
T. Incharoen
Laboratory of Animal Science, Faculty of Agriculture, Kagawa University, Miki-cho, Kagawa-ken, 761-0795, Japan
O. Khambualai
Laboratory of Animal Science, Faculty of Agriculture, Kagawa University, Miki-cho, Kagawa-ken, 761-0795, Japan
K. Yamauchi
Laboratory of Animal Science, Faculty of Agriculture, Kagawa University, Miki-cho, Kagawa-ken, 761-0795, Japan
Asian Journal of Poultry Science
Year: 2009 | Volume: 3 | Issue: 2 | Page No.: 42-50







ABSTRACT
To investigate the effects of dietary combinations of zeolite, plant extract and vermiculite (ZEM®) on growth performance and histological intestinal changes, male Sanuki Cochin chickens were allotted to 3 groups of 12 birds each. They were fed ad libitum on a basal commercial mash diet with 0 (control), 1 and 2 g kg-1 dietary ZEM®. A starter diet (CP, 21%; ME, 3,000 kcal kg-1) was fed to chickens until 28 day of age, at which point a grower diet (CP, 18%; ME, 2,850 kcal kg-1) was fed until 70 day of age. Feed intake and body weight gain tended to increase with increasing levels of dietary ZEM® and feed efficiency tended to be highest in the 1 g kg-1 ZEM® group. Most intestinal villus height, villus area, cell area and cell mitosis numbers were better than those of the control group and the duodenal cell areas and cell mitosis numbers of all intestinal parts were greater (p<0.05) in the ZEM® groups than in the control group. Protuberated cells were observed on the villus tip of the ZEM® groups. These histological changes indicate that the intestinal villi and epithelial cells were hypertrophied by ZEM®. The present results suggest that chicken growth performance would be improved due to hypertrophied intestinal villi and epithelial cells after feeding ZEM® and that ZEM® is effective alimentation for chicken diets.
PDF Abstract XML References Citation
How to cite this article
T. Incharoen, O. Khambualai and K. Yamauchi, 2009. Performance and Histological Changes of the Intestinal Villi in Chickens Fed Dietary Natural Zeolite Including Plant Extract. Asian Journal of Poultry Science, 3: 42-50.
DOI: 10.3923/ajpsaj.2009.42.50
URL: https://scialert.net/abstract/?doi=ajpsaj.2009.42.50
DOI: 10.3923/ajpsaj.2009.42.50
URL: https://scialert.net/abstract/?doi=ajpsaj.2009.42.50
INTRODUCTION
Zeolites and vermiculite are frequently considered as supplements that reduce odour emission and enhance growth due to hypertrophied functions of intestinal villi (Khambualai et al., 2009). Zeolites are crystalline, hydrated aluminosilicates of alkali (e.g., Na+, K+) and alkaline (e.g., Mg+2, Ca+2) earth cations, having infinite three-dimensional structures (Ozturk et al., 1998). Vermiculite is a clay mineral, magnesium alumino-silicate, which has a high cation-exchanging capacity (Evangelou and Lumbanraja, 2002). Zeolite has been shown to be a beneficial feed additive that exhibits a strong preference for binding nitrogenous cations like NH4+ (Nakaue et al., 1981). Dietary zeolite has been shown to reduce the toxicity of litter ammonia (Gupta et al., 1997), as well as aflatoxins, which are contaminants in feed stuffs such as corn, wheat and soybeans (Miazzo et al., 2000). Some researchers found that adding zeolites to broiler diets improved body weight gain, feed conversion ratio and tibia ash content (Zhang and Hung 1992; Debeic, 1994). In a earlier study, Khambualai et al. (2009) reported that dietary natural zeolite including plant extract (ZEM®) showed hypertrophied intestinal villus height, villus area, cell area, cell mitosis and absorptive epithelial cells in Aigamo ducks, resulting in improved body weight gain. However, data on the effects of ZEM® on the growth performance and histological changes of the intestinal villi in chickens are lacking.
Therefore, the aim of this study was to investigate the effects of dietary ZEM® on the performance and morphological changes of the duodenum, jejunum and ileum of male Sanuki Cochin chickens. Histologically, the intestinal villus height, villus area, cell area and cell mitosis numbers in each intestinal segment of these birds were observed using a light microscope. Additionally, epithelial cell changes were compared using a scanning electron microscope.
MATERIALS AND METHODS
Birds, Housing and Diets
One-day-old male Sanuki Cochin chickens were obtained from the Animal Science Farm in Kagawa prefecture. They were housed in an electrically heated brooder and given access ad libitum to water and diet. At 7 day of age, all birds were weighed and randomly allotted to 3 groups of 12 birds each. Chickens were maintained in individual cages in an environmentally controlled room under a photo-period of 14 h of light. From July 2007 to September 2007, experiment was conducted at the Faculty of Agriculture, Kagawa University.
The basal diet was supplemented with commercial ZEM® (ZEM Association, Kanagawa Prefecture, Japan), which is composed of zeolite (70%), enzymes extracted from plants such as pineapple and papaya (20%) and vermiculite (10%), at 0 (control group), 1 and 2 g kg-1. A commercial starter mash diet was fed to chickens up to 28 day of age and then changed to a grower mash diet until 70 day of age (Table 1). All birds were allowed ad libitum access to water and feed throughout the experimental period. Feed intake and body weight were measured weekly. All animal management and sampling procedures were in accordance with the Guidelines and Rules for Animal Experiments of Kagawa University, Japan.
| Table 1: | Feed ingredients and chemical compositions of the basal diet |
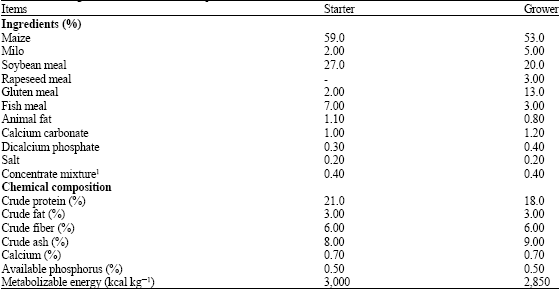 | |
| 1Concentrate mixture including (per kg of diet): vitamin A, 10,000 IU; vitamin D3, 2,000 IU; vitamin E, 15.63 mg; vitamin K, 2.6 mg; vitamin B1, 2.88 mg; vitamin B2, 3.7 mg; vitamin B6, 7.48 mg; vitamin B12, 0.01 mg; biotin, 0.14 mg; pantothenic acid, 16.12 mg; folic acid, 1.0 mg; niacin, 53.68 mg; choline, 1,400 mg; zinc, 75.00 mg; copper, 15.79 mg; manganese, 81.75 mg; iron, 170.76 mg; iodine, 0.35 mg; selenium, 0.15 mg | |
Small Intestine Sampling
At the end of the experimental period, 4 chickens form each treatment were chosen and killed by decapitation under light diethyl ether anesthesia. After that, the entire intestine was removed and placed immediately into a mixture of 3% glutaraldehyde and 4% paraformaldehyde fixative solution in 0.1 M cacodylate buffer (pH 7.4). The midpoint of the duodenum, the midpoint between the entry of the bile duct and Meckel’s diverticulum (jejunum) and the midpoint between Meckel’s diverticulum and the ileo-caecal junction (ileum) were cut and prepared for light and scanning electron microscopy.
Specimen Preparation for Light Microscope
The segment (2 to 3 cm in length) was washed with 0.1 M phosphate buffered saline (pH 7.4), fixed in Bouin’s solution overnight and dehydrated in a graded ethanol series. Each segment was embedded in paraffin wax using standard techniques. All samples were performed on 4 μm sections (8 sections/sample) and stained with haematoxylin and eosin and the resulting values were measured using an image analyzer (Nikon Cosmozone IzS, Nikon Co., Tokyo, Japan).
Measurement of Villus Height and Area
Villi were selected from each section (2 villi per section) that demonstrated the longitudinal view of a villus from base to tip. The villus height was measured from the tip to the bottom excluding the crypt, while the villus width was measured at the basal and apical parts. The average villus height from 4 birds (16 villi from 8 different sections in each segment per bird) was expressed as a mean villus height for one group. The value of villus area was calculated from the villus height, basal width and apical width according to the method described by Iji et al. (2001). The average villus area from 4 birds (16 calculations of villus area from 8 different sections in each segment per bird) was expressed as a mean villus area for one group.
Measurement of Cell Area
Villi were randomly selected from each section (2 villi per section). The villus area was measured at the middle part of the epithelial cell layer and then the cell nuclei within the cell layer were counted. Next, the area of the epithelial cell layer was divided by the number of cell nuclei. The average cell area from 4 birds (8 calculations of cell area from 4 different sections in each segment per bird) was expressed as a mean cell area for one treatment group.
Measurement of Cell Mitosis
Mitoses were identified as darkly stained basophilic nuclei with haematoxylin. The number of mitoses within the entire crypt was counted from 4 different sections per bird. The average of these values was expressed as a mean cell mitosis number for each bird. These 4 mean cell mitosis numbers from 4 birds were expressed as a mean cell mitosis number for one group.
Scanning Electron Microscope
The intestinal segment (length, 2 cm) was cut longitudinally, opened and washed with 0.1 M phosphate buffered saline (pH 7.4). Each segment was placed in a mixture of 3% glutaraldehyde and 4% paraformaldehyde in 0.1 mol L-1 cacodylate buffer (pH 7.4) at room temperature for 1 h. Afterward, the segment was cut into 5x6 mm squares and fixed in the same solution for a further 1 h. The specimens were stored in 0.1 mol L-1 sodium cacodylate buffer (pH 7.4) and post-fixed with 1% osmium tetroxide in an ice-cold buffer for 2 h. Next, these specimens were rinsed with deionized distilled water and kept in 70% ethanol. After freeze-drying, the dried samples were mounted on aluminium stubs with electrically conducting carbon paste, coated (E-1030 ion sputter, Hitachi Ltd., Tokyo, Japan) and the epithelial cells on the villus tip surface observed under a scanning electron microscope (Hitachi S-4300SE/N, Hitachi Ltd., Tokyo, Japan) at 8 kV.
Statistical Analysis
Data on growth performance and light microscopic examination were subjected to statistical analysis by one-way analysis of variance (ANOVA) of SPSS statistical software package (version 10.0 for windows, SPSS, Inc., Chicago, IL). The means of different treatments were compared with Duncan’s multiple range tests. Significance was considered at p<0.05 level.
RESULTS
Growth Performance
Feed intake, body weight gain and feed efficiency did not show a significant difference among groups (Table 2). However, they tended to increase with increasing levels of dietary ZEM®, with feed efficiency highest in the 1 g kg-1 ZEM® group.
Light Microscopic Observations
The intestinal villus height and villus area of all segments did not vary among groups (Fig. 1A-D). The cell area and cell mitosis numbers of all intestinal parts tended to slightly increase with increasing levels of ZEM® and the cell area of the duodenum and cell mitosis numbers of the all intestinal parts were greater (p<0.05) in the ZEM® groups than in the control group.
Morphological Changes of the Epithelial Cells
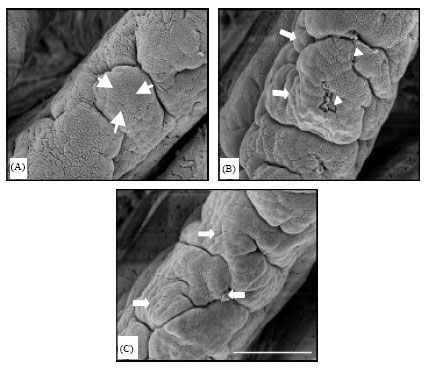
On the duodenal villus apical surface of the control, flat (small arrows) and protuberant cells (large arrows) were seen around the central sulcus (Fig. 2A). In the 1 g kg-1 ZEM® group (Fig. 2B), in addition to protuberating cells (large arrows), some cells devoid of any microvilli (arrowheads) were found among intact cells that were totally covered with microvilli. In the 2 g kg-1 ZEM® group (Fig. 2C), clearly protuberated cells (large arrows) and cell clusters (stars) composed of many epithelial cells were visible.
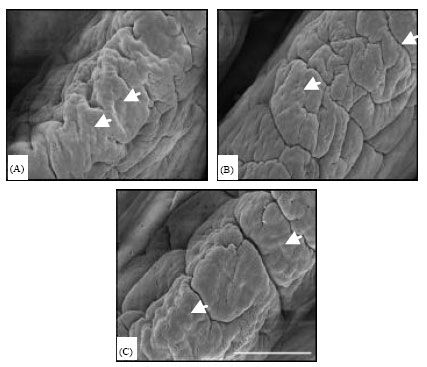
The jejunal villus apical surface of the control was covered by flat cell areas (small arrows) (Fig. 3A). In the experimental groups (Fig. 3B, C), such flat cells developed into faintly protuberated cells (large arrows). In the 1 g kg-1 ZEM group, areas having cells with no microvilli and deeper cells at sites of recently exfoliated cells (arrowheads) were observed in the exfoliative zone of the villus tip.
| Table 2: | Effects of dietary natural zeolite including plant extract on feed intake, body weight gain and feed efficiency in chickens (n = 4, Mean±SE) |
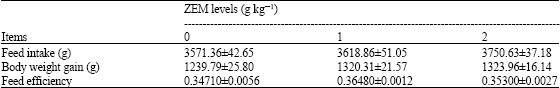 | |
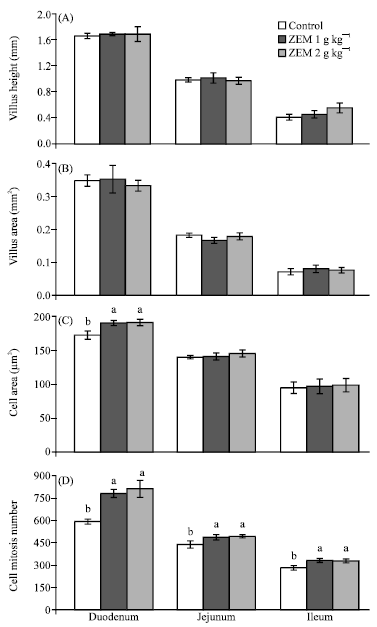 | |
| Fig. 1: | (A-D) Intestinal villus height, villus area, cell area and cell mitosis number of duodenal, jejunal and ileal segments in chickens fed a basal commercial mash diet with 0 (control), 1 and 2 g kg -1dietary ZEM. (Mean±SE, n = 4). Mean with different letters significantly differ (p<0.05) from each other |
On the ileal villus tip surface, faintly protuberating cells (small arrow) were observed in the control (Fig. 4A) and the ZEM groups (Fig. 4B, C). In addition, many intestinal sulci were observed.
DISCUSSION
In the present study, feed intake, body weight gain and feed efficiency were greater with increasing levels of a mixture of zeolite and plant extract (protease). Total weight gain was improved in the 1 and 2 g kg-1 ZEM® groups by approximately 6.49 and 6.79%, respectively, as compared to the control. Dietary zeolite improved body weight gain, feed conversion ratio and tibia ash content (Zhang and Hung, 1992; Debeic, 1994) and feed efficiency (Elliot and Edwards Jr., 1991).
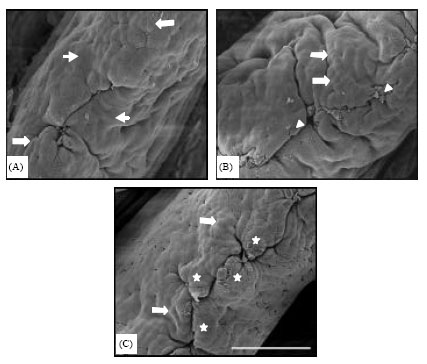 | |
| Fig. 2: | Duodenal villus apical surface in chickens fed fed a basal commercial mash diet with 0 (A; small arrow, flat cell; large arrows, protuberant cell), 1 (B; large arrows, protuberant cell; arrowheads, exfoliated cells) and 2 g kg-1 dietary ZEM (C; large arrows, protuberant cell, star, cell cluster). Scale bar = 50 µm (x600) |
 | |
| Fig.3: | Jejunal villus apical surface in chickens fed fed a basal commercial mash diet with 0 (A; small arrow, flat cell area), 1 (B; large arrows, protuberant cell; arrowheads, exfoliated cells) and 2 g kg-11 dietary ZEM (C; large arrows, protuberant cell). Scale bar = 50 µm (x600) |
 | |
| Fig. 4: | Ileal villus apical surface in chickens fed a basal commercial mash diet with 0 (A; small arrow, flat protuberating cell), 1 (B; small arrow, flat protuberating cell) and 2 g kg-1 dietary ZEM (C; small arrow, flat protuberating cell). Scale bar = 50 μm (x600) |
Also, dietary natural zeolite improved weight gain and feed efficiency in cattle (Koknaroglu et al., 2006). The addition of natural zeolite at 25 g kg-1 to the aflatoxin-containing diet also provided a slight improvement in body weight gain. In addition, many enzymes, such as protease, induced body weight gain and feed conversion (Cowieson and Adeola, 2005), due to improvements in total protein digestibility (Zanella et al., 1999). These reports suggest that the present slightly better growth performance might be the result of zeolite and plant extract. This is in agreement with the findings that dietary ZEM® (0.1, 0.5 and 1.0 g kg-1) improved weight gain in Aigamo ducks (Khambualai et al., 2009).
In these ZEM® groups, most light microscopic parameters were higher than those of the controls and the duodenal cell area and the cell mitosis numbers in all intestinal parts were increased. Most of the digestive and absorptive processes of ingested feeds occur in the intestine. It has been reported that intestinal villus and epithelial cell morphology is associated with intestine function and growth rate (Ruttanavut et al., 2009). Large-sized villi have been associated with activated cell proliferation (Lauronen et al., 1998). Narrow and long villi indicate a faster multiplication of the base of the crypt which migrated faster to the tip of villi and that the turnover of the epithelial cells would, therefore, be shorter (Nordstrom and Dahlqvist, 1973). Greater villus height and numerous mitotic cells in the intestine are indicators that the function of the intestinal villi is activated (Langhout et al., 1999; Yasar and Forbes, 1999). Furthermore, the present epithelial cells on the villus apical surface were protuberated in the duodenum and jejunum of the ZEM® groups. Areas having cells with no microvilli and deeper cells at sites of recently exfoliated cells were also observed on the duodenal and jejunal villus surface in the 1 g kg-1 ZEM group. Absorptive epithelial cells originate at the base of the crypts as immature proliferative cells, differentiate as they migrate up the villus tip and are finally extruded from the villous tips (Cheng and Leblond, 1974). The cell cycle of intestinal absorptive cells from origin to shedding at the villus tip is 2 to 3 days (Leblond, 1981). Thus, that the epithelial cells seem to show dramatic morphological changes at the exfoliative zone might be due to the villus function. Numerous epithelial cells were observed on the villus apical surface in chickens with increased body weight (Yamauchi et al., 2006) and cell protuberance into the intestinal lumen has been demonstrated to show hypertrophied cells (Shamoto and Yamauchi, 2000; Tarachai and Yamauchi, 2000). These histological reports suggest that the present intestinal villi and epithelial cells can be considered hypertrophied.
Wood charcoal could induce the hypertrophied villi and epithelial cells due to its many pores adsorption in chickens (Ruttanavut et al., 2009) and pigs (Mekbungwan et al., 2008). Zeolite also has many framework structures (Coombs et al., 1997) and is able to absorb water and exchange nitrogen molecules (Mumpton, 1999). Dietary zeolite reduced the toxicity of ammonia in the litter (Gupta et al., 1997) and aflatoxins in corn, wheat and soybeans (Miazzo et al., 2000). Exogenous enzymes, such as carbohydrases, proteases and phytases, have been used in poultry to improve growth performance (Bi and Chung, 2004; Kidd et al., 2001) and nutrient digestibility (Cowieson and Adeola, 2005) and to reduce the excretion of phosphorus (Maguire et al., 2004). These functions of zeolite and enzymes might induce hypertrophied intestinal villi and epithelial cells.
In conclusion, as ZEM could improve the growth performance of chickens due to the hypertrophy of the intestinal villi and epithelial cells, ZEM is an effective alimentation for chicken diets.
REFERENCES
- Bi, Y. and T.K. Chung, 2004. Effects of multiple-Enzyme mixtures on growth performance of broilers fed corn-soybean meal diets. J. Applied Poult. Res., 13: 178-182.
CrossRefDirect Link - Cheng, H. and C. P. Leblond, 1974. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine V. Unitarian theory of the origin of the four epithelial cell types. Am. J. Anatomy, 141: 537-561.
CrossRefPubMedDirect Link - Cowieson, A.J. and O. Adeola, 2005. Carbohydrases, protease and phytase have an additive beneficial effect in nutritionally marginal diets for broiler chicks. Poult. Sci., 84: 1860-1867.
CrossRefPubMedDirect Link - Debeic, M., 1994. Influence of clinoptilolites on chicken growth. Poult. Abstr., 21: 309-309.
Direct Link - Elliot, M.A. and H.M. Edwards Jr., 1991. Comparison of the effects of synthetic and natural zeolite on laying hen and broiler chicken performance. Poult. Sci., 70: 2115-2130.
PubMed - Evangelou, V.P. and J. Lumbanraja, 2002. Ammonium-potassium-calcium exchange on vermiculite and hydroxyl-aluminum vermiculite. Soil Sci. Soc. Am. J., 66: 445-455.
Direct Link - Mumpton, F.A., 1999. La roca magica: Uses of natural zeolites in agriculture and industry. Proc. Natl. Acad. Sci. USA., 7: 3463-3470.
CrossRefDirect Link - Gupta, G., J. Borowiec and J. Okoh, 1997. Toxicity identification of poultry litter aqueous leachate. Poult. Sci., 76: 1364-1367.
PubMedDirect Link - Iji, P.A., A. Saki and D.R. Tivey, 2001. Body and intestinal growth of broiler chicks on a commercial starter diet. 1. Intestinal weight and mucosal development. Br. Poult. Sci., 42: 505-513.
CrossRefDirect Link - Khambualai, O., J. Ruttanavut, M. Kitabatake, H. Goto, T. Erikawa and K. Yamauchi, 2009. Effects of dietary natural zeolite including plant extract on growth performance and intestinal histology in Aigamo ducks. Br. Poult. Sci., 50: 123-130.
CrossRefPubMedDirect Link - Kidd, M.T., G.W. Morgan and C.J. Price, 2001. Enzyme supplementation to corn and soybean meal diets for broilers. J. Applied Poult. Res., 10: 65-70.
Direct Link - Koknaroglu, H., M.T. Toker and Y. Bozkurt, 2006. Effect of zeolite and initial weight on feedlot performance of brown Swiss cattle. Asian J. Anim. Vet. Adv., 1: 49-54.
CrossRefDirect Link - Langhout, D.J., J.B. Schutte, P. van Leeuwen, J. Wiebenga and S. Tamminga, 1999. Effect of dietary high-and low-methyllated citrus pectin on the activity of the ileal microflora and morphology of the small intestinal wall of broiler chickens. Br. Poult. Sci., 40: 340-347.
CrossRefPubMedDirect Link - Lauronen, J., M.P. Pakarinen, P. Kuusanmakai, E. Savilahti, P. Vento, T. Paavonen and J. Halttunen, 1998. Intestinal adaptation after massive proximal small-bowel resection in the pig. Scand. J. Gastroenterol., 33: 152-158.
CrossRefPubMedDirect Link - Maguire, R.O., J.T. Sims, W.W. Saylor, B.L. Turner, R. Angel and T.J. Applegate, 2004. Influence of phytase addition to poultry diets on phosphorus forms and solubility in litters and amended soils. J. Environ. Qual., 33: 2306-2316.
PubMedDirect Link - Mekbungwan, A., K. Yamauchi, T. Sakaida and T. Buwjoom, 2008. Effects of a charcoal powder-wood vinegar compound solution in piglets for raw pigeon pea seed meal. Animal, 2: 366-374.
CrossRef - Miazzo, R., C.A. Rosa, E.C. De Queiroz Carvalho, C. Magnoli and S.M. Chiacchiera et al., 2000. Efficacy of synthetic zeolite to reduce the toxicity of aflatoxin in broiler chicks. Poult. Sci., 79: 1-6.
PubMedDirect Link - Nordstrom, C. and A. Dahlqvist, 1973. Quantitative distribution of some enzymes along the villi and crypts of human small intestine. Scand. J. Gastroenterol., 8: 406-416.
PubMed - Ozturk, E., G. Erener and M. Sarica, 1998. Influence of natural zeolite on performance of laying hens and egg quality. Turk J. Agric. For., 22: 623-628.
Direct Link - Ruttanavut, J., K. Yamauchi, H. Goto and T. Erikawa, 2009. Effects of dietary bamboo charcoal powder including vinegar liquid on growth performance and histological intestinal change in aigamo ducks. Int. J. Poult. Sci., 8: 229-236.
CrossRefDirect Link - Shamoto, K. and K. Yamauchi, 2000. Recovery responses of chick intestinal villus morphology to different refeeding procedures. Poult. Sci., 79: 718-723.
PubMedDirect Link - Tarachai, P. and K. Yamauchi, 2000. Effects of luminal nutrient absorption, intraluminal physical stimulation and intravenous parenteral alimentation on the recovery responses of duodenal villus morphology following feed withdrawal in chickens. Poult. Sci., 79: 1578-1585.
CrossRefPubMedDirect Link - Yamauchi, K., T. Buwjoom, K. Koge and T. Ebashi, 2006. Histological intestinal recovery in chickens refed dietary sugar cane extract. Poult. Sci., 85: 645-651.
PubMedDirect Link - Yasar, S. and J. M. Forbes, 1999. Performance and gastrointestinal response of broiler chicks fed on cereal gain-based foods soaked in water. Br. Poult. Sci., 40: 65-76.
PubMedDirect Link - Zanella, I., N.K. Sakomura, F.G. Silversides, A. Fiqueirdo and M. Pack, 1999. Effects of enzyme supplementation of broiler diets based on corn and soyabeans. Poult. Sci., 78: 561-568.
PubMed - Coombs, D.S., A. Alberti, T. Armbruster, G. Artioli and C. Colella et al., 1997. Recommended nomenclature for zeolite minerals: Report of the subcommittee on zeolites of the international mineralogical association, commission on new minerals and mineral names. Can. Mineral., 35: 1571-1606.
Direct Link