
Okonkwo Francis Obiora
Environmental and Occupational Toxicology Unit, Department of Biochemistry, Faculty of Natural and Applied Sciences, Plateau State University, Bokkos, Nigeria
LiveDNA: 234.31834
Shockden Chinnan Isaac
Environmental and Occupational Toxicology Unit, Department of Biochemistry, Faculty of Natural and Applied Sciences, Plateau State University, Bokkos, Nigeria
Botson Mafeng Christopher
Environmental and Occupational Toxicology Unit, Department of Biochemistry, Faculty of Natural and Applied Sciences, Plateau State University, Bokkos, Nigeria
Ogbu Ambrose Ejiofor
Department of Biochemistry, University of Nigeria, Nsukka, Nigeria
Singapore Journal of Scientific Research
Year: 2021 | Volume: 11 | Issue: 1 | Page No.: 46-58







ABSTRACT
Background and Objective: Environmental pollution resulting from the discharge of azo dyes into water bodies is an onerous challenge presenting human and ecological risks. In the present study, the use of biocatalysts, enzymes, in the degradation of six azo dyes (Azo Trypan Blue, Azo Blue 5, Azo Yellow 6, Citrus Red, Azo purple and Azo pink) is considered as an eco-friendly approach using commercially available low-cost materials, fresh and ripening cucumber. Materials and Methods: Standard methods and protocols were utilized in the purification, characterization and determination of optimal conditions for the resulting peroxidase enzymes. The decolorization of azo dyes was determined using the difference between initial and final absorbance of the dye solutions over 20 hrs under the determined optimal conditions. Results: From the results, the optimal temperature was observed to be 40 and 45°C for fresh and ripening cucumber peroxidase respectively and a pH of 5.5 for both the fresh and ripening cucumber peroxidase. Azo trypan blue had the highest % decolorization of 89.4 for ripening cucumber followed by Azo Blue 5 and Azo Purple with 82.3 and 74.5% decolorization respectively. For fresh cucumber peroxidase, Azo Blue 5 had the highest % decolorization of 76.3 followed by Azo purple and Azo trypan blue with 71.1 and 63.8%, respectively after a contact time of 20 hours. Conclusion: The outcome of the present work can find application in the enzymatic treatment of wastewater containing azo dyes especially with ripening cucumber considered as waste and not fit for consumption.
PDF Abstract XML References
Received: December 21, 2019;
Accepted: July 15, 2020;
Published: December 15, 2020
How to cite this article
Okonkwo Francis Obiora, Shockden Chinnan Isaac, Botson Mafeng Christopher and Ogbu Ambrose Ejiofor, 2021. Comparative Decolorization of Synthetic Azo Dyes using Peroxidase Purified from Fresh and Ripening Cucumber. Singapore Journal of Scientific Research, 11: 46-58.
URL: https://scialert.net/abstract/?doi=sjsres.2021.46.58
URL: https://scialert.net/abstract/?doi=sjsres.2021.46.58
INTRODUCTION
Liquid wastes (effluents) resulting from dyeing and finishing of textile materials are considered the most noxious of all industrial effluents. This is putting into consideration the volume of production and the chemical composition of the ensuing wastes1. With the world population on the increase leading to an increase in demand and supply of textile goods and the resultant increase in the production and utilization of synthetic dyes to ensure quality, have equally contributed to the substantial and significant rise in dye wastewater pollution of water sources in current times2,3. Azo dyes make up a significant percentage of dyestuffs employed in the textile industries, as well as equally playing substantial roles in the printing, food, paper making and cosmetic industries.
Azo dyes are recalcitrant and xenobiotic compounds which are characterized by the presence of one or more azo groups (-N = N-), which makes them extremely difficult to eliminate by biodegradative processes4. Studies have shown that the ingestion of water contaminated with azo dyes and their by-products, may be carcinogenic and/or mutagenic5. Wastewater treatment systems and water treatment plants as seen in developing countries are largely ineffective both in the removal of color and the amelioration of the mutagenic properties of the dyestuffs hence endangering the environment as well as human lives. This ineffectiveness and difficulty are as a result of these dyes being produced to withstand wear and tear and ensure optimum quality of the textile and other products. Hence these azo dyes are stable and are designed to withstand biodegradation all in a bid to meet market quality demands and durability of colors in the textile and sundry products. Consequently, these dyes tend to persist in the environment for a very long time6.
A conventional biological wastewater treatment process is not very efficient in treating dye wastewater due to the low biodegradability of dyes. It is usually treated by physical or chemical treatment processes, but these procedures have inherent drawbacks as they generate a significant amount of the sludge or cause secondary pollution due to the formation of hazardous by-products7. Therefore, treatments that are more efficient and economical than those currently available are required as a matter of urgency and importance. The use of biological systems such as enzymatic approach for the treatment of these wastewaters has become an attractive alternative because enzymes can act on specific recalcitrant pollutants and the catalytic action of enzymes are extremely high as compared to chemical catalysts due to higher reaction rates and milder reaction conditions. It is a low cost, environmentally friendly technology for the extraction, degradation, or fixation of the contaminants8.
Peroxidase (EC.1.11.1.7.) is a ubiquitous enzyme that belongs to the oxidoreductase class of enzyme and generally catalyzes the reaction between H2O2 as electron acceptor and many kinds of substrates by means of O2 liberation from H2O2 9. They are found in bacteria, fungi, algae, plants and animals10. The Scientific Committee of the European Union has also defined peroxidases as the group of proteins of greatest biotechnological interest in the 21st century11. The definition above is based on the potential contribution of these enzymes to the conservation of the environment as they are potential substitutes of other polluting industrial catalysts. It represents a group of enzymes widely distributed in the plant kingdom and has been extracted from several plants including horseradish (Armoracia rusticana), peach (Prunus persica), yam (Alocasia macrorrhiza), manioc (Manihot utilissima), artichoke (Cynara scolymus L.), sweet potato (Ipomoea batatas ex L., Lam.), turnip (Brassica campestris apifera), zucchini (Cucurbita pepo) and others12. It has also been found in the spleen, lungs, mammary and thyroid glands, bone marrow and intestine13. Zia et al.14 purified this enzyme from Streptococcus faecalis.
In biotechnology and associated research areas (enzymology, biochemistry, medicine, genetics, physiology, histo- and cytochemistry), peroxidases have conquered a prominent position15 and it remains the major enzyme used to evaluate the heat processing of vegetables, having wide range of industrial applications in the area of agriculture, analytical, environmental, medical sectors, etc. Thus, peroxidases have been used in bioremediation of contaminating environmental pollutants such as phenols, delignification in paper and pulp industry, diagnosis kit development, immunoassay, organic and polymer synthesis as well as in biosensor technology16. They are also used for developing convenient and quick methods for the determination and quantification of hydrogen peroxide in both the biological and industrial samples17,18. Additional applications of peroxidases include determination of extent of lipid peroxidation in meat food products, in polymerization and precipitation of aqueous phenols as well as in decolourization of industrial effluents19. The limitation on the use of plant peroxidases is the low yield and high production cost compared with bacterial or fungal enzymes. However, the production cost of these enzymes can be reduced by optimizing the extraction conditions and by using plant material with high enzymatic activity that can be directly applied with the same efficiency as the purified enzyme20.
Due to the widespread use of peroxidases, mainly as an environmental biocatalyst, there is a growing interest in new sources of this enzyme. The development of environmentally sustainable processes is also an onerous challenge for the current bio-economy. In this direction, the use of biocatalysts, enzymes, in various processes is considered as eco-friendly approach. Therefore, the identification of newer sources for such novel enzymes with desired properties to find effective treatments for effluents and drinking water is important. It is suggested that the fresh cucumber and the wastes obtained from ripening cucumber could be considered as an economic peroxidase source to replace the commercial Horseradish peroxidase and other peroxidase sources.
The most widely used and commercially available peroxidase is Horseradish peroxidase (HRP), from the root of horseradish (Amoracia rusticana L.). Although accounting for 90% of the world's production of peroxidases21, HRP has drawbacks affecting its stability and inactivation under certain conditions and is not commonly available. In search of increasingly stable enzymes to overcome these problems, peroxidases from fresh and ripening cucumbers could be a cheaper substitute. There is a dearth of literature on the use of fresh and ripening cucumber as a veritable source of peroxidase enzyme. Therefore, the main purpose of this study was to extract, purify and characterize the peroxidase from fresh and ripening cucumber (Cucumis sativus) for decolorization of synthetic dyes and determine which sample has the highest activity and percentage decolorization. Cucumbers are generally cheap and suffer lots of post-harvest losses and the ripening ones which are not fit for consumption could be channeled to the production of enzymes, reducing wastes and increasing income status of the rural farmer. This study is strictly limited to the extraction, purification and characterization of peroxidase enzyme from fresh and ripening cucumber and checking its potential in the decolorization of some azo dyes.
