
S.O. Okoth
Trypanosomiasis Research Centre-Kenya Agricultural Research Institute, Box 362, Kikuyu
E.D. Kokwaro
School of Pure and Applied Sciences, Kenyatta University, Box 43844 Nairobi
J.M. Kiragu
Trypanosomiasis Research Centre-Kenya Agricultural Research Institute, Box 362, Kikuyu
G.A. Murila
Trypanosomiasis Research Centre-Kenya Agricultural Research Institute, Box 362, Kikuyu
Trends in Medical Research
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 75-85







ABSTRACT
The vectorial capacity of Glossina pallidipes from two allopatric populations of Kenya was evaluated. Tenerals flies from the two subpopulations were infected with T.b. rhodesiense in vivo, maintained in vitro and allowed to transmit the infection to susceptible mice 25 days post infection. Infection status of the flies was analyzed parasitologically and biochemically through microscopy and Polymerase Chain Reaction (PCR). Results showed that flies from Nguruman had lower survival rates and were less susceptible to trypanosome infection than those from Busia, although the differences were insignificant. There was no significant difference in infection rates of the proboscis, midgut and salivary gland between the subpopulations. Infection load of the dissected fly parts varied significantly among the Nguruman (F = 20.043, df = 2, p<0.001) and also among the Busia (F = 26.167; df = 2; p<0.001) subpopulations. Mean infection load and transmission success to mice between the two subpopulations were however not significantly different. An insignificant inverse correlation between means of PCV and parasitaemia was observed. However, trypanosome infection did not significantly vary PCV between infected and control mice. Implications of these results are discussed with respect to epidemiology of rhodesian sleeping sickness.
PDF Abstract XML References
How to cite this article
S.O. Okoth, E.D. Kokwaro, J.M. Kiragu and G.A. Murila, 2006. Susceptibility and Transmission Capacity of Subpopulations of Glossina pallidipes to Human Infective Trypanosoma brucei rhodesiense. Trends in Medical Research, 1: 75-85.
URL: https://scialert.net/abstract/?doi=tmr.2006.75.85
URL: https://scialert.net/abstract/?doi=tmr.2006.75.85
Introduction
Sleeping sickness is endemic in 36 countries of sub Saharan Africa where approximately 60 million people are at risk (Aksoy, 2003; WHO, 1994). Glossina pallidipes is a known vector for rhodesian sleeping sickness. Vector-related factors that influence the epidemiology of sleeping sickness in Kenya are not well understood. Intraspecific variations in vector competence have been reported among subpopulations of Glossina sp. and are believed to contribute to the focal distribution of rhodesian sleeping sickness (Welburn and Maudlin, 1999). According to Aksoy (2003) and Aksoy et al. (1997), both intrinsic genetic factors and biological factors such as presence of endosymbionts influence vector susceptibility to infection by parasites and the subsequent transmission. The occurrence of endosymbiont and gut molecules that modulates vector competence varies across tsetse subpopulations (Aksoy, 2003). The intracellular endosymbiont are thought to act in diverse ways (Scott et al., 1993; Aksoy et al., 1997), for example, Wigglesworthia and Wolbachia have been reported to play roles in tsetse reproduction and survival (Aksoy et al., 1997) and survival of tsetse population is a key determinant of its vectorial capacity. In addition, midgut lectins (agglutinins), trypanoagglutinins, trypanolysins (Ibrahim et al., 1984) and digestive proteases (Stiles et al., 1991; Welburn and Maudlin, 1999) mediate establishment and maturation of trypanosomes in tsetse flies. Earlier attempts to compare the vectorial capacity of allopatric populations in Kenya used G. pallidipes from non-sleeping sickness foci and non-human infective Trypanosomes (Moloo, 1993) and did not therefore reveal much with respect to epidemiology of rhodesian sleeping sickness. In the present study, tsetse subpopulations from an endemic and non-endemic areas with a human infective T. b. rhodesiense were used. The role of vector-associated factors in the transmission of T. b. rhodesiense and their possible contribution to the focal nature of rhodesiense sleeping sickness is discussed.
Materials and Methods
Study Areas
This study was carried out in Busia and Nguruman. Busia study area lies between latitude 0°136' South and 0°North and longitudes 33°54' east and 340 25' 24'’ East (Fig. 1). The area is infested with G. f. fuscipes along the riparian forest patches and G. pallidipes which has patchy distribution, associated with woody hillside vegetation (Ford, 1971). Nguruman lies at latitude 1°55' S and longitude 35°25' E on the floor of the rift valley in southern Kenya (Fig. 2). The area is infested by G. pallidipes and G. longipennis within the woodlands and G. swynnertoni on the adjoining escarpments (Brightwell et al., 1997).
 | |
| Fig. 1: | A map of Western Kenya showing the study area |
 | |
| Fig. 2: | Map of Nguruman showing the study area |
Collection of Field Flies and Maintenance in the Insectary
Unbaited biconical traps (Challier and Laveissiere, 1973) were used for collecting flies from the field. Caught flies were transported to the rearing facilities, sorted and placed in rearing cages. The flies were maintained by in vivo feeding from rabbit ears. Room conditions were maintained at 70±2% relative humidity and 23±2°C.
Pupae Production and Maintenance
Each day, pupae were collected from the rearing trays and placed in a 20 mL plastic tube lined with cotton wool. Every succeeding three-day collection were grouped together to maintain uniform age groups and to synchronize emergences. The pupae were transported by road to the experimental insectary at KARI-TRC, Nairobi for incubated.
In vivo Infection, in vitro Maintenance and Probing of Flies
Emerged teneral flies were allowed to obtain their first blood meal from a mouse infected with a T. b. rhodesiense clone, KETRI 3537. At peak parasitaemia, three or four tsetse flies in a cage were held close to the mouse belly to feed. Engorged flies were returned to the insectary where they were maintained in vitro on bovine blood for 25 days. Day 23 post infective feed, flies were starved for two days and probed on warmed slide.
In vivo Transmission of Trypanosomes to Mice
In vivo transmission was used to transfer infection from the tsetse fly to mice through natural feeding process. Tsetse flies that had been fed on infectious mice as tenerals were allowed to feed on clean Swiss white mice, one fly per mouse. Each mouse was monitored for parasitaemia and Packed Cell Volume (PCV) every day for 20 days. Trypanosomes were detected in the whole blood by dark-ground buffy coat phase contrast technique (Murray et al., 1977). PCV from heparinised whole blood samples was measured after haematocrit centrifugation using the method described by Schalm et al. (1975).
Determination of Infection Rate and Infection Load in Tsetse Flies
Flies were starved for two days then dissected at the mouthparts, midgut and salivary gland. The fly parts found infected with trypanosomes were cut-off and transferred into a 0.2 mL conical ampoule. Zero point one mL of 20% glycerine Phosphate-Buffer Saline Glucose (PBS), was used to make a homogenous suspension from which a wet smear was made and mean trypanosome load determined underx400 magnification. Sample of each fly found parasitologically negative in all its three dissected parts were pooled into an ampoule and stored at -4°C for PCR analysis.
Preparation of Template for PCR
Ampoules were thawed and the contents expelled into single 1.5 mL micro centrifuge tubes containing 0.5 mL lysis buffer (1%v/v Triton-100 in 10 mM Tris-HCL, pH 7.5) in which they were vortexed briefly. The material was pelleted by centrifugation at 13,000 g for 10 min and the supernatant discarded. This step was repeated until the material was free of haemoglobin. The final pellet was resuspended in 100 μL of PCR buffer (10 mM Tris-HCl, pH 8.3, 50 mM KCl, 1.5 mM MgCl2) with 60 μg mL-1 proteinase K. The mixture was incubated at 55°C for 1 h and at 95°C for 10 min to denature the protainase K. Two microliters of this extract was used as a template for PCR amplification.
DNA Amplification, Electrophoresis and Filming
Amplification of trypanosome DNA was performed as a 20 μL reaction mix in a micro-centrifuge tube. The reaction mixture consisting of 10 mM Tris-HCl, pH 8.3, 1.5 mM MgCl2, 50 mM KCl, 150 μM of each deoxynucleotide triphosphate, 0.8 μM of each pair of trypanozoon specific primers (TBR1 and TBR 2) (PE Applied Biosystems, USA), 2 μL of the DNA extract and 0.5 units of Taq DNA polymerase (Fisher). The reaction mixture was placed in a thermo- cycler and incubated at 94°C for 1 min, followed by 30 cycles of denaturation at 92°C for 30 sec, annealing at 60°C for 45 sec, extension at 72°C for 45 sec and a final extension at 72°C for 4 min. Fifteen microliters of each PCR product was analyzed by gel electrophoresis through 1.5% agarose gels stained with ethidium bromide and photographed using a Polaroid camera.
RESULTS
Tsetse Rearing and Pupae Production in Field Insectaries
Pupae production per day averaged 22 in Nguruman and 35 in Busia subpopulation colonies. A total of 879 and 1,256 pupae were collected from Nguruman and Busia respectively. Out of the 2135 pupae collected, 564 (26.42%) emerged, 303 (24.12%) from Busia and 261 (29.69%) from Nguruman subpopulation groups.
| Table 1: | Mean daily survival and mean percent survival of tsetse flies to day 25 |
| Table 2: | Mean (%) trypanosome infection rate of tsetse flies from Busia and Nguruman |
 | |
| Figures followed by the same letters within a column are not significantly different at 0.05% LSD | |
| Table 3: | Mean trypanosome infection load in Glossina pallidipes 25 days post infection |
 | |
| Figures followed by the same letters within a column are not significantly different at 0.05% by LSD | |
| Table 4: | Transmission success of trypanosomes to mice by tsetse flies detected with infection by microscopy |
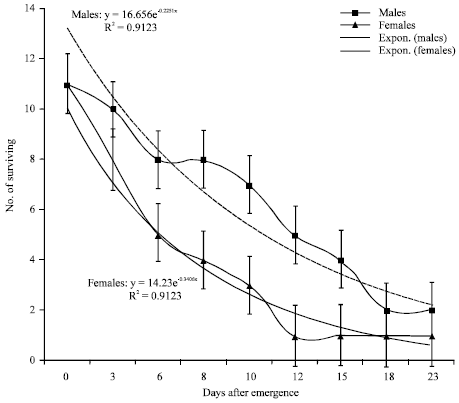 | |
| Fig. 3: | Survival curves and exponential trend line for male and female tsetse flies from Nguruman |
Survivals of Experimental Flies
Analysis of variance showed that the daily survival rates of flies from Nguruman and Busia subpopulations did not differ significantly (t = 0.420, df = 7, p = 0.687). Paired sample t-test for mean percent survival upto day 25 showed no significant difference (t = 0.989, df = 8, p = 0.395) between the two-subpopulation groups (Table 1).
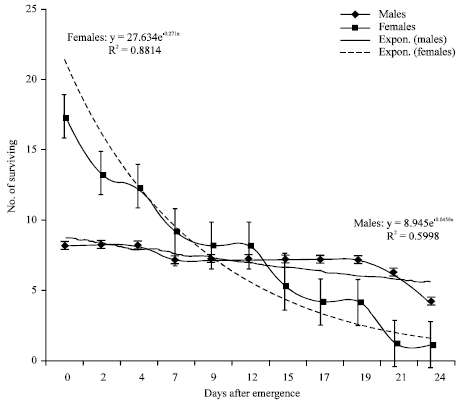 | |
| Fig. 4: | Survival curves and exponential trend lines for male and female tsetse flies from Busia |
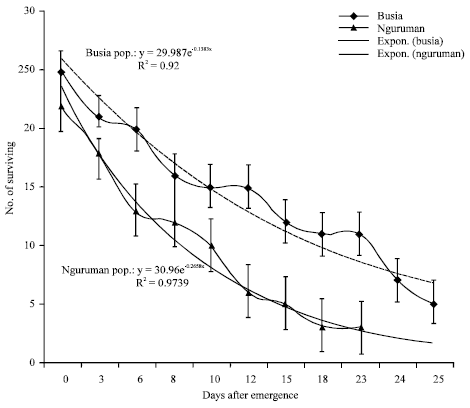 | |
| Fig. 5: | Survival curves and exponential trend line for tsetse flies from Busia and Nguruman subpopulations |
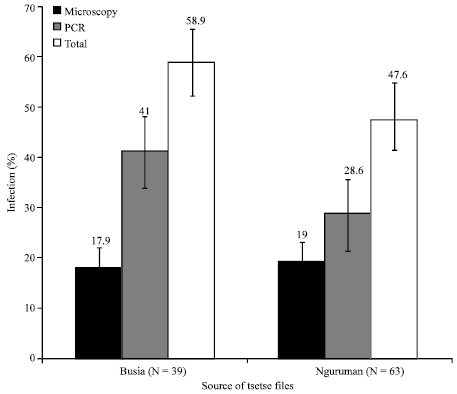 | |
| Fig. 6: | Percent infection of flies from Nguruman and Busia subpopulation flies as detected by microscopy and PCR |
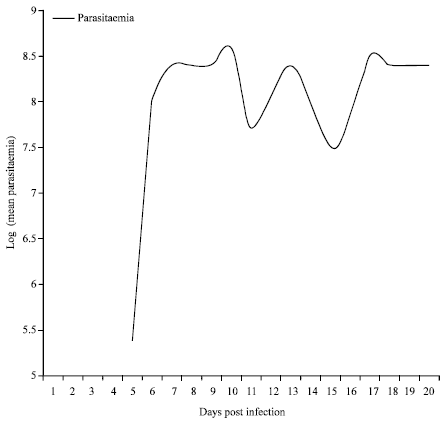 | |
| Fig. 7: | Mean parasitaemia in mice infected with T. b. rhodesiense using flies from Busia sub population |
Both sexes had similar survival in the Nguruman subpopulation (Fig. 3). Males had higher survival rates than females in the Busia subpopulation (Fig. 4). On average, Nguruman subpopulation recorded lower survival than the Busia subpopulation (Fig. 5).
Susceptibility of Experimental Flies to Trypanosome Infection
Infection rates between the Nguruman and Busia subpopulation flies were not significantly different (χ2 = 0.387, df = 1, p = 0.534). However, Busia subpopulation flies had higher infection rate (58.9%; N = 63) than Nguruman subpopulation flies (47.6%; N = 39). Figure 6 shows that higher infection was detected by microscopy among the Busia subpopulation (19%) than the Nguruman subpopulation (17.9%) flies. Contrastingly, PCR detected higher proportion of trypanosome infections among the Nguruman (41.0%) than the Busia (28.6%) subpopulation flies. Table 2 shows that all the infected flies had midgut infection, 19% for Busia and 17.9% for Nguruman subpopulation. Table 3 shows that infection load was highest in the midgut (8.2) followed by the salivary gland (1.95) and proboscis (1.18) in the Busia subpopulation flies.
Infection rates in the proboscis were 2.6% (N = 39) and 3.2% (N = 63) in the Nguruman and Busia subpopulations flies, respectively. Salivary gland infection was 2.6% (N = 39) in the Nguruman subpopulation and 4.8% (N = 63) in the Busia subpopulation flies. Chi-square test showed no significant difference among infected parts (χ2 = 0.024, df = 2, p = 0.988).
Analysis of variance of the Busia flies showed significant difference in infection load among the three parts dissected (F = 26.167; df = 2; p<0.001). Separation of means by Least Significance Difference (LSD) showed that infection load in the midgut was significantly higher than in the proboscis and salivary gland. Analysis of variance of the Nguruman subpopulation showed that there were significant differences in infection load among the three parts dissected ((F = 20.043, df = 2, p<0.001). Separation of means by LSD showed that midgut had significantly higher infection load than proboscis and salivary gland. Comparison of mean infection loads between the Busia and Nguruman subpopulation groups using chi square test showed no significant difference (χ2 = 0.646, df = 2, p = 0.439).
In vivo Transmission of Trypanosomes to Mice by Infected Tsetse Flies
Forty one point six percent (N = 12) of flies from the Busia subpopulation successfully transmitted trypanosomes to mice, while none (0%, N = 7) of the flies from the Nguruman subpopulation group transmitted the infection to mice (Table 4). Chi square test of association between subpopulation group and transmission success indicated insignificant difference (χ2 = 2.601, df = 1, p = 0.107).
Parasitaemia and Packed Cell Volume in T. b. Rhodesiense Infected Mice
Five flies successfully transmitted trypanosomes to mice in the Busia subpopulation. The mean prepatent period in mice was 5 days. The first parasitaemia wave peaked on day 10 followed by the second and third on days 13 and 17, respectively (Fig. 7). Mice survival was 100% to day 15. Mortality 60% was recorded between on day 16, while 40% survived to day 20 (Fig. 8). There was however no successful transmission of trypanosomes to mice to Nguruman subpopulation flies. Mean Packed Cell Volume (PCV) of infected mice was 36.96±1.1 SE while that of control mice was 38.95±1.0 SE. Two-sample t- test for mean PCV indicated no significant difference (t = 1.35, df = 31, p = 0.187). Linear trend lines indicated stable PCV level for the control mice and a gradual decline in PCV for the infected mice with time (Fig. 9). Mean PCV and mean antilog parasitaemia showed a weak and insignificant inverse correlation (r = -0.390, p = 0.135).
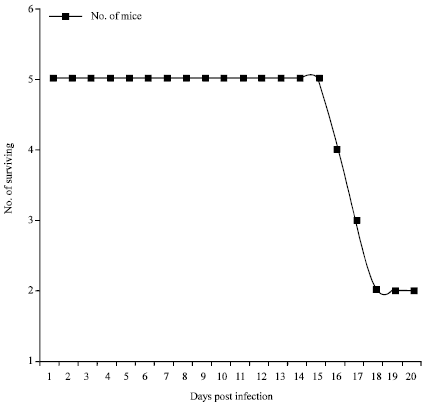 | |
| Fig. 8: | Survival of mice infected with T. b. rhodesiense using flies from Busia |
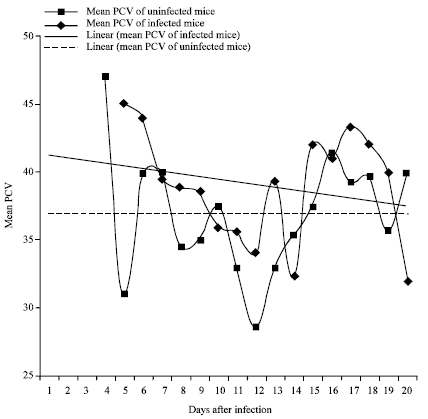 | |
| Fig. 9: | Changes in mean packed cell volume and linear trend lines in mice infected naturally with T. b. rhodesiense by flies from Busia |
DISCUSSION
The present study demonstrated remarkable differences in vector density between Western Kenya (Busia) and Southeastern Kenya (Nguruman). Vector density plays a central role in transmission of vector borne parasites to susceptible hosts, as the average host-biting rate is dependent upon the per capita vector load. Although Weir and Davidson (1965) reported that other factors such as host abundance, host preference and human-vector contact also influence transmission, higher human biting rate would be expected at Nguruman. Human biting rate is a measure of risk of transmission of humans and therefore human population at Nguruman stand at higher risk of contracting the infection should the human infective T. b. rhodesiense circulate in the vector population. The present study also showed that G. pallidipes from Western Kenya had higher survival and longevity than those from Southeastern Kenya. The competence of a vector population depends on the average longevity of individuals in the population (Macdonald, 1957). For T. b. rhodesiense, which requires considerably longer time to undergo its reproductive cycle in the invertebrate host, a vector population with high survival and longevity would be more suitable and this therefore, indicate that Busia subpopulation would be a better vector.
The present study showed that G. pallidipes subpopulation from Western Kenya was more susceptibility to trypanosomes infection than those from Southeastern Kenya. Susceptibility to trypanosome infection is regulated by several factors such as endosymbionts and genetic predisposition (Aksoy, 2003). Influence of heritability on vectorial capacity in insects has been reported (Gooding, 1988). Wakelin (1978) insinuated that susceptibility to infection depend on the genotype of the insect and often on the inheritance of a single gene. Comparative genetic studies based on microsatellite loci between the two subpopulations showed significant variability (Nei, 1987). Part of the variation in susceptibility to trypanosome infection between the two subpopulations may therefore be attributed to genetic variability, although this needs to be quantified. In an earlier study (Moloo, 1993) found that G. pallidipes from Nguruman were more susceptible to T. congolense infection than those from Shimba hills. This finding is consistent with the genetic data (Nei, 1987) where Nguruman and Shimba hills subpopulations were found to be closer and Shimba hills was genetically further from Busia subpopulation than Nguruman subpopulation. Data on tsetse endosymbionts in the two areas is however lacking and it would be interesting to study the occurrence and possible contribution of endosymbionts in the observed variations in susceptibility. Another striking difference between the two tsetse subpopulations was in their ability to transmit infection to susceptible host. The reason Nguruman subpopulation flies which showed mature transmissible metacyclics could not transmit the parasites to mice is not clear. However, it could have been due to the low metacyclic load detected in the salivary gland or other factors peculiar to the subpopulation.
These findings reveal the underlying differences between the two subpopulations and, by extension, other G. pallidipes subpopulations with respect to tsetse longevity and refractoriness to trypanosome infection and transmission capacities, which are important factors in vectorial capacity and overall epidemiology of sleeping sickness. In general, the present study revealed that G. pallidipes subpopulation from western Kenya was more competent in acquiring, establishing and transmitting T. b. rhodesiense than that from the southeastern Kenya, an observation which partially explains the localised distribution of rhodesian sleeping sickness to specific foci in Kenya.
ACKNOWLEDGMENTS
I am grateful to WHO/TDR, Swiss Tropical Institute (STI) and KARI-TRC for funding this work. Thank also to Godfrey Emase, Joseph Etyang, Charles Nambiro Sam Guya, Kariuki Ndungu and Purity Gitonga for their invaluable effort in making the bench work a success.
REFERENCES
- Aksoy, S., 2003. Control of tsetse flies and trypanosomes using molecular genetics. Vet. Parasitol., 115: 125-145.
PubMedDirect Link - Aksoy, S., I. Maudlin, C. Del, A. Robinson and S. ONeill, 2001. Prospects for control of African Trypanosomiasis by tsetse vector manipulation. Trends Parasitol., 17: 29-35.
CrossRefDirect Link - Aksoy, S., X. Cheng and V. Hypsa, 1997. Phylogeny and potential transmission routes of midgut-associated endosymbionts of tsetse (Diptera: Glossinidae). Insect Mol. Biol., 6: 183-190.
PubMedDirect Link - Brightwell, R., R.D. Dransfield, P. Stevenson and B. Williams, 1997. Changes over twelve years in populations of Glossina pallidipes and Glossina longipennis (Diptera: Glossinidae) subject to varying trapping pressure at Nguruman, south- west Kenya. Bull. Entomol. Res., 87: 349-370.
Direct Link - Ibrahim, E.A., G.A. Ingram and D.H. Molyneux, 1984. Haemagglutinins and parasite agglutinins in haemolymph and gut of Glossina. Tropenmedezin unde Parasitol., 35: 151-156.
PubMedDirect Link - Moloo, S.K., 1993. A comparison of the susceptibility of two allopatric populations of Glossina pallidipes for stocks of T. congolense. Med. Vet. Entomol., 7: 369-372.
PubMedDirect Link - Murray, M., P.K. Murray and W.I.M. McIntyre, 1977. An improved parasitological technique for the diagnosis of african trypanosomiasis. Trans. R. Soc. Trop. Med. Hyg., 71: 325-326.
PubMedDirect Link - Welburn, S.C., I. Maudlin and D.H. Molyneux, 1994. Midgut lectin activity and sugar specificity in tenerals and fed tsetse. Med. Vet. Entomol., 8: 81-87.
PubMedDirect Link - Welburn, C.S. and I. Maudlin, 1999. Tsetse-trypanosome interactions: Rites of passage. Parasitol. Today, 15: 399-403.
PubMed