cells differ from those presented above (Fig. 3A). No plateau was observed in time interval of 60-120 min. The differences in Area among the four cell variants can be observed up to 30 min. At a later time (to 180 min) all curves are converged approximately at the same level. This probably corresponds to the upper boundary of Area that cells of this type in this experiment cannot exceed. This upper boundary does not seem to depend either of environment or of substrate.
The results of the Perimeter measurements (Fig. 3A) in the first two points are qualitatively similar to those of the Area, that is, during the first hour both parameters increased proportionally. If the cells would remained rounded the Ks values (Fig. 3C) would be identical at 30 and 60 min. Here, we see that in all four variants polarization (or formation of the stellate form) took place. As the slopes of all the curves in this time interval are approximately identical we can suppose that dynamics of spreading for all the cells is nearly the same, too. Thereby, the cells have the largest efficiency of spreading on matrigel and the least one on plastic.
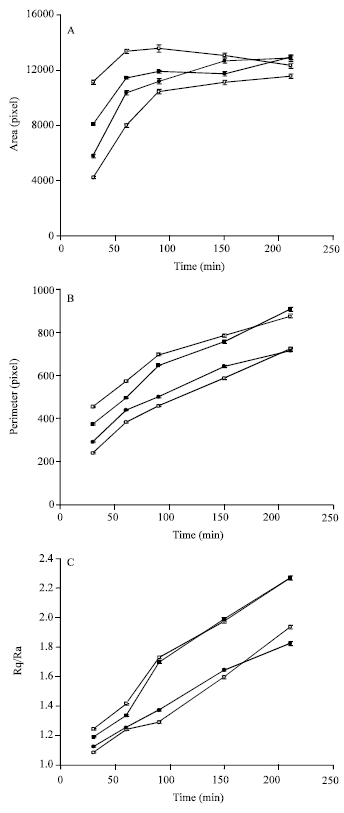 | |
| Fig. 3: | Spreading of mesenchymal cells on plastic (F), matrigel (~), own extracellular matrix (�) in a conditioned medium (M) Each point is the experimental value (mean±SE). (A)-Area, (B)- Perimeter, (C)-Rp/Ra |
In spite of the fact that the plateau typical for the cells presented above is not revealed in Fig. 3A the bent of the curves (especially on Fig. 3C) can be revealed in the same region (60-90 min). It could be noted that the cells on the matrigel and on their own ECM spread faster than on plastic or in a conditioned medium.
Interconnection of the character and dynamics of cell spreading: Our previous data (Kuz'minykh and Petrov, 2004) indicate that the cell population is heterogeneous in terms of Area. In the previous work we have suggested that the complicated profile of the curves is due either to the existence of stable functional states of the cells or to the presence in the cell culture of genuine subpopulations. To distinguish between these possibilities in the present work we used mesenchymal cells that are known to be heterogeneous in terms of genuine subpopulations. To reveal the groups of the cells we increased the amount of the cells per point up to 1000 and more. Moreover, only one dish was used in a single point to avoid randomization which could smooth boundaries between of cell groups (subpopulations).
Figure 4 and 5 present the data on spreading of mesenchymal cells on plastic and matrigel (as two extremes) and demonstrate as Area (or Perimeter) changes with increase of spreading coefficient. The every dot corresponds to an individual cell. It is seen that these plots do not display the expected clusters. Thus, in spite of the natural heterogeneity of mesenchymal cells no groups that differ by dynamics of spreading can be revealed.
The values of Area remained practically invariable with the increase of the spreading coefficient in all the cases, whereas the values of Perimeter grew gradually. The majority of the dots for the cells on plastic is shifted to the left and down, whereas on matrigel the majority of the dots is posed to the right and up. Nevertheless, the points are always assorted along the same trend and do not dependent of the plating conditions. This is well shown in plots reflecting the cell spreading up to 210 min. The solid lines are actually coinciding.
For Area this result (the coinciding regression lines) is not so surprising because the Area approaches the natural limit. For this reason the regression line goes almost in parallel to abscissa. However, for Perimeter this result seems to be quite unexpected. Why? First, many data in literature demonstrate different behavior of cell spreadings on different substrates. Second, even our data in Fig. 4 show that the different substrates produce the different effect on the average value of Perimeter. The difference of the average values is, however, a quantitative one, whereas the change of the dynamics is qualitative. If the cell spreading would depends on substrate qualitatively, the regression lines for plastic and matrigel should be different, too: they should either go at a different level or have a different slope. The data in Fig. 4 and 5 show that the substrate does not change the mechanism of cell spreading, but only influences its rate.
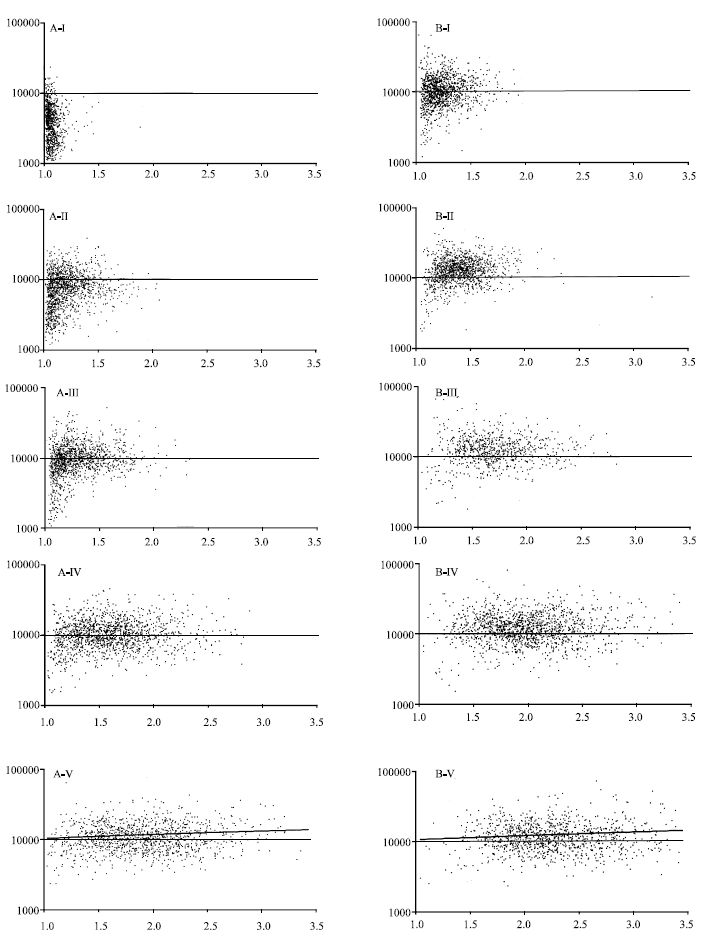 | |
| Fig. 4: | Changes in the Area of mesenchymal cells during spreading on plastic (A) and matrigel (B). The abscissa is values of spreading-coefficient (Rp/Ra). The ordinate is values of the Area, pixels (logarithmic scale). The curves of linear regression are shown only on the (e) plots. Scale is the same on all plots |
 | |
| Fig. 5: | Changes in the Perimeter of mesenchymal cells during spreading on plastic (A) and matrigel (B). The abscissa is values of spreading-coefficient (Rp/Ra). The ordinate is values of Perimeter, pixels (logarithmic scale). The curves of linear regression are shown only on the (e) plots. Scale is the same on all plots |
The nature of spreading is not determined by substrate: The data obtained for mesenchymal cells were confirmed on other cell types. The fragment of a similar experiment with A-431 cells is presented in Fig. 6. As an example, the data on Perimeter for 180 min are shown. A-431 cells have no such high capacity to spreading as mesenchymal cells (their spreading coefficient is 1.5-2). Nevertheless, the results of the Perimeter measurements for these cells revealed the distribution also described by the same equation of linear regression both for spreading on plastic and on the ECM of L-cells. At the same time, the average value of Perimeter of these cells on plastic is smaller than that on matrigel.
Again, the extracellular matrix influences only the rate of cell spreading. In addition, the spreading can depend on concentration of the cells at plating. The fragment of the experiment with STO cells presented in Fig. 7 shows that the higher is concentration of the cells the higher is the rate of spreading (dots are shifted to the right).
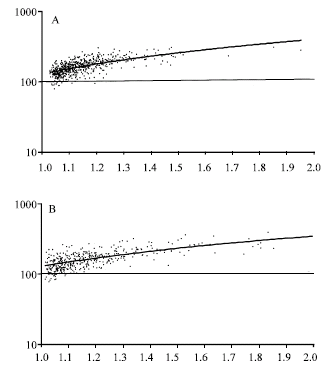 | |
| Fig. 6: | Comparison of spreading of A-431 cells for 180 min upon plating on plastic (A) and on ECM of L-cells (B). The abscissa is a spreading-coefficient (Rp/Ra). The ordinate is values of Perimeter, pixels (logarithmic scale) |
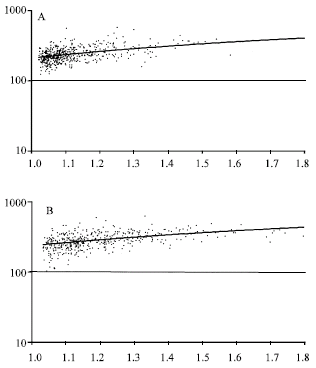 | |
| Fig. 7: | Effect of cell number at plating on perimeter of STO cells. Time of cultivation is 180 min. Concentration of cells at plating: (A)-400000, (B)-800000 cells/dish. The abscissa is spreading-coefficient (Rp/Ra). The ordinate is values of Perimeter, pixels (logarithmic scale) |
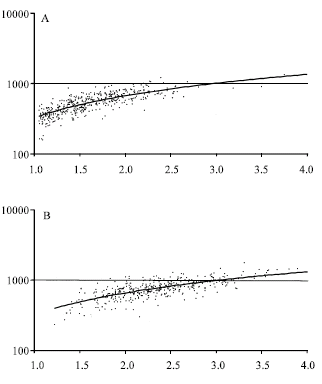 | |
| Fig. 8: | Effect of the cultivation time on perimeter of mesenchymal cells plated on plastic: (A) - 210 min, (B) - 1170 min. The abscissa is a spreading coefficient (Rp/Ra). The ordinate is values of Perimeter, pixels (logarithmic scale) |
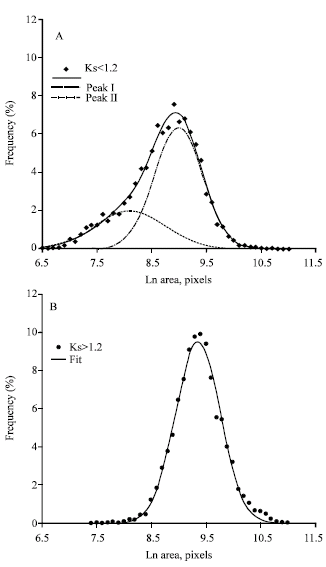 | |
| Fig. 9: | Proportion of rounded (A) and non-rounded (B) mesenchymal cells. All the data for plastic, matrigel, ECM, conditioned medium were used. The number of the cells on the every curve is about 4000. (A) - the cells at 30 min upon plating (the cells which a spreading coefficient is less than 1.2 are presented). (B) - the cells at 210 min upon plating (the cells which spreading coefficient is more than 1.2 are presented) |
In one of the experiments with mesenchymal cells we observed spreading within almost 24 h. But even after this time, no deviation from the regression line shown for the cells in 3,5 h upon plating was revealed (Fig. 8).
The shape and spreading of cells: The presented results indicate two facts: (1) spreading is a two-phase process (there is an intermediate plateau in S-curves of spreading, (Fig. 1 and 2); (2) an external factors cannot qualitatively change spreading of the cells (spreading of the same cells is described by the same equation independent of culture conditions (Fig. 4 to 8). To some extent both facts contradict each other. However it is not so.
As a rule, intermediate plateau in spreading curves was revealed at 30-60 min after the cell plating. A high dispersion of the points corresponding to rounded cells (Ks less 1.2) is observed in Fig. 4A, B and 5A, B. arising a question of whether rounded cells are homogeneous? To address this question we have analysed the data on mesenchymal cells grown at any substrate but sorted according to their spreading coefficient (Fig. 9).
The results of this analysis revealed a surprising homogeneity of non-rounded cells. They were grouped in one acute peak described by the normal distribution with a correlation coefficient of 0,996. On the contrary, the rounded cells are turned out to be heterogeneous. Two peaks (small and large cells) are clearly displayed. The common curve has a high correlation coefficient of 0.995, too. It is note worthy that the average values for the main big peak (the large rounded cells) of the plot in Fig. 9A and of the peak in the plot in Fig. 9B (non-rounded cells) belong to the same group with a linear dependence shown in Fig. 4 (B, V). It means that they belong to the same cell population with a common mechanism of spreading. On the other hand, the presence of two distinct peaks in the subpopulation of rounded cells point to a fundamental difference between small and large rounded cells.
DISCUSSION
The results of the presented work indicate that spreading of cultured cells can be a mirror reflecting a state of cell population. Statistical analysis of spreading behaviour revealed two relevant features of cultured cells that were not seen when individual cells were analyzed. We showed, for the first time, that spreading of cells on substrate is not a permanent process to be described by a standard S-curve. This process has two phases. Moreover, specificity of spreading is determined by the cell type whereas external factors (type and structure of substrate, concentration of cells at plating, time cultivation) affect only the spreading rate.
Although no special works on this subject were found in the literature, a similar phenomenon can be seen in a plot describing spreading of cells plated on colllagen (Clarke et al., 2004). In spite of the fact that the sample was small (up to 100 cells), an intermediate plateau can be revealed in the plots reflecting changes in the Area of the spreading cells within the same time of 60-120 min. The presence of the intermediate plateau on the S-curves of spreading cell tells about serious intracellular rearrangements that require time for switching from one state to another. The fact that this time is similar (about 1-1, 5 h after plating) for different cell lines points to a fundamental character of this phenomenon. We assume that the intermediate plateau on the spreading curves is due to the presence of two linked processes of cytoskeletal rearrangements. The interesting data on the changes of action cytoskeleton during first hour upon cell plating were obtained on fibroblasts (Zimerman at al., 2004). The authors have shown that small (of about 1 microns) dot-like structures with F-action core are broadly distributed over the ventral surfaces of the cells during cell spreading, being persisted for up to 30-60 min upon plating. At longer incubation, they were gradually replaced by stress fibers. We cannot be sure that these data and ours observations reflect the same intracellular events at early phases of spreading. However coincidence of the time intervals indicates that both phenomena can be tightly connected.
A two-phase spreading can reflect interaction of cell receptors with substrate and the subsequent reorganization of cytoskeleton (Zhou et al., 2000; Holly et al., 2000; Goldman, 2002; Hinz et al., 2003; Sepulveda et al., 2005). On the one hand, cytoskeleton reorganization determines formation of various structures and states of cells during their spreading on substrate (formation of lamella, filopodia, leading edge, cell polarization, etc.) (Menager et al., 1999; Kharitonova and Vasiliev, 2004; Chandrasekar et al., 2005). On the other hand, the presence, lack or redistribution of receptors on the cell surface, in particular integrins, determine how the cell will react with the certain substrate (Stupack, 2005).
Numerous data show dependence of cytoskeleton rearrangements on substrate (Cook at al., 1999; Vossmeyer et al., 2000; Khyrul et al., 2004). As a rule, these works present data on individual cells rather than on the cells of whole population. Therefore it is not always clear whether the same cells are compared in various time intervals or the authors show the different cells to illustrate the most typical examples of cytoskeleton patterns (Menager et al., 1999; Are at al., 2001; Chen et al., 2004; Sagnella et al., 2005). Attempts to formalize the cytoskeleton structures by classifying some stable states are subjective enough (Petukhova et al., 2004). Moreover, in the latter work, subjectivity is combined with incorrect statistical calculations.
The phenomenology of the influence of ECM proteins on cell shape resulted in an established idea of a possibility of their active and purposeful effect on a cell both in terms of cell phenotype and programs of differentiation (Hohn and Denker, 1994; Reh and Pittack, 1995; Deugnier et al., 1999; Gustafsson and Fässler, 2000). At present this approach prevail in the works with stem cells (Takito and Al-Awqati, 2004). It has played some role for working up the concept of a niche for a stem cell (Watt and Hogan, 2000; Campos, 2005). Our results show, however, that at spreading resistance of the cells to external factors is rather high. For example, the cells cannot be forced to change the spreading mode by changing quality of substrate. Some authors arrive at the same conclusion that cell adhesion is determined not by the proteins of an extracellular matrix (as external factor), but by the type of a cell line, by morpho-physiology state of cell population (Kurihara and Nagamune, 2005).
We have revealed that only two cell shapes, rounded and non-rounded, can be easily distinguished (Fig. 9). The two-phase nature of spreading is probably connected with the presence of the cells of both shapes. This is the superficial manifestation of spreading. It is more important, however, that the rounded cells are separated into two clear groups, one of which (large rounded cells) seems to have the same mechanism of spreading as the non-rounded cells (see the regression curves). We assume that some fundamental cell properties should determine this phenomenon being connected with basic properties of action. Assembly and disassembly of action filaments is a basis for spreading and locomotion of any cells (Carlier and Pantaloni, 1997; Pollard et al., 2000; dos Remedios et al., 2003). A key role at cell spreading may be ability to sharp transition from a globular to polymeric state.
The data presented in Fig. 9 indicate that transformation of the small rounded cells into the large rounded cells occurs step-wise. If our assumption is correct preparation for formation of cytoskeleton occurs in small rounded cells (action is mostly globular). Transformation of the small rounded cells into the large rounded cells implies a massive formation of action filaments.
ACKNOWLEDGMENTS
We thank Dr. S. Yu. Khaitlina and B.A. Margulis for the help and discussion.
REFERENCES
- Chandrasekar, T.E., B. Stradal, M.R. Holt, F. Entschladen, B.M. Jockusch and W.H. Ziegler, 2005. Vinculin acts as a sensor in lipid regulation of adhesion-site turnover. J. Cell Sci., 118: 1461-1472.
Direct Link - Chen, C.S., J.L. Alonso, E. Ostuni, G.M. Whitesides and D.E. Ingber, 2003. Cell shape provides global control of focal adhesion assembly. Biochem. Biophys. Res. Commun., 307: 355-361.
Direct Link - Chen, Y., D.J. Abraham, Xu Shi-wen, J.C.M. Black, K.M. Lyons and A. Leask, 2004. CCN2 (Connective Tissue Growth Factor) fibroblast adhesion to fibronectin. Mol. Biol. Cell, 15: 5635-5646.
Direct Link - Clarke, D.M., M.C. Brown, D.P. LaLonde and C.E. Turner, 2004. Phosphorylation of actopaxin regulates cell spreading and migration. J. Cell Biol., 166: 901-912.
Direct Link - Cook, G.A., D.A. Wilkinson, J.T. Crossno, Jr. R. Raghow and L.K. Jennings, 1999. The Tetraspanin CD9 influences the adhesion, spreading and pericellular fibronectin matrix assembly of chinese hamster ovary cells on human plasma fibronectin. Exp. Cell Res., 251: 356-371.
CrossRef - Deugnier, M.A., M.M. Faraldo, P. Rousselle, J.P. Thiery and M.A. Glukhova, 1999. Cell-extracellular matrix interactions and EGF are important regulators of the basal mammary epithelial cell phenotype. J. Cell Sci., 112: 1035-1044.
Direct Link - Dos Remedios, C.G., D. Chhabra, M. Kekic, I.V. Dedova, M. Tsubakihara, D.A. Berry and N.J. Nosworthy, 2003. Actin binding proteins: Regulation of cytoskeletal microfilaments. Physiol. Rev., 83: 433-473.
Direct Link - Goldman, W.H., 2002. Mechanical aspects of cell shape regulation and signaling. Cell Biol. Int., 26: 313-317.
CrossRef - Gustafsson, E. and R. F�ssler, 2000. Insights into extracellular matrix functions from mutant mouse models. Exp. Cell Res., 261: 52-68.
CrossRef - Hinz, B., V. Dugina, C. Ballestrem, B. Wehrle-Haller and C. Chaponnier, 2003. Smooth muscle actin is crucial for focal adhesion maturation in myofibroblasts. Mol. Biol. Cell, 14: 2508-2519.
Direct Link - Hohn, H.P. and H.W. Denker, 1994. The role of cell shape for differentiation of choriocarcinoma cells on extracellular matrix. Exp. Cell Res., 215: 40-50.
Direct Link - Kapur, R. and A.S. Rudolph, 1998. Cellular and cytoskeleton morphology and strength of adhesion of cells on self-assembled monolayers of organosilanes. Exp. Cell Res., 244: 275-285.
CrossRef - Kharitonova, M.A. and J.M. Vasiliev, 2004. Length control is determined by the pattern of cytoskeleton. J. Cell Sci., 117: 1955-1960.
Direct Link - Khyrul, W.A.K.M., D.P. LaLonde, M.C. Brown, H. Levinson and C.E. Turner, 2004. The integrin-linked kinase regulates cell morphology and motility in a Rho-associated kinase-dependent manner. J. Biol. Chem., 279: 54131-54139.
Direct Link - Kurihara, H. and T. Nagamune, 2005. Cell adhesion ability of artificial extracellular matrix proteins containing a long repetitive Arg-Gly-Asp seguence. J. Biosci. Bioeng., 100: 82-87.
Direct Link - Kuzminykh, E.V. and Y.P. Petrov, 2004. A simple model for the study of effects of the extracellular matrix on the cell morphology in vitro. Biochim. Biophys. Acta, 1671: 18-25.
PubMed - Menager, C., J. Vassy, C. Doliger, Y. Legrand and A. Karniguian, 1999. Subcellular localization of RhoA and ezrin at membrane ruffles of human endothelial cells: Differential role of collagen and fibronectin. Exp. Cell Res., 249: 221-230.
CrossRef - Nelson, C.M., D.M. Pirone, J.L. Tan and C.S. Chen, 2004. Vascular endothelial-cadherin regulates cytoskeletal tension, cell spreadingand focal adhesions by stimulating RhoA. Mol. Biol. Cell, 15: 2943-2953.
Direct Link - Orend, G. and R. Chiquet-Ehrismann, 2000. Adhesion modulation by antiadhesive molecules of the extracellular matrix. Exp. Cell Res., 261: 104-110.
Direct Link - Petukhova, O.A, L.V. Turoverova, I.V. Kropacheva and G.P. Pinaev, 2004. Morphological peculiarities of epidermoid carcinoma A431 cells spread on immobilized ligands. Tsitologia, 46: 5-15. (In Russia).
Direct Link - Pollard, T.D., L. Blanchoin and R.D. Mullins, 2000. Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Annu. Rev. Biophys. Biomol. Struct., 29: 545-576.
CrossRefDirect Link - Reh, T.A. and C. Pittack, 1995. Transdifferentiation and retinal regeneration. Sem. Cell Biol., 6: 137-142.
CrossRef - Sagnella, S., E. Anderson, N. Sanabria, R.E. Marchant and K. Kottke-Marchant, 2005. Human endothelial cell interaction with biomimetic surfactant polymers containing peptide ligands from the heparin binding domain of fibronectin. Tis. Eng., 11: 226-236.
CrossRefDirect Link - Sastry, S.K. and K. Burridge, 2000. Focal adhesions: A nexus for intracellular signaling and cytoskeletal dynamics. Exp. Cell Res., 261: 25-36.
Direct Link - Sepulveda, J.L., V. Gkretsi and C. Wu, 2005. Assembly and signaling of adhesion complexes. Curr. Top. Dev. Biol., 68: 183-225.
Direct Link - Stupack, D.G., 2005. Integrins as a distinct subtype of dependence receptors. Cell Death Differ., 12: 1021-1030.
CrossRefDirect Link - Takito, J. and Q. Al-Awqati, 2004. Conversion of ES cells to columnar epithelia by hensin and to squamous epithelia by laminin. J. Cell Biol., 166: 1093-1102.
Direct Link - Watt, F.M. and B.L. Hogan, 2000. Out of Eden: Stem cells and their niches. Sci., 287: 1427-1430.
Direct Link - Zimerman, B., T. Volberg and B. Geiger, 2004. Early molecular events in the assembly of the focal adhesion-stress fiber complex during fibroblast spreading. Cell Motil. Cytoskeleton, 58: 143-159.
Direct Link - Zhou, L., E.L. Cheng, P. Rege and B.Y.J.T. Yue, 2000. Signal transduction mediated by adhesion of human trabecular meshwork cells to extracellular matrix. Exp. Eye Res., 70: 457-465.
CrossRef