MATERIALS AND METHODS
Materials/Reagents: Between July and September 2018, fresh cucumber samples were purchased from local markets in Maikatako, Barkin-Ladi Local Government Area of Plateau State, Central Nigeria while ripening and rotting cucumber samples, not fit for consumption, were collected from the waste dumps behind the same markets. The synthetic dyes (Azo Trypan Blue, Azo Blue 5, Azo Yellow 6, Citrus Red, Azo purple and Azo pink) used for the study were purchased from the Clothing and Textile Unit, Fine and Applied Arts Department, Faculty of Arts, University of Jos, Plateau State. Ammonium sulphate, Bovine Serum Albumin (BSA), Folin Ciocalteu, O-dianisidine, Sephadex G-75 and hydrogen peroxide were obtained from Sigma-Aldrich, Taufkirchen, Germany. All other chemicals used in this study were of analytical grade and obtained from several reputable commercial sources in Europe and North America.
Extraction of crude enzymes: Peroxidase was extracted from fresh and ripening cucumber (Cucumis sativum) by the method of Genga et al.22 with minor modifications as seen below. The fresh and ripening cucumber samples were thoroughly washed in distilled water and chopped separately using a blender. Two hundred and fifty grams of each cucumber sample was mixed with 500 mL of phosphate buffer (0.05M) pH 7.0. The mixture was left for 24 hrs with constant stirring using a magnetic stirrer. The homogenate was filtered with double-layered cheesecloth. The filtrates were centrifuged at 4000 rmp for 30 min to remove cell debris. The supernatant solution (the crude enzyme extract) was collected, volume recorded and stored at a temperature of 4°C in the refrigerator and used as the enzymatic source of the peroxidases.
Determination of protein concentration and enzymes activity: The protein content of crude enzymes was determined by the method of Lowry et al.23, using Bovine Serum Albumin (BSA) as the standard protein. Peroxidase activity was determined using the method of Eze et al.24, with little modification. The assay mixture contained 2.7 mL of sodium phosphate buffer pH 7.0 (0.05M), 0.1 mL of 0.8% H2O2 (0.025 M), 0.1 mL of 1% O-dianisidine and 0.1 mL of the enzyme extract, giving a total volume of 3 mL. The mixture components were added in the order stated above. The mixture without the enzyme was first put in a cuvette and blank before adding the enzyme. Immediately the enzyme was added, the change in absorbance due to oxidation of O-dianisidine in the presence of H2O2 was monitored using UV spectrophotometer at 460 nm. The readings were taken at 30 sec interval for 5 min.
In this case, one unit of enzyme activity was defined as the amount of enzyme that gave an absorbance change of 0.1/minute at 30°C. The equations below were used to calculate the purification steps of the experiments using the method of Khurshid et al.25:
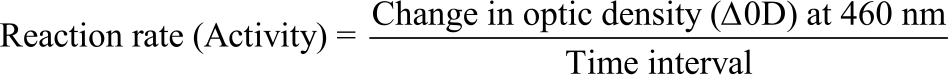 |
Since the volume of the enzyme used was 0.1 mL and the time interval was 60 sec (1 min):
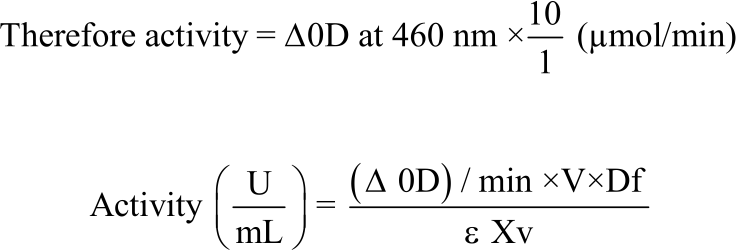 |
where, V is total volume of reaction mixture = 3 mL, v is enzyme volume = 0.1 mL, Df is the dilution factor and ∈ is micromoler extinction coefficient of O-dianisidine = 11.3 mM–1 cm–1 at 460nm26.
Therefore, Activity = Δ0D×2.655×Dilution factor:
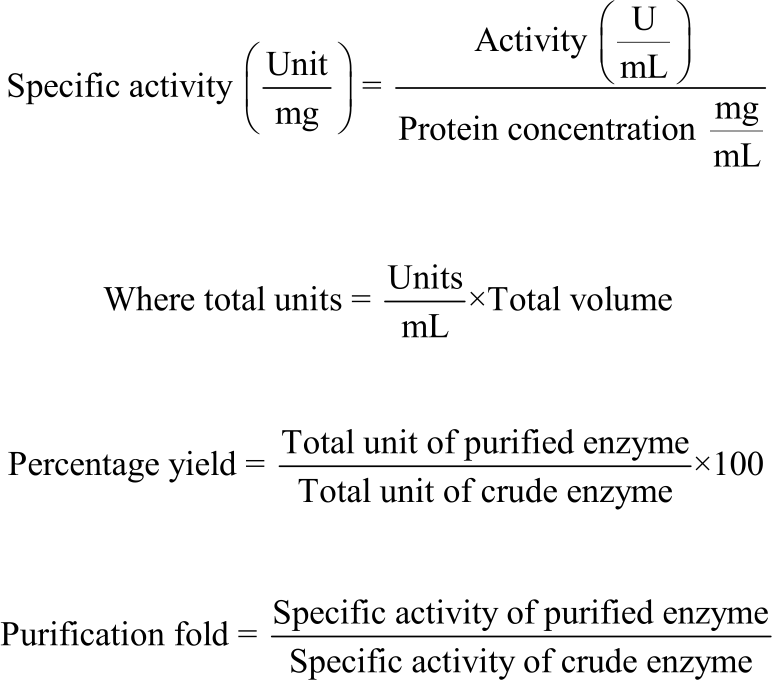 |
Partial purification and dialysis of peroxidases enzyme: In the first stage, solid ammonium sulphate of different saturation ranging from 10-90% at an interval of 10% in each test tube containing 10mL of the crude enzymes was used to determine the % saturation with the highest activity for both the fresh and ripening cucumbers as shown in Fig. 1. From the assayed precipitates, highest peroxidases activity and protein concentrations were achieved at 80 and 50% ammonium sulphate saturation for the fresh and ripening cucumber respectively and were used to precipitate the crude enzymes. Solid ammonium sulphate salt was added slowly to the crude enzymes in a beaker placed on a magnetic stirrer plate until the salt was completely dissolved to attain optimum % saturation. The mixture was maintained at 4°C for 30 hrs. The solution was then centrifuged at 4000 rmp for 30 min after which the pellet was collected and dissolved in a small amount of buffer in which the enzymes were originally extracted. The dissolved pellets were taken as the partially purified peroxidases. Protein concentrations and peroxidases activity of the precipitates were then determined following already described methods23,24.
The solutions were kept separately in a 10 cm pretreated dialysis bag after sealing securely and dialyzed against sodium phosphate buffer pH 7.0 (0.05 M) suspended in a beaker.
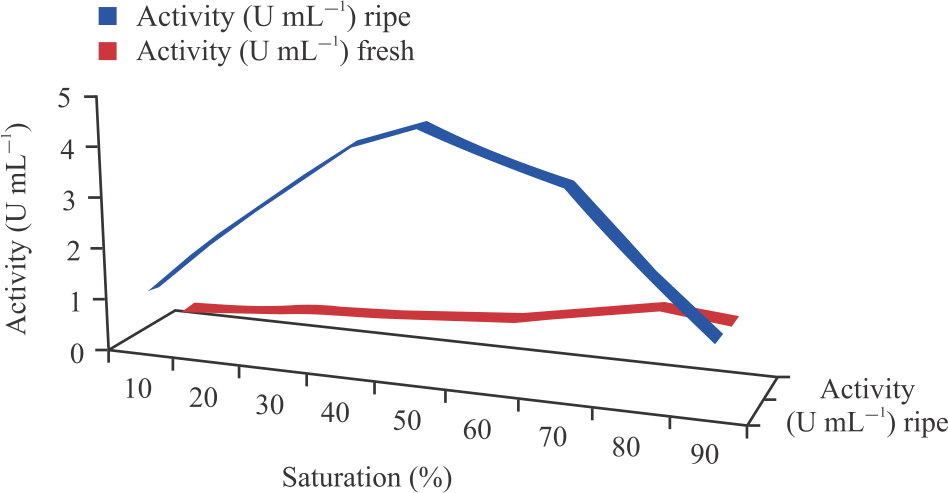 |
| Fig. 1: | Ammonium sulphate precipitation profile for peroxidases extracted from fresh and ripening cucumber |
This is placed on a magnetic stirrer for 18 hrs with continuous stirring. The buffer was changed every 6 hrs to remove ions and other low molecular weight substances that have diffused from the dialysis bag into the buffer. This was to prevent the ions and low molecular weight substances from diffusing back to the dialysis bag upon the establishment of equilibrium. The dialysate was also assayed for peroxidases activity and protein concentrations while the remaining sample was stored at -10°C.
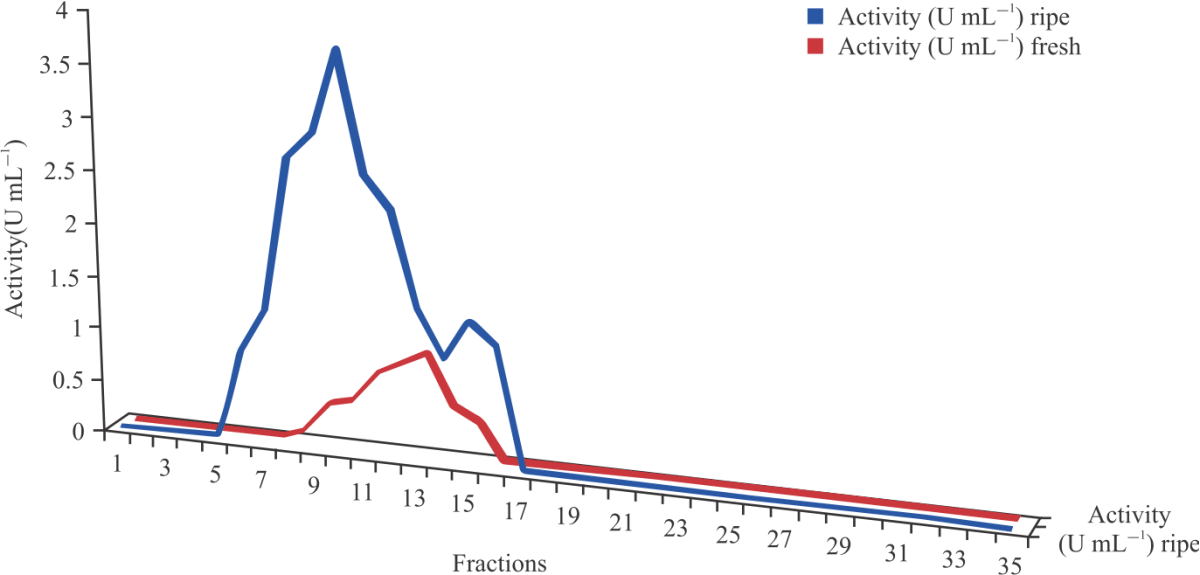
Gel filtration chromatography: Partially purified peroxidases enzyme (10 mL) obtained from dialysis were subjected to gel filtration chromatography using Sephadex-G-75. The columns were packed to the height of 17 cm in a glass column with an internal diameter of 1.5 cm. The sodium phosphate buffer pH 7.0 (0.05 M) was used to wash down the gel bed until equilibration, (pH of eluted sodium phosphate equals 7.0) and allowed to settle under gravity. After equilibration, the enzyme sample was introduced on top of the column in a slanted manner using a glass rod and eluted with the same phosphate buffer. A total of 35 fractions of 2 mL each were collected at drop rate of 2 mL/10 min constant drop rate and both the enzyme activity as well as the protein content were determined for each fraction separately, as stated in the previous sections above23,24.
Characterization of the peroxidase enzymes
Effect of pH change on peroxidase activity: The optimum pH value of peroxidases activity was determined using the method of Arnnok et al.27, with little modification by assaying enzymes activity at different pH level. The assay was performed by using sodium acetate buffer (0.05 M) at pH 3.5-5.5, sodium phosphate buffer (0.05 M) at pH 6.0-7.5 and Tris- HCL buffer (0.05 M) pH 8.0-9.0 at 0.5 intervals. Peroxidase activities were assayed by using O-dianisidine as a substrate and the enzyme introduced as stated earlier24.
Effect of temperature change on peroxidases activity: The optimum temperature of the peroxidases activity was determined by assaying enzyme activity at different temperatures from 10 to 90°C (interval of 5°C) for 1 h at pH 5.5. The activities were then assayed using the method described earlier27. The temperature was controlled by using a water bath.
Kinetic studies of peroxidase enzyme: Effect of different H2O2 concentration on peroxidases activity
Different concentrations of H2O2 (1-25 mM) were prepared and used to assay for the peroxidase activity using acetate buffer pH of 5.5 at 40°C for fresh and 45°C for ripening cucumber. The final volume of the reaction solution in a quartz cuvette was 3 mL using the method described earlier27. The change in activity was plotted against the volume of H2O2 hence, the Km and Vmax were determined from Lineweaver-Burk plots.
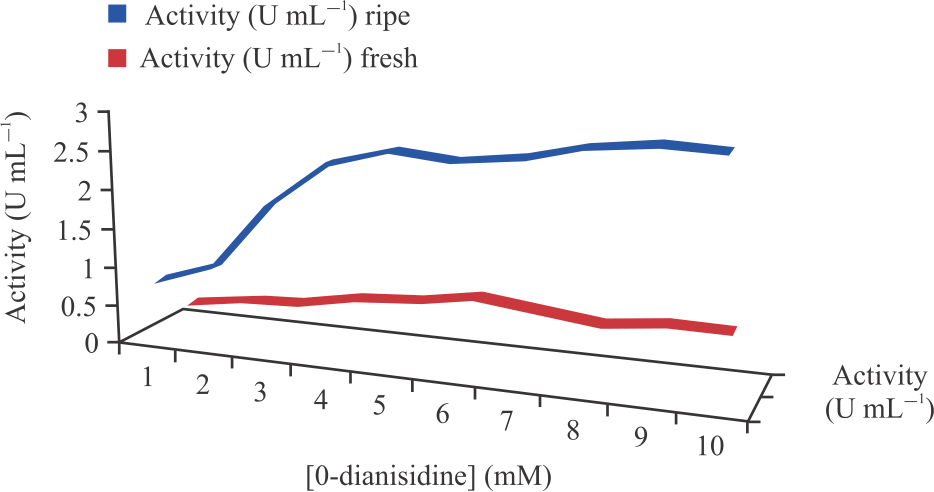
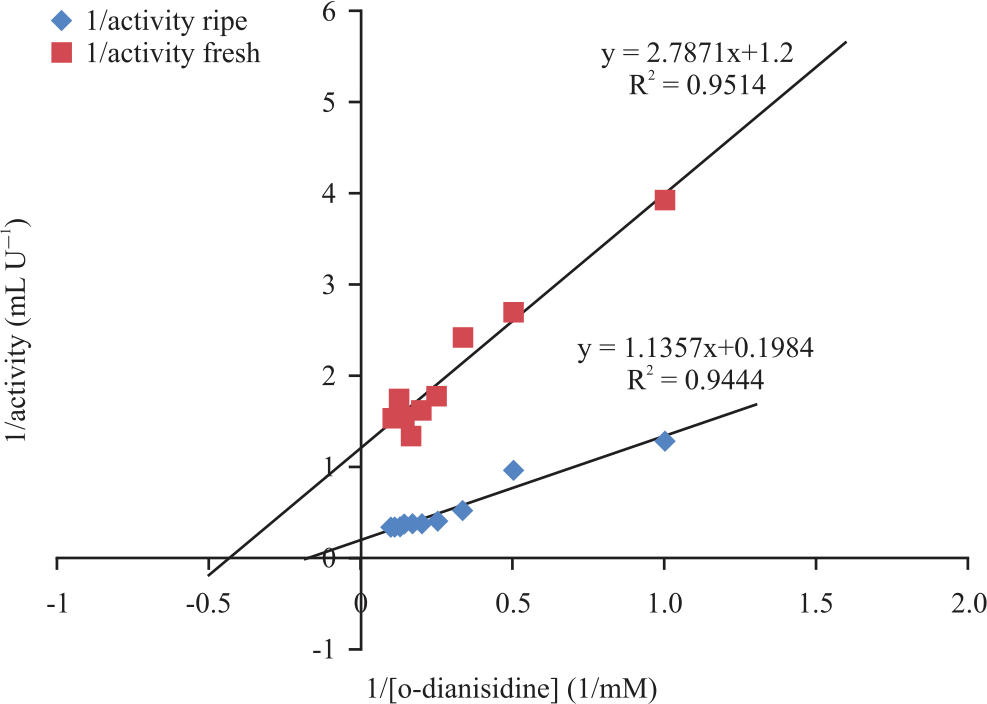
Effect of different O-dianisidine concentrations on peroxidase activity: Different concentrations of O-dianisidine (1mM-10mM) were prepared and used to assay for peroxidases activity as described above. The change in activity was calculated and plotted against change in O-dianisidine concentration; hence, Km and Vmax were assayed from Lineweaver-Burk plots.
Decolorization assays: Based on the method described by Khan and Robinson28 with slight modifications, the percentage decolorization of each dye was calculated by taking the absorbance of each dye mixture at their various optimum temperature containing 0.2 mL of the dye solution, 2.4 mL of acetate buffer (0.05 M) pH 5.5, 0.2 mL of H2O2 and 0.2 mL of the purified peroxidase, added in the order listed above. The total volume of the reaction mixture was 3 mL. The absorbance was taken immediately the enzyme was added after which it was incubated for 1 h before the final absorbance was taken. Monitoring of the oxidation was done at the maximum wavelength of the various dyes.
The percentage decolorization was calculated thus:
 |
Where Ai is initial absorbance before incubation, Af is final absorbance after incubation.
Statistical analysis: Data was expressed as mean values of three replicates. All calculations and data analyses including graphing were done using Microsoft Excel.
RESULTS
Six hundred and fifty milliliter each of the crude enzymes was extracted from both the fresh and ripening cucumber. Protein concentrations of the crude peroxidases extracted from the fresh and ripening cucumber were found to be 0.72 and 0.921 mg mL–1, respectively (Table 1, 2). Peroxidase activity of the crude enzymes which were monitored by the change in absorbance at 460 nm due to the oxidation of the O-dianisidine in the presence of H2O2 and peroxidase were found to be 0.764 and 2.124 U mL–1 for fresh and ripening cucumber respectively (Table 1). Crude peroxidases when saturated from 10-90% with ammonium sulphate were found to give the highest precipitation of the enzymes at 50 and 80% for the ripening and fresh cucumbers respectively. Hence 50 and 80% saturation (Fig. 1) were used as benchmark to precipitate 200 mL of the crude enzymes.
| Table 1: Purification table for ripening cucumber peroxidase | |||||||
| Purification step | Volume of enzyme (mL) | Protein concentration (mg mL–1) | Peroxidase activity (U mL–1) | Specific activity (U mg–1) | Total activity (U) | Purification fold | Yield (%) |
| Crude enzyme | 650 | 0.921 | 2.124 | 2.306 | 1380.6 | 1 | 100 |
| 50% (NH4)2 SO4 precipitation | 200 | 0.482 | 4.715 | 9.782 | 943 | 4.24 | 68.31 |
| Dialyzed enzyme | 25 | 0.428 | 2.464 | 5.757 | 61.6 | 2.5 | 4.46 |
| Gel filtrated enzyme | 10 | 0.143 | 3.942 | 27.566 | 39.42 | 11.95 | 2.86 |
| Table 2: Purification table for fresh cucumber peroxidase | |||||||
| Purification step | Volume of enzyme (mL) | Protein concentration (mg mL–1) | Peroxidase activity (U mL–1) | Specific activity (U mg–1) | Total activity (U) | Purification fold | Yield (%) |
| Crude enzyme | 650 | 0.72 | 0.76 | 1.11 | 496.6 | 1 | 100 |
| 80% (NH4)2 SO4 precipitation | 200 | 0.28 | 1.34 | 4.79 | 268.6 | 4.5 | 54.08 |
| Dialyzed enzyme | 25 | 0.36 | 1.41 | 3.91 | 35.2 | 3.68 | 7.08 |
| Gel filtrated enzyme | 10 | 0.23 | 0.29 | 1.25 | 2.87 | 1.18 | 0.58 |
 |
| Fig. 2: | Gel filtration chromatography profile for fresh and ripening cucumber peroxidases activity |
 |
| Fig. 3: | Protein concentration of the partially purified peroxidase |
| Table 3: Characterization table for ripening and fresh cucumber peroxidase | ||
| Results | ||
| Properties | Ripening cucumber | Fresh cucumber |
| pH | 5.5 | 5.5 |
| Optimum Temperature (°C) | 45 | 40 |
| Vmax (U mL–1) H2O2 | 1.383 | 0.69 |
| Km (mM) H2O2 | 1.355 | 3.26 |
| Vmax (U mL–1) o-dianisidine | 5.04 | 5.13 |
| Km (mM) o-dianisidine | 5.73 | 5.84 |
The protein concentrations of ammonium sulphate fractionations in both the fresh and ripening cucumber were found to be 0.28 and 0.482 mg mL–1, respectively while the peroxidase activities were found to be 1.343 and 4.715U mL–1 in both the fresh and ripening cucumber respectively as shown in Table 2.
After ammonium sulphate precipitation and dialysis, the enzyme was further purified by gel filtration chromatography using Sephadex G369-75. Figure 2 shows that peroxidase activity (U mL–1) peaks were observed at fractions 7, 8, 9, 10, 11, 12 and 15 for ripening cucumber and 10, 11, 12, 13, 14 and 15 for fresh cucumber.
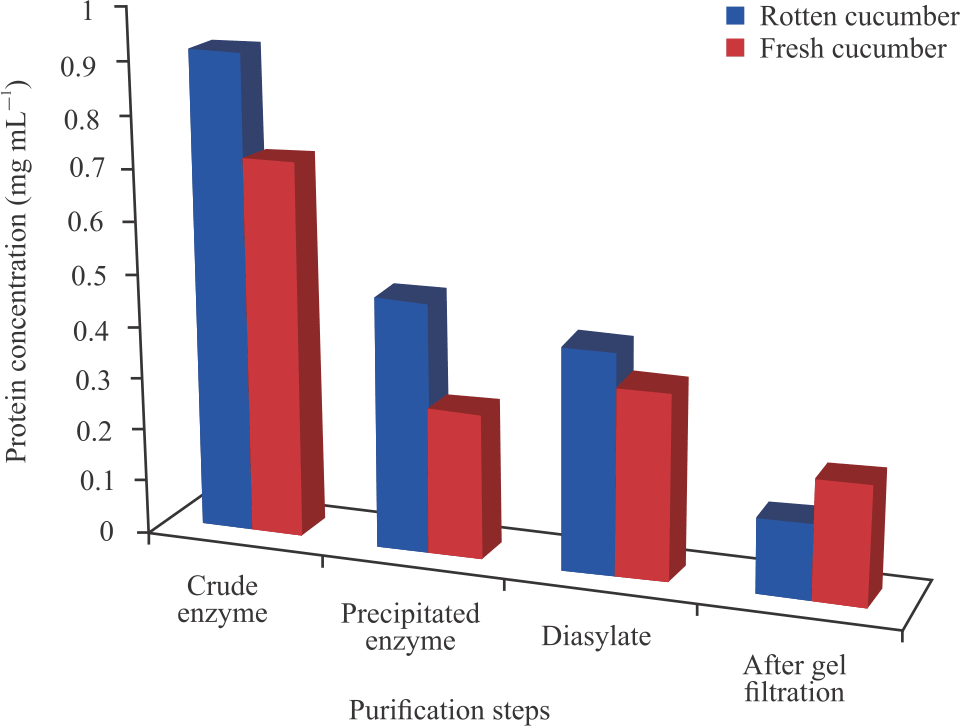
The protein concentration of the peroxidase extracted from the cucumber after dialysis and the peroxidase activity as seen in Fig. 3 and 4 were found to be 0.428 mg mL–1 and 2.464 U mL–1, respectively for ripening cucumber and 0.36 mg mL–1 and 1.41U mL–1, respectively for fresh cucumber.
Protein concentration of the crude peroxidase were found to be 0.921 and 0.72 mg mL–1 for the ripening and fresh cucumber respectively, which decreased after each purification step with the gel filtrated enzyme having the least value for both the enzyme sources.
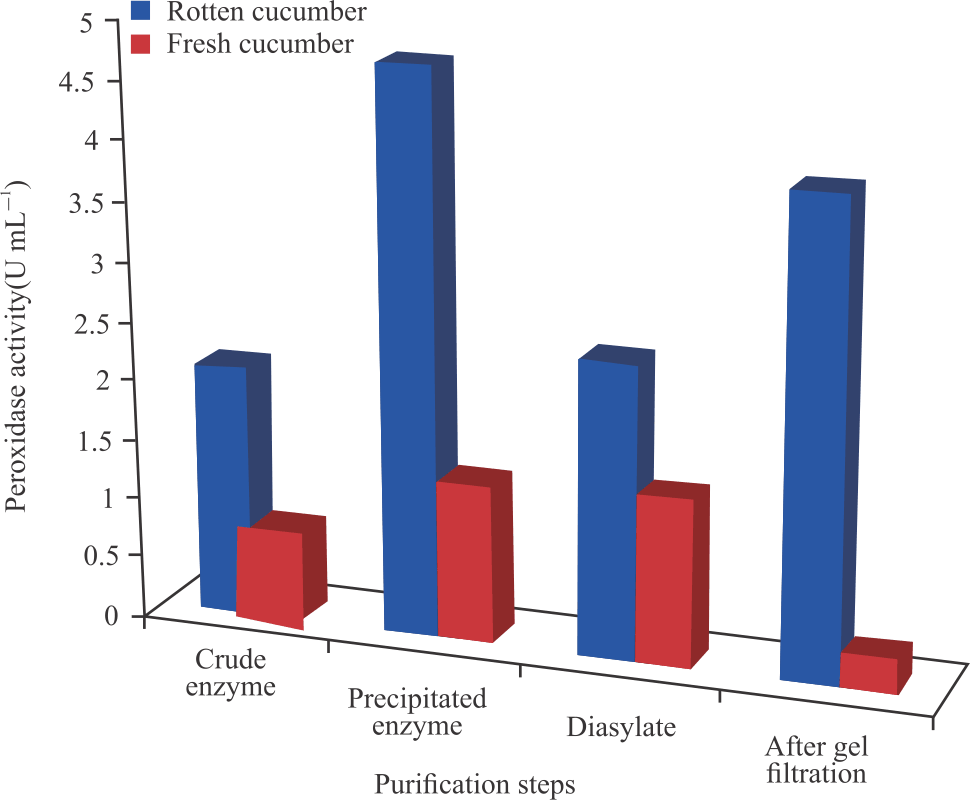
Peroxidase activity was also found to increase after each purification step with the peroxidase activity after ammonium sulphate precipitation having the highest activity for the ripening cucumber peroxidase and dialysate having highest activity for the fresh cucumber peroxidase (Fig. 3).
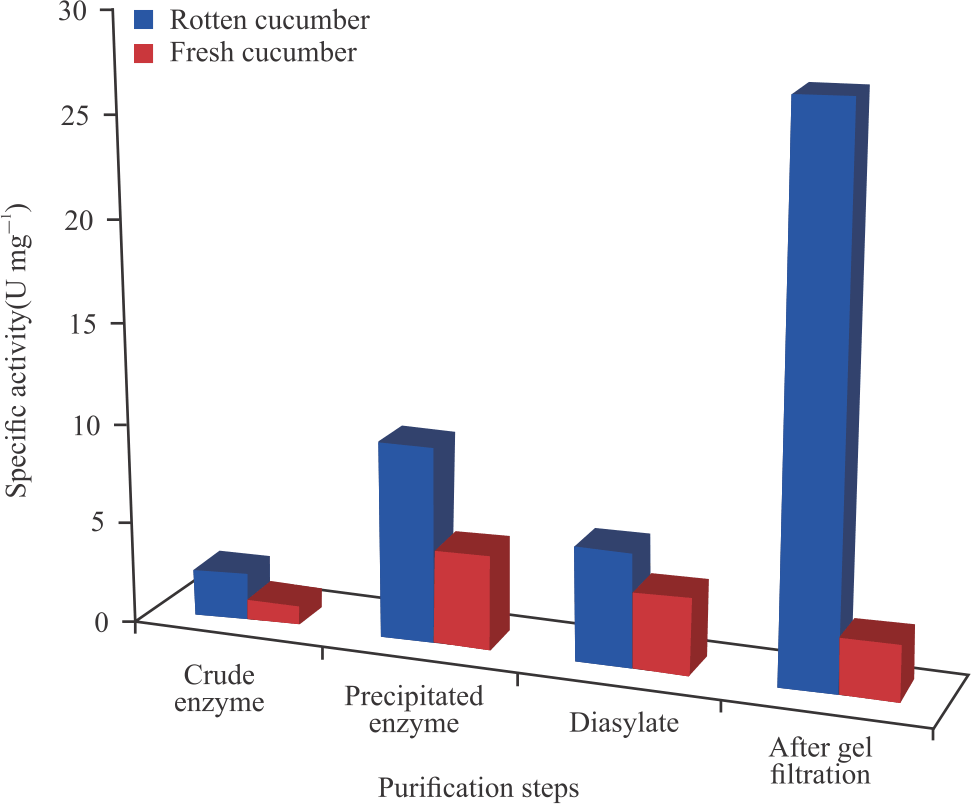
The specific activity of the crude ripening cucumber peroxidase was found to be 2.306 and 1.061 U mg–1 for fresh cucumber peroxidase. As seen in Fig. 5, the value increased in each purification step with the gel filtrated enzyme having the highest value for the ripening cucumber peroxidase and precipitated enzyme for the fresh cucumber peroxidase.
Characterization of cucumber peroxidases: Table 3 and Fig. 6 show the optimum pH for the peroxidases extracted from fresh and ripening cucumber was found to be 5.5. An increase or decrease in pH beyond or below this optimum pH showed a decline in peroxidase activity. The optimum temperature for fresh and ripening cucumber peroxidases was found to be 40 and 45°C, respectively (Table 3 and Fig. 7).
 |
| Fig. 4: | Peroxidase activity of the partially purified cucumber peroxidase |
 |
| Fig. 5: | Specific activity of the partially purified peroxidase from cucumber |
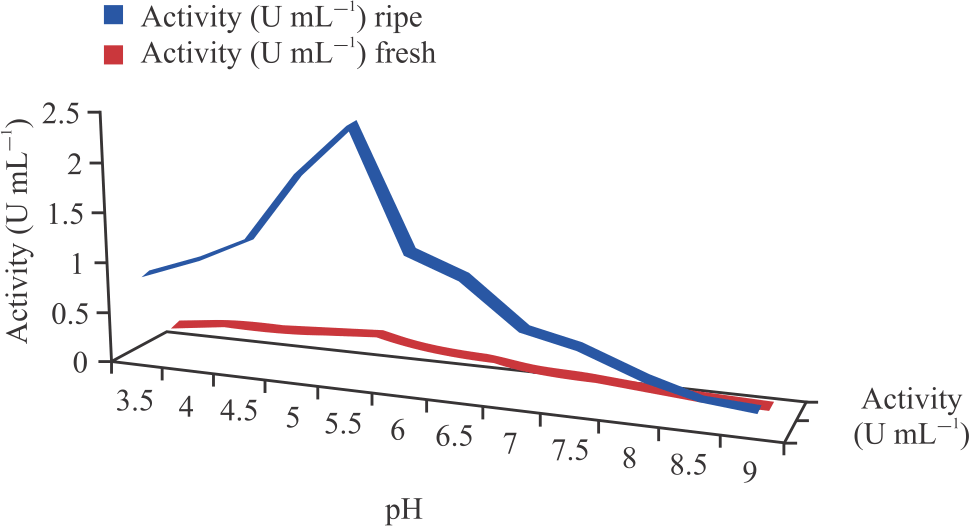 |
| Fig. 6: | Effect of pH on peroxidases activity |
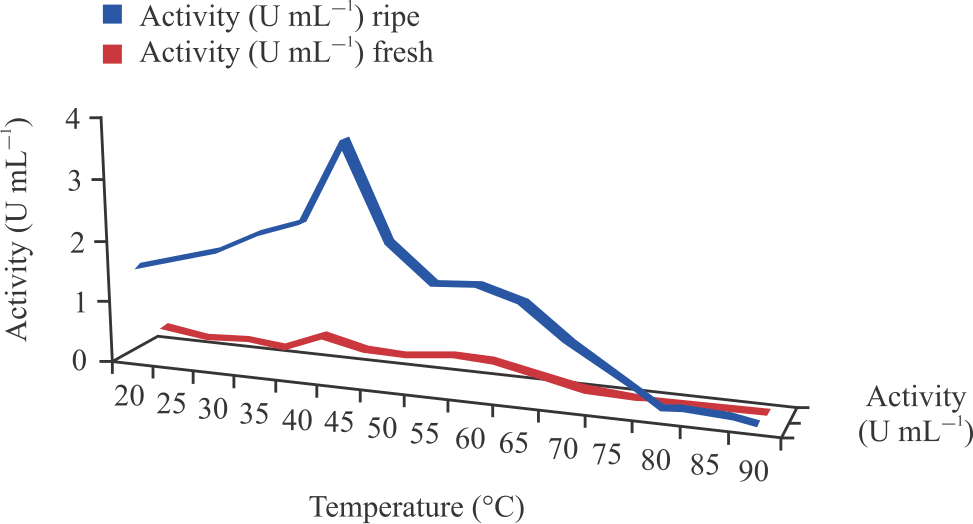 |
| Fig. 7: | Effect of temperature change on peroxidases activity |
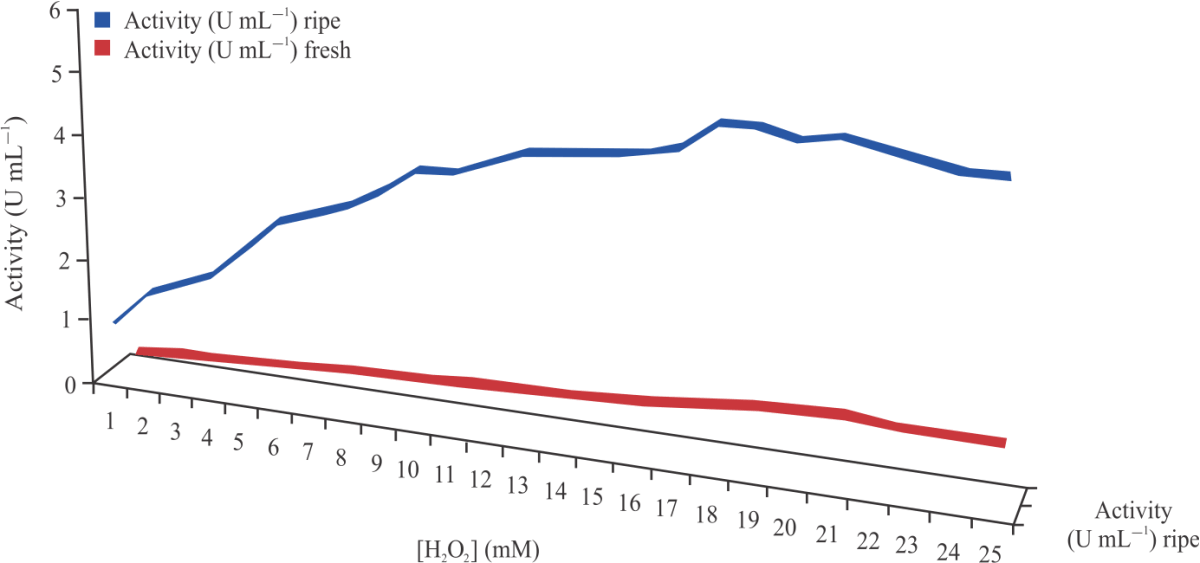 |
| Fig. 8: | Michaelis-Menten’s plots for hydrogen peroxide |
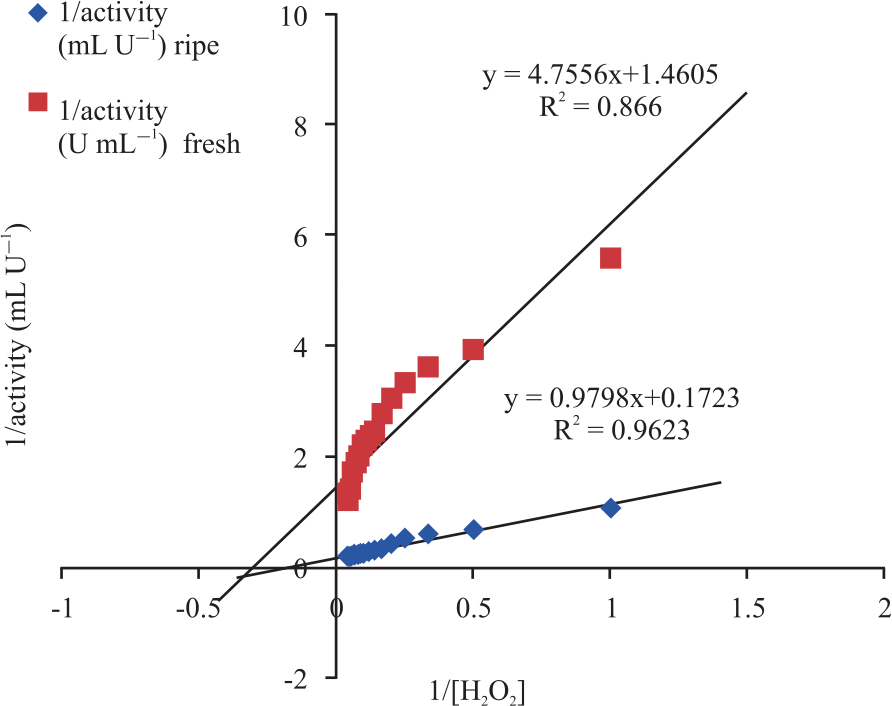 |
| Fig. 9: | Lineweaver-Burk plots 1/activity against 1/[H2O2] |
 |
| Fig. 10: | Michaelis-Mentens’s plots for o-dianisidine |
As in the case of the effect of the pH on enzyme activity, any increase or decrease in temperature beyond or below this optimum value showed a decline in peroxidase activity.
 |
| Fig. 11: | Lineweaver-Burk plots of o-dianisidine |
Peroxidase activity was also found to increase gradually at different concentrations of hydrogen peroxide until it reached its peak at 21 and 19 mM for fresh and ripening cucumber, respectively. At its peak, an increase in the concentration of the hydrogen peroxide caused no change on the peroxidases activity as seen in Fig. 8. Kinetic parameters such as Vmax and Km of the peroxidase using H2O2 were calculated from the Lineweaver-Burk plot and are as shown in Fig. 9. Peroxidase activities were found to increase gradually at different concentrations of O-dianisidine until it reached its peak at 6 mM and 9 mM for fresh and ripening cucumber respectively. At its peak, an increase in the concentration of the O-dianisidine caused no change on the peroxidases activity as shown in Fig. 10. The Vmax and Km of the peroxidase using O-dianisidine were calculated from the Lineweaver-Burk plots as shown in Fig. 11.
 |
| Fig. 12: | Decolorization (%) of dyes treated with ripening fresh cucumber peroxidase after 1 hour |
 |
| Fig. 13: | Decolorization (%) of the dyes treated with ripening cucumber peroxidase after 20 hours |
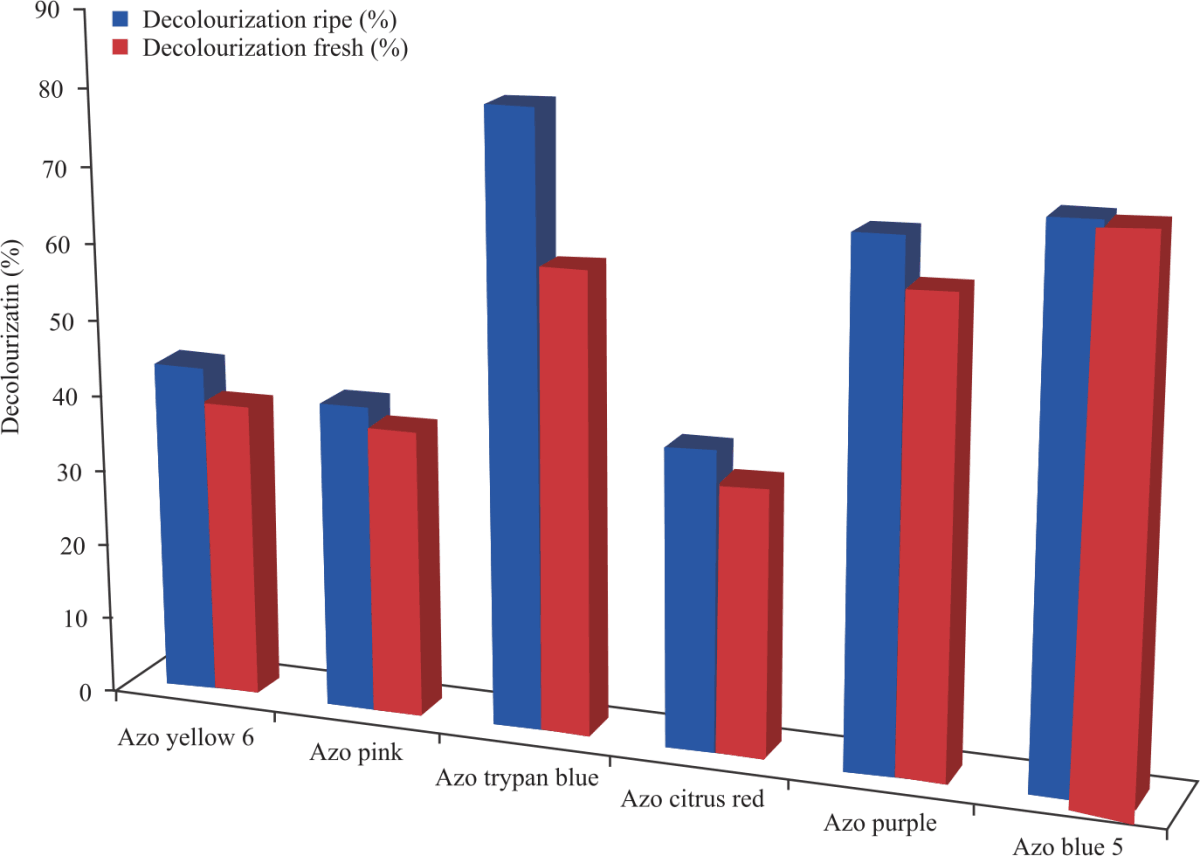
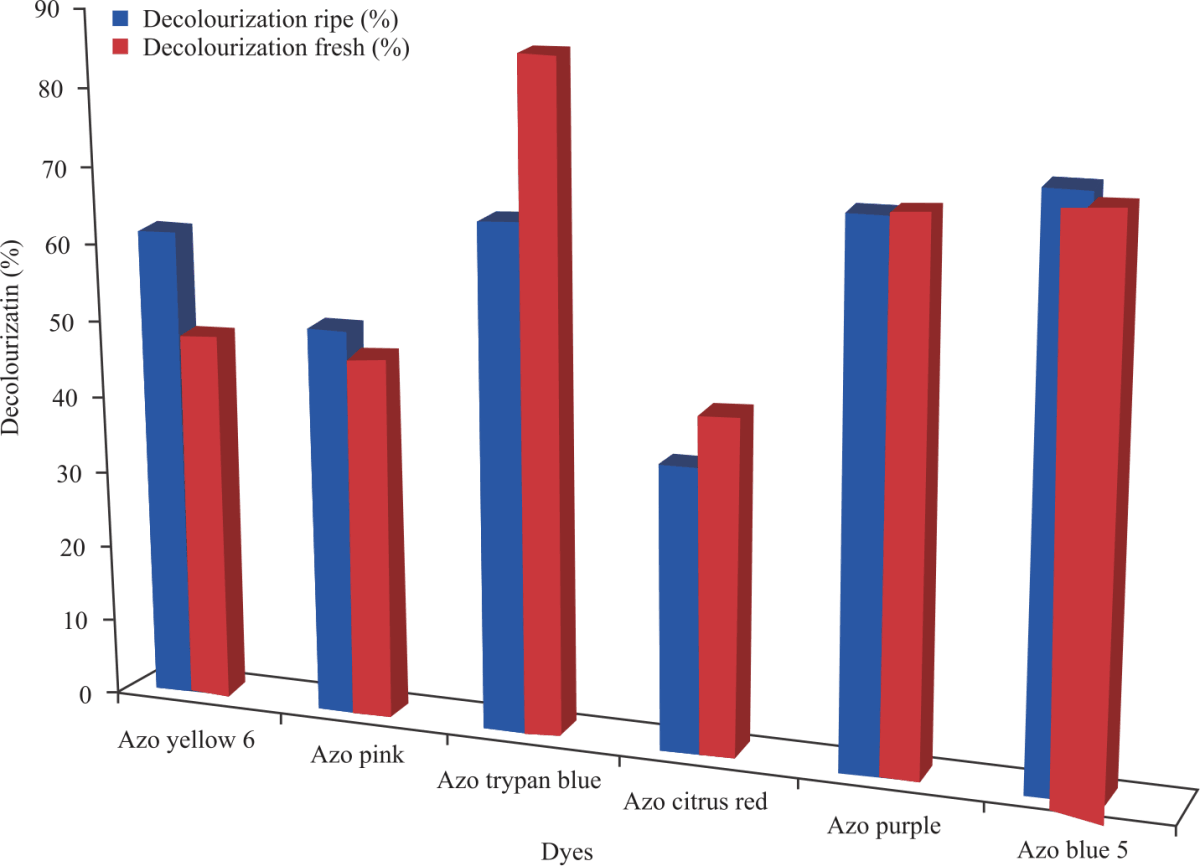
Percentage decolorization of azo dyes: Percentage decolorization of each dye was calculated at the end of 1 and 20 hrs. From the results, the dyes that had the highest % decolorization were Azo Trypan blue for ripening cucumber and Azo blue 5 for fresh cucumber as seen in Fig. 12 and 13.
DISCUSSION
This study has shown that biocatalysts in form of peroxidases could be easily extracted and purified from both fresh and ripening cucumber for possible industrial use. From the results, it could be deduced that the resultant peroxidases had characteristics that can easily be optimized for industrial purposes and various conditions at minimal costs. These qualities are highly desirable for candidates for industrial biocatalysts.
Secondly, the high yield of peroxidase enzyme from ripening cucumber which has already been discarded as not fit for human consumption is another plus as the use of ripening cucumber will reduce losses by rural farmers who could not afford modern storage facilities to increase the shelf life of their farm produce. Hence the cucumber that could not be sold for food could be sold to industries for the production of biocatalysts.
In this work, 80 and 50% (NH4)2SO4 saturation were found suitable for precipitation of cucumber peroxidases from fresh and ripening cucumbers respectively as it precipitated almost all proteins with activities of 1.34 and 4.72 U mL–1 and purification fold of 4.50 and 4.24, respectively which indicate the presence of enzymes. Significant activities were not observed below a range of 10 to 30% and peroxidases began to precipitate only when at least 40% (NH4)2SO4 precipitation in both cucumbers. Eze et al.24 reported 70% ammonium sulphate saturation for peroxidase from sorghum. Saboora et al.29 reported 85% for peroxidase from tomato, horseradish legumes and horseradish roots. Interestingly, Rehman et al.30, purifying peroxidase from various vegetables sources achieved 50% saturation ratio and Silva and colleagues31 reported 80% ammonium sulphate saturation for peroxidase from turnip. Ammonium sulphate precipitation however, is best used to optimize yield rather than to optimize purity32.
After ammonium sulphate precipitations, the enzymes were desalted by subjecting them to dialysis for 18 hours before gel filtration column chromatography. The peroxidase activity after dialysis significantly increased to 1.41 and 2.46 U mL–1 for the fresh and ripening cucumbers, respectively. The activity after gel filtration chromatography was found to be 3.94 U mL–1 with specific activity of 27.57 U mg–1 for ripening cucumber, while for fresh cucumber was 0.29 U mL–1 with a specific activity of 1.25 U mg–1. In the ripening cucumber peroxidase, the increased activity showed that the peroxidase was further purified by further removal of impurities such as other proteins and salts that interfered with the peroxidase activity, while a decrease in fresh cucumber peroxidase activity could be attributed to protein denaturation and/or other environmental factors. The results of this study slightly differed from the result recorded by Zia et al.9 with specific activity of 9.2 U mg–1 from purified apple and orange seed peroxidases.
The protein concentrations of the crude extracts were found to be 0.921 mg mL–1 which reduced upon purification to 0.482, 0.428 and 0.143 mg mL–1 for ripening cucumber, while that of the fresh cucumber was 0.72 mg mL–1 further reducing to 0.28, 0.38 and 0.23 mg mL–1 after ammonium sulphate precipitation, dialysis and gel filtration chromatography respectively. The drastic reduction in protein concentrations after dialysis could be attributed to denaturation of the enzyme arising from the environmental factors and the removal of other proteins and salt which could serve as impurities and hence, increased activity.
The partially purified peroxidases were further characterized based on the effects of pH change, temperature change and substrates (H2O2 and O-dianisidine) concentration. The optimum pH’s of the partially purified peroxidases from both fresh and ripening cucumbers was found to be 5.5 as shown in Fig. 6. This is contrary to optimum pH 6.0 reported by Bania and Mahanta33 using O-dianisidine as the hydrogen donor. Chanwun and colleagues26 reported an optimum pH of 5.0 for peroxidase from rubber trees (Hevea brasiliensis) using O-dianisidine as the H donor. The present study concurred with Nwanguma and Eze34 who reported optimum pH of 5.5 for sorghum using O-dianisidine as the H donor. Rapid significant decreases in activities were found on either the basic or acidic side of this optimum pH. These decreases might have occurred mainly via ionic alterations of the enzymes that alter the form of the enzymes and consequently the active sites31.
The optimum temperatures for the fresh and ripening cucumber peroxidases were found to be 40 and 45°C respectively as in Fig. 7. Above optimum temperatures, the peroxidases activity decreased steadily up to 70°C at which the enzymatic activities were not lost completely. Optimum temperatures for fresh and ripening cucumbers peroxidases were compared with other reports too. Singh et al.35 reported an optimum temperature of 40°C for peroxidase from apple using O-dianisidine as the hydrogen donor, Matto and Husain36, also observed an optimum temperature of 40°C for radish and tobacco peroxidases, these studies agreed with that of the fresh cucumber peroxidase. Yazdi et al.37 reported the optimum temperature of 50°C for peroxidase from cabbage. Studies on the ripening cucumber peroxidase agreed with the studies of Hu et al.38 reported an optimum Temperature of 45°C for peroxidase extracted from lettuce stem. According to the results obtained in a temperature range of 35 to 50°C, the resulting enzymes did not present a significant decrease in the enzymatic activities, which favors their use in industrial processes. From all indications, it appears that the optimum pH and Temperature for peroxidase activity varies with the enzyme source, the isoenzyme composition, the hydrogen donor substrate and the buffer used for the analysis.
Substrate concentration effects on the partially purified enzymes from ripening and fresh cucumbers were also studied using hydrogen peroxide and O-dianisidine. Hydrogen peroxide concentrations ranged from 1-25 mM while O-dianisidine concentrations were from 1-10 mM. The peroxidase activity was found to increase with the increase in hydrogen peroxide and O-dianisidine concentration until [H2O2] and [O-dianisidine] reached 19 and 8 mM for ripening cucumber peroxidase, while that of the fresh cucumber peroxidase were 21 and 6 mM respectively, after which further increase in [H2O2] and [O-dianisidine] did not cause any observable increase in the peroxidases activities.
Thongsook and Barrett39 reported Km value of 9.731 mM of hydrogen peroxide for basic peroxidase from broccoli using guaiacol as the H donor. Interestingly, Daurte-Vazquez et al.21 reported Km value of 3.7 mM for peroxidase from turnip using guaiacol as H donor. Vmax value for H2O2 can be compared with those obtained from acidic, basic and neutral peroxidase from broccoli which was 5.7, 11.7 and 438 mM/min, respectively using guaiacol as the H donor39.
Application of the partially purified ripening and fresh cucumber peroxidases were also evaluated. It has been shown that plant peroxidases are efficient for synthetic dyes decolorization and degradation. It is explicitly observed by the decrease in concentration and colour disappearance in the dye solutions under study.
This study evaluated the ability of peroxidases extracted from ripening and fresh cucumbers to decolorize 6 azo dyes. Azo Trypan Blue had the highest percentage decolorization of 80.4% followed by Azo Blue 5 and Azo Purple with 71.3 and 67.8% decolorization respectively after a contact time of 1 h. On the other hand, for fresh cucumber peroxidase, Azo Blue 5 had the highest percentage decolorization of 70.8% followed by Azo purple and Azo Trypan Blue with 61.6 and 60.6% respectively after contact time of 1 h which the percentage decolorizations later increased after a contact time of 20 hrs with the enzyme. Mugdha and Usha40 reported about 70% decolorization of azo yellow 12 was obtained due to HRP-catalyzed reaction at pH of 4 after contact time of 1 h.
CONCLUSION
In conclusion, ripening cucumber which is considered as waste and usually thrown away or composted has yielded peroxidase enzyme with industrial potentials for utilization both in the industries and the environment. In as much as fresh cucumber showed good potential also, the wastage arising from post-harvest losses and low sales resulting from bumper harvests and market glut could also be minimized as excess products which are not consumed as food could be channeled to the industries for use as a veritable source of biocatalysts. The results also indicated that the purified peroxidase has wide substrate specificity over a wide range of pH and temperature. Therefore, fresh and ripening cucumbers are a potential source of peroxidase for bioanalytical or biotechnological applications including pollution remediation at a lower cost than other peroxidases such as horseradish peroxidase.
SIGNIFICANCE STATEMENT
This study discovered that an alternative source of peroxidase could be ripening cucumber instead of the usual horseradish peroxidase. More research, however, is needed to fully characterize and elucidate this enzyme and compare its activity with the horseradish peroxidase. Further research should also be undertaken to find the enzyme characteristics when immobilized for industrial and environmental management purposes.
REFERENCES
- Mansour, H.B., I. Houas, F. Montassar, K. Ghedira, D. Barillier, R. Mosrati and L. Chekir-Ghedira, 2012. Alteration of in vitro and acute in vivo toxicity of textile dyeing wastewater after chemical and biological remediation. Environ. Sci. Pollut. Res. Int., 19: 2634-2643.
CrossRefDirect Link - Dos Santos, A.B., F.J. Cervantes and J.B. van Lier, 2007. Review paper on current technologies for decolourisation of textile wastewaters: Perspectives for anaerobic biotechnology. Bioresour. Technol., 98: 2369-2385.
CrossRefDirect Link - Ogugbue, C.J. and T. Sawidis, 2011. Bioremediation and detoxification of synthetic wastewater containing triarylmethane dyes by Aeromonas hydrophila isolated from industrial effluent. Biotechnol. Res. Int., Vol. 2011.
CrossRef - Melgoza, R.M., A. Cruz and G. Bultron, 2004. Anaerobic/aerobic treatment of colorants present in textile effluents. Water Sci. Technol., 50: 149-155.
CrossRefDirect Link - Ferraz, J.M., C.H. Del Menezzi, D.E. Teixeira and S.A. Martins, 2011. Effects of treatment of coir fiber and cement/fiber ratio on properties of cement-bonded composites. BioResources, 6: 3481-3492.
Direct Link - Firmino, P.I.M., M.E.R. da Silva, F.J. Cervantes and A.B. dos Santos, 2010. Colour removal of dyes from synthetic and real textile wastewaters in one- and two-stage anaerobic systems. Bioresour. Technol., 101: 7773-7779.
CrossRefPubMedDirect Link - Khehra, M.S., H.S. Saini, D.K. Sharma, B.S. Chadha and S.S. Chimni, 2005. Comparative studies on potential of consortium and constituent pure bacterial isolates to decolorize azo dyes. Water Res., 39: 5135-5141.
CrossRefDirect Link - Paquin, D.G., W.H. Sun, C.S. Tang and Q.X. Li, 2006. A phytoremediation study: Selection of tropical and other vascular plants for decolorization of Poly R‐478 dye. Remediation: J. Environ. Cleanup Costs Technol. Tech., 16: 97-107.
CrossRefDirect Link - Zia, M.A., M. Kousar, I. Ahmed, H.M.N. Iqbal and R.Z. Abbas, 2011. Comparative study of peroxidase purification from apple and orange seeds. Afr. J. Biotechnol., 10: 6300-6303.
Direct Link - Sakharov, I.Y., J.L. Castillo, J.C. Areza and I.Y. Galaev, 2000. Purification and stability of peroxidase of African oil palm Elaies guineensis. Bioseparation, 9: 125-132.
CrossRefDirect Link - Hidalgo-Cuadrado, N., P. Pérez-Galende, T. Manzano, C.D. De Maria, V.L. Shnyrov and M.G. Roig, 2012. Screening of postharvest agricultural wastes as alternative sources of peroxidases: Characterization and kinetics of a novel peroxidase from lentil (Lens culinaris L.) stubble. J. Agric. Food Chem., 60: 4765-4772.
CrossRefDirect Link - Floris, G., R. Medda and A. Rinaldi, 1984. Peroxidase from Ipomoea batatas seedlings: Purification and properties. Phytochemistry, 23: 1527-1529.
CrossRefDirect Link - Harris, D.L. and G.H. Loew, 1996. Identification of putative peroxide intermediates of peroxidases by electronic structure and spectra calculations. J. Am. Chem. Soc., 118: 10588-10594.
CrossRefDirect Link - Zia, M.A., Khalil-ur-Rehman, M.K. Saeed, A. Ahmed and A. Ghaffar, 2001. Partial purification of peroxidase from tomato. J. Med. Sci., 1: 404-406.
CrossRefDirect Link - Azevedo, A.M., V.C. Martins, D.M.F. Prazeres, V. Vojinovic, J.M.S. Cabral and L.P. Fonseca, 2003. Horseradish peroxidase: A valuable tool in biotechnology. Biotechnol. Annu. Rev., 9: 199-247.
CrossRefDirect Link - Pandey, V.P., M. Awasthi, S. Singh, S. Tiwari and U.N. Dwivedi, 2017. A comprehensive review on function and application of plant peroxidases. Biochem. Anal. Biochem., Vol. 6, No. 1.
CrossRefDirect Link - Adams, J.B., 1997. Regeneration and the kinetics of peroxidase inactivation. Food Chem., 60: 201-206.
CrossRefDirect Link - Sergeyeva, T.A., N.V. Lavrik, A.E. Rachkov, Z.I. Kazantseva, S.A. Piletsky and A.V. El'skaya, 1999. Hydrogen peroxide-sensitive enzyme sensor based on phthalocyanine thin film. Anal. Chim. Acta, 391: 289-297.
CrossRefDirect Link - Jia, J., B. Wang, A. Wu, G. Cheng, Z. Li and S. Dong, 2002. A method to construct a third-generation horseradish peroxidase biosensor: Self-assembling gold nanoparticles to three-dimensional sol-gel network. Anal. Chem., 74: 2217-2223.
CrossRefDirect Link - Dec, J. and J.M. Bollag, 1994. Use of plant material for the decontamination of water polluted with phenols. Biotechnol. Bioeng., 44: 1132-1139.
CrossRefPubMedDirect Link - Duarte-Vazquez, M.A., B.E. Garcia-Almendarez, C. Regalado and J.R. Whitaker, 2001. Purification and properties of a neutral peroxidase isozyme from turnip (Brassica napus L. Var. purple top white globe) roots. J. Agric. Food Chem., 49: 4450-4456.
CrossRefDirect Link - Geng, Z., K.J. Rao, A.S. Bassi, M. Gijzen and N. Krishnamoorthy, 2001. Investigation of biocatalytic properties of soybean seed hull peroxidase. Catal. Today, 64: 233-238.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Khurshid, S., M.Z. Qureshi, A. Ibrahim, Z. Nawaz, A. Sajid and M. Ikram, 2012. Production and purification of horseradish peroxidase in Pakistan. Int. J. Phys. Sci., 7: 2706-2712.
Direct Link - Chanwun, T., N. Muhamad, N. Chirapongsatonkul and N. Churngchow, 2013. Hevea brasiliensis cell suspension peroxidase: Purification, characterization and application for dye decolorization. AMB Express, Vol. 3, No. 1.
CrossRefDirect Link - Khan, A.A. and D.S. Robinson, 1994. 1994. Hydrogen donor specificity of mango isoperoxidases. Food Chem., 49: 407-410.
CrossRefDirect Link - Saboora, A., L. Parsiavash and Z. Moosavi-Nejad, 2012. Purification and kinetic properties of guaiacol peroxidase in turnip (Brassica napus var. okapi) root during different growth stages. Progress Biol. Sci., 2: 76-86.
CrossRefDirect Link - Khalil-ur-Rehman, M. Yaqub, M.A. Sheikh and M. Arshadt, 1999. Extraction and evaluation of peroxidases from various vegetable sources. Int. J. Agric. Biol., 13: 170-173.
Direct Link - Silva, M.C., J.A. Torres, A.D. Corrêa, A.M.B. Junqueira, M.T.P. Amorim and C.D. dos Santos, 2012. Obtention of plