
M. Khalily
Payame Noor University, P.O. Box 438, Mahabad, Iran
LiveDNA: 98.3105
M. Moghaddam
Department of Agronomy, Faculty of Agriculture, Tabriz University, Tabriz, Iran
H. Kanouni
Research Center for Agriculture and Natural Resources of Kurdistan, P.O. Box 714, Sanandaj, Iran
LiveDNA: 98.744
ORCID: 0000-0002-4386-578X
E. Asheri
Payame Noor University, P.O. Box 438, Mahabad, Iran
Asian Journal of Crop Science
Year: 2010 | Volume: 2 | Issue: 2 | Page No.: 60-69







ABSTRACT
The major objective of this study was to assess grain yield and its components in some genotypes of maize under different levels of drought stress. Therefore, eight medium maturity hybrids of maize were evaluated using a split-plot arrangement in randomized complete block design with four replications in three stress (the vegetative, reproductive and grain filling phases) and normal conditions, at the Agricultural Research station of Miando'ab, West Azerbaijan, North West of Iran. Some phenological and morphological characteristics were recorded during cropping season. Analysis of variance and mean comparisons revealed that yield loss due to drought stress at vegetative growth phase recorded highest role in crop yield reduction comparing to other stress media. The analysis of variability parameters revealed presence of substantial variability for all traits. The phenotypic correlation coefficients unfolded positive and significant associations of grain yield with 300-kernel weight, kernel girth, kernel rows/ear and kernels/row, at normal conditions. The maturity traits recorded significant and negative correlation with yield, at stress prone conditions. The path analysis revealed that the most important direct effects on grain yield are number of kernels/row, ear length, ear girth and kernel rows/ear thus, should be used as target traits for tailoring an ideal plant type for higher yield of maize exposed to drought stress at vegetative period. According to the results, at drought condition, anthesis to silking interval, days to silk emergence, green cover percentage and much more kernel number in row are major traits which recommended for selecting drought tolerant hybrid varieties of corn.
PDF Abstract XML References Citation
How to cite this article
M. Khalily, M. Moghaddam, H. Kanouni and E. Asheri, 2010. Dissection of Drought Stress as a Grain Production Constraint of Maize in Iran. Asian Journal of Crop Science, 2: 60-69.
DOI: 10.3923/ajcs.2010.60.69
URL: https://scialert.net/abstract/?doi=ajcs.2010.60.69
DOI: 10.3923/ajcs.2010.60.69
URL: https://scialert.net/abstract/?doi=ajcs.2010.60.69
INTRODUCTION
Corn is currently the third most important crop in the world in terms of cash receipts and was grown on about 129 million hectares worldwide in 2004. Average productivity of this crop is 6.7 t ha-1 in developed countries and 2.4 t ha-1 in developing countries, including Iran (Food and Agriculture Organization of the United Nations, 2005).
Drought is a major constraint to increase yield in crop plants. Improving drought tolerance is therefore of outmost importance for breeding. However, the complexity of the trait allowed only marginal progress. Genetic improvement of adaptation to drought is addressed through the conventional approach by selecting for yield and its stability over locations and years. Such selection programs are slow in attaining progress because of the low heritability of yield under stress, the inherent variation in the field and the limitation that there is usually only one experimentally droughted crop per year (Tollenaar et al., 2004).
Alternatively, yield improvements in water-limited environments could be achieved by identifying secondary traits contributing to drought resistance and selecting for those traits in a breeding program. The effectiveness of selection for secondary traits to improve yield under water-limiting conditions has been demonstrated in maize (Zea mays L.) (Venugopal et al., 2003), wheat (Triticum aestivum L.) (Moghaddam et al., 1997; Richards et al., 2002) and sorghum (Sorghum bicolor L. Moench) (Galeta et al., 2005).
A number of studies in maize have been conducted to elucidate the nature of association between yield and its components which identify traits like ear length, ear diameter, kernel /row, ears/plant, 100-kernel weight and rows/ear as potential selection criteria in breeding program aiming at higher yield (Najeeb et al., 2009; Tollenaar et al., 2004). Estimation of simple correlation between various agronomic characters may provide good information necessary for maize breeders, when selection is based on two or more traits simultaneously (Singh and Singh, 1993). Information obtained from correlation coefficients for these characters could also be useful as indicators of the more important ones under consideration. The association among traits may be measured by genotypic and/or phenotypic coefficients correlation depending on the types of studied materials and the kind of experimental design used (Agrama, 1996).
Since drought resistance is a quantitatively inherited performance based trait, selection needs to be practiced with advanced generation lines in replicated trials over years and locations (Denmead and Shaw, 1960). Drought resistance can only be estimated by comparing the performance of breeding lines under stress and non-stress (irrigated) conditions. Using data from the two water treatments, breeders can calculate drought intensity index for the experiment and the different susceptibility indices and means to assist in selection of drought resistant genotypes (Abdmishani et al., 2004).
Our intention from this investigation was to study performance of grain yield and its components of eight maize hybrids in four different levels of soil moisture contents as well as, to assess the association between traits and path analysis of grain yield at those conditions.
MATERIALS AND METHODS
Site Characteristics
This study was carried out during Spring and Summer of 2006 at the Miando'ab Agricultural Research Station, 46°09' N, 36°58' E and 1371 m altitude, in the province of West Azerbaijan, North West of Iran. An open pan evaporation class A was used to avoid of side effects of climatically parameters on the irrigation periods. The pan was installed in a 15 m distance apart from the experiment. Each period of irrigation in the non-stressed treatments was conducted after 90±5 mm evaporation of the pan, which was approximately equal to 10 days evaporation during June-July and for stressed plots in a 20 days intervals equal to 180±5 mm open pan evaporation during different growth phases.
Experimental Design
A split plot arrangement in randomized complete block design with four main plots, different levels of irrigation, including complete irrigation, stress at vegetative phase, stress at reproductive phase and stress at grain filling period and eight sub plot, consist of eight medium maturity hybrids of corn (SC 604, SC 647, SC 686, SC 687, SC 688, SC 689, SC 690 and SC 691), with three replications was performed.
Data Collected
Each hybrid was raised in two rows of 5 m length with a spacing of 60 cm between the rows and 20 cm between plants within the rows. Recommended package of practices were followed to raise the crop and normal cultural practices were followed through out the crop growth. Five plants were randomly selected from each hybrid, replication-wise for recording observations on fifteen characters, viz., days to silk emergence, plant height, ear length, number of rows per ear, number of kernels per row, number of branches per tassel, total number of leaves, number of leaves per ear, number of dry leaves, kernel depth, green cover percentage, leaf rolling, 300 kernels weight, single plant yield and Anthesis to Silking Interval (ASI).
Data Analysis
Standard statistical procedures were used for the analysis of variance and mean comparisons of the selected cultivars were conducted using Fisher’s LSD. The phenotypic correlation coefficients for yield and yield components were evaluated using the formula suggested by Jha et al. (1998). Further, the path coefficient analysis which measure the direct influence of one variable upon another and permits the separation of the simple correlation coefficient into components of direct and indirect effects was done according to Wright (1921) and Dewey and Lu (1959).
RESULTS AND DISCUSSION
The results of analysis of variance showed highly significant differences for most of the traits (Table 1). Effects of drought stress at different stages of growth, on traits such as 300 kernels weight, ASI, number of kernels per row, number of branches of tassel, green cover percentage and grain yield; and on characters like number of kernel rows, kernel depth, plant height and ear length, were significant at 5 and 1% levels, respectively. Therefore, the performance of aforementioned traits in destined hybrids under drought stress treatments (vegetative, reproductive and grain filling periods) toward normal condition has changed. There were significant differences between hybrids regarding to 300 kernels weight, ASI, number of kernel rows, number of branches of tassel, number of kernel rows, green cover percentage and grain yield at 1% significant level and also for number of days to silk emergence, ear length and number of top leaves at 5% level, showing presence of variation for these traits among hybrids.
| Table 1: | Phenotypic variation in different characters of maize hybrids |
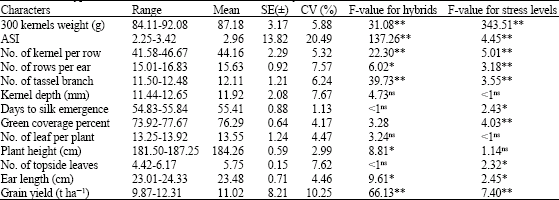 | |
| ns: Not significant; *: Significant at p = 0.05; **: Significant at p = 0.01 | |
| Table 2: | Performance of each character of maize hybrids in normal situation and different levels of drought stress |
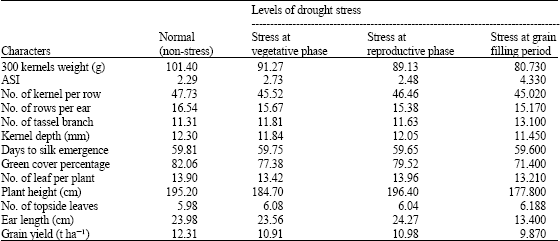 | |
Number of days to silk emergence and ASI had lowest and highest coefficients of variation, respectively. In general, results of ANOVA showed that there were a dramatically genetic variation among investigated hybrids and it could be exploited to select genotypes for drought prone environments.
Mean grain yield among the trials ranges from 9.9 to 12.3 t ha-1 (Table 1, 2). Interaction of hybridxstress levels was significant for ASI and ear length (data not shown). Overall mean of different characters for each single cross have showed in Table 3. Hybrid number six (SC 686) produced higher grain yield and lowest ASI (2.25 days). This genotype was superior over others regarding to some traits such as 300 kernel weight, number of kernel per row, number of rows per ear, kernel depth, green cover percentage and ear length. Hybrid number five (SC 687), showed highest value of ASI (3.42 days) and lowest number of kernels per row (41.58). Westgate (1994) and Edmeades et al. (1994) reported negative correlation between ASI and number of kernels per row. However, with ASI duration more than 8 days, number of kernels per row will be decrease significantly to produce even seedless ears.
Simple correlation coefficients among traits at different moisture contents were shown in Table 4-7. The coefficients of correlation for grain yield vs. 300 kernels weight, green cover percentage, number of kernel per row, leaf number and plant height were 0.63, 0.77, 0.57, 0.54 and 0.42, respectively and significant. Whereas correlation for grain yield vs. ASI and number of branches of tassel were negative and significant (Table 4). Bolanos and Edmeades (1996) found positive and significant coefficients of correlations between grain yield vs. number of kernel per row and one-hundred kernels weight at drought stress condition.
In the present study, correlation between grain yield vs. 300 kernels weight (0.51), number of kernel per row (0.72), kernel depth (0.54) and green cover percentage (0.41) were positive and significant at vegetative phase stress (Table 5). Also, correlation coefficient between grain yield and ASI was negative (-0.53) and significant (p = 0.05). Abrecht and Carberry (1993) in their study on effect of drought stress on different properties of maize reported that, moisture stress at vegetative phase increases association between grain yield and ASI.
Correlation coefficients between grain yield against 300 kernels weight (0.63), number of kernels per row (0.66), plant height (0.63) and ear length (0.41) were significant. Correlation between grain yield and ASI (-0.84), as well as grain yield and number of branches of tassel (-0.81) were significant (p = 0.01).
| Table 3: | Response of maize hybrids |
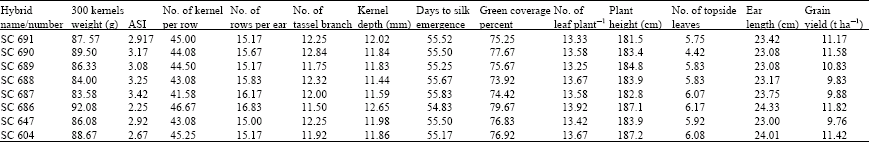 | |
| Table 4: | Phenotypic correlation coefficients between grain yield and some yield components and growth characters of maize hybrids under drought stress at normal condition |
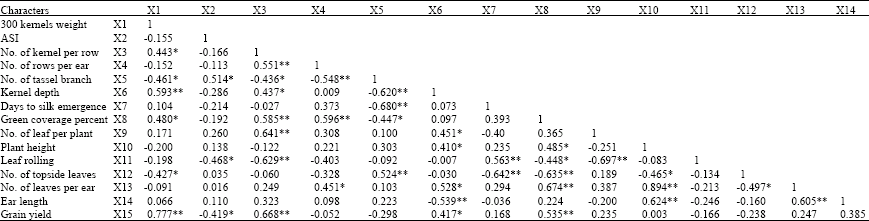 | |
| *Significant at p = 0.05; **significant at p = 0.01 | |
| Table 5: | Phenotypic correlation coefficients between grain yield and some yield components and growth characters of maize hybrids under drought stress at vegetative phase |
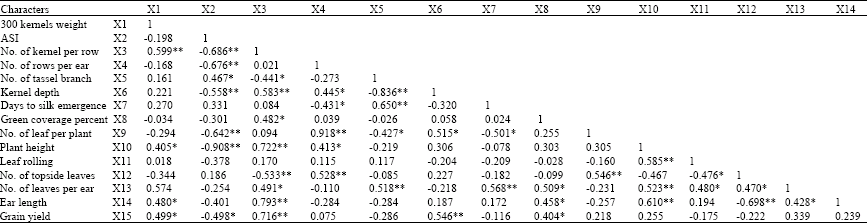 | |
| *Significant at p = 0.05; **significant at p = 0.01 | |
| Table 6: | Phenotypic correlation coefficients between grain yield and some yield components and growth characters of maize hybrids under drought stress at reproductive phase |
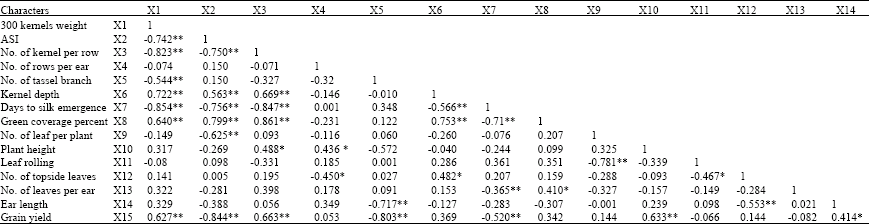 | |
| *Significant at p = 0.05; **Significant at p = 0.01 | |
| Table 7: | Phenotypic correlation coefficients between grain yield and some yield components and growth characters of maize hybrids under drought stress at grain filling phase |
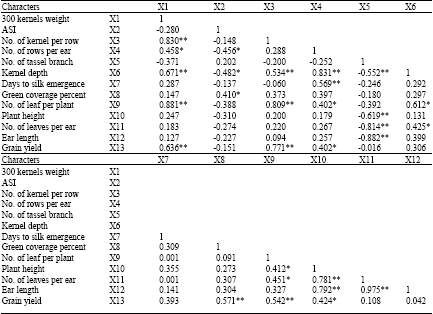 | |
| *Significant at p = 0.05. **Significant at p = 0.01 | |
Meanwhile, correlation of ASI with yield components and green cover percentage was negative and significant, but with number of branches of tassel and days to silk emergence was positive and significant (Table 6). As shown in Table 7, stress at kernel filling period brought positive and significant correlation between grain yield and 300 kernel weight (0.78), number of kernel per row (0.67), kernel depth (0.42) and green cover percentage (0.53). According to Claassen and Shaw (1970), stress at kernel filling period has not any effect on kernel number, but due to low leaf photosynthesis and less remobilization of assimilate toward seeds at this condition, weight of kernels will be decreased. Three key functions for water deficiency in maize are intensity and duration of drought stress, rate of stress development and phonological timing of stress (Galeta et al., 2005; Bolanos et al., 1993). Present analysis and several investigations show that, anthesis and silking phase (two weeks before emergence of silks), are most tender stages to drought and stress in these scenes will induce prolonged interval between anthesis to silking, decline of kernel number and thereby, reduced grain yield (Abdmishani et al., 2004; Cavalieri and Smith, 1985; Downey, 1977; Kabdal et al., 2003; Venugopal et al., 2003; CIMMYT, 2000).
The simple correlation coefficients between grain yield and some yield component and growth characters of under study single cross hybrids of maize were partitioned into direct and indirect effects using path analysis (Table 8-11). The estimated residual effects had indicated that the characters chosen for path analysis were appropriate. Moreover, the values of direct effects indicating the minimum inflation due to multi co-linearity (Mohammadi et al., 2003). The path analysis on grain yield of drought stress at reproductive phase (Table 8), revealed that 300 kernels weight (0.33) recorded high positive direct effect on grain yield and its indirect effects on grain yield via kernels/row (0.32) and ASI (0.85) was positive and via days to silk emergence was negative (-0.86).
| Table 8: | Phenotypic correlations with grain yield, direct and indirect effects of yield components at reproductive phase stress conditions |
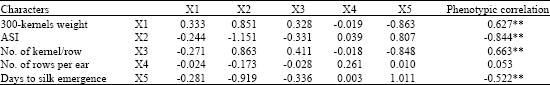 | |
| Bold values have direct effects. *Significant at p = 0.05. **Significant at p = 0.01, Residual effect: 0.107, R2 = 0.93 | |
| Table 9: | Phenotypic correlations with grain yield, direct and indirect effects of yield components at grain filling phase stress conditions |
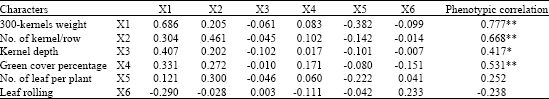 | |
| Bold values have direct effects. *Significant at p = 0.05. **Significant at p = 0.01, Residual effect: 0.107, R2 = 0.81 | |
| Table 10: | Phenotypic correlations with grain yield, direct and indirect effects of yield components at vegetative phase stress conditions |
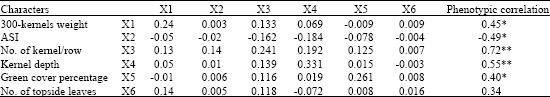 | |
| Bold values have direct effects. *Significant at p = 0.05. **Significant at p = 0.01, Residual effect: 0.004, R2 = 0.72 | |
| Table 11: | Phenotypic correlations with grain yield, direct and indirect effects of yield components at non-stress conditions |
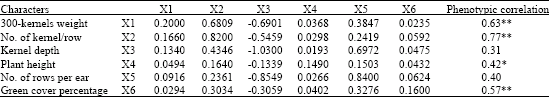 | |
| Bold values have direct effects. *Significant at p = 0.05. **Significant at p = 0.01, Residual effect: 0.102, R2 = 0.94 | |
The ASI had negative direct effect (-1.15) and negative indirect effects on grain yield via 300 kernel weight (-0.24) and kernels/row (-0.30). Direct effect of this trait proved to have impact on its correlation with grain yield at mentioned conditions. Therefore, simultaneously screening for ASI and days to silk emergence might be accomplished to select shorter ASI and drought tolerant single crosses of maize.
Considering drought stress at grain filling stage, 300 kernels weight (0.69), kernels/row (0.45), green cover percentage (0.17) and leaf rolling (0.23) had direct effect on grain yield (Table 10). On the other hand, indirect effects of green cover percentage via 300 kernel weight and kernels per row was more than its direct effect on grain yield. So, it could be concluded that, at grain filling period of drought prone condition, high green cover percentage plants with broad blade level are capable to produce more photosynthetic assimilates and remobilization them from vegetative organs to the grains and ears, which ultimately increase 300 kernels weight. The obtained outcomes are in consistent with those reported by Abdmishani et al. (2004).
At vegetative period drought stress, 300 kernels weight recorded medium level of direct effect and low level of indirect effect via kernels per row on grain yield. The path analysis revealed that, number of kernels per row (0.24) recorded positive direct effect and positive indirect effects via 300 kernels weight, kernel depth and green cover percentage on grain yield. Regarding to positive direct and indirect effects of this trait on grain yield and its components, it could be exploited for selection at drought resistance programs. The results presented by Singh and Singh (1993) emphasized that number of kernels per row; kernels weight and number pf ear per plant are the most important traits, respectively.
Number of kernels per row proved to have a high direct effect on grain yield (0.82) and its indirect effects via 300 kernels weight, number of kernels per row and kernel depth exhibited positive and noteworthy.
From the foregoing discussion, it may be concluded that, drought resistance dedicating traits are polygenic since drought tolerant single crosses had got highest yields at all moisture stress conditions (SC 686, SC 690 and SC 604, respectively). Moreover, the present study revealed that phenotypic correlations of important traits with each other and with grain yield at normal and stressed conditions mostly were concurrent. Though, values of correlations of ASI, number of kernels per row, days to silk emergence and ear length with grain yield at drought prone conditions varied considerable, which showing divers response of hybrids at different situations of moisture, since, sign and value of association of these traits affected sorely by media and stage of growth as well as intensity of drought stress.
REFERENCES
- Abdmishani, C., S. Vaezi, Y.B. Samadi and M. Ghannadha, 2004. Correlation and path analysis of grain yield and its components in maize. Maize Genet. Conf. Abstr., 46: 1-2.
Direct Link - Abrecht, D.G. and P.S. Carberry, 1993. The influence of water deficit prior to tassel initiation on maize growth, development and yield. Field Crops Res., 31: 55-69.
CrossRef - Agrama, H.A.S., 1996. Sequential path analysis of grain yield and its components in maize. Plant Breed., 115: 343-346.
CrossRefDirect Link - Bolanos, J. and G.O. Edmeades, 1996. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crops Res., 48: 65-80.
CrossRefDirect Link - Cavalieri, A.J. and O.S. Smith, 1985. Grain filling and field drying of a set of maize hybrids released from 1930 to 1982. Crop Sci., 25: 856-860.
Direct Link - Claassen, M.M. and R.H. Shaw, 1970. Water deficit effects on corn. I. Grain components. Agron. J., 62: 652-655.
CrossRefDirect Link - Denmead, O.T. and R.H. Shaw, 1960. The effects soil moisture at different stages of growth on the development and yield of corn. Agron. J., 52: 272-274.
Direct Link - Dewey, D.R. and K.H. Lu, 1959. A correlation and path-coefficient analysis of components of crested wheatgrass seed production. Agron. J., 51: 515-518.
CrossRefDirect Link - Downey, L.A., 1971. Effect of gypsum and drought stress on maize (Zea mays L.). I. growth, light absorption and yield. Agron. J., 63: 569-572.
CrossRefDirect Link - Galeta, N., H. Mohammed and H. Zelleke, 2005. Genetic variability, heritability and genetic advance in sorghum [Sorghum bicolor (L.) Moench] germplasm. Crop Res., 30: 439-445.
Direct Link - Kabdal, M.K., S.S. Verma, N. Ahmad and U.B.S. Panwar, 2003. Genetic variability and correlation studies of yield and its attributing characters in Maize (Zea mays L.). Indian Agri. Sci. Dig., 23: 137-139.
Direct Link - Moghaddam, M., B. Ehdaie and J.G. Waines, 1997. Genetic variation and interrelationships of agronomic characters in landraces of bread wheat from Southeastern Iran. Euphytica, 95: 361-369.
CrossRefDirect Link - Mohammadi, S.A., B.M. Prasanna and N.N. Singh, 2003. Sequential path for determining interrelationships among grain yield and related characters in Maize. Crop Sci., 43: 1690-1697.
Direct Link - Najeeb, S., A.G. Rather, G.A. Parray, F.A. Sheikh and S.M. Razvi, 2009. Studies on genetic variability, genotypic correlation and path coefficient analysis in maize under high altitude temperate ecology of Kashmir. Maize Genet. Cooperat. Newslett., 83: 1-8.
Direct Link - Richards, R.A., G.J. Rebetzke, A.G. Condon and A.F. van Herwaarden, 2002. Breeding opportunities for increasing the efficiency of water use and crop yield in temperate cereals. Crop Sci., 42: 111-121.
CrossRefDirect Link - Tollenaar, M., A. Ahmadzadeh and E.A. Lee, 2004. Physiological basis of heterosis for grain yield in maize. Crop Sci., 44: 2086-2094.
Direct Link - Westgate, M.E., 1994. Water status and development of the maize endosperm and embryo during drought. Crop Sci., 34: 76-83.
CrossRefDirect Link - Singh, G. and M. Singh, 1993. Correlation and path analysis in maize under mild-hills of Sikkim. Crop Improvement, 20: 222-225.
Direct Link
mohammad sadegh khosravi Reply
Its uniqe work. I suggest you to work on tolerate of lipids, phenols,sugers and ect......
good luck
K. Tehranian Reply
It's perfect. There are some profitable points in this article. I think it has been a part of another investigation to contrast different maturity groups. God bless you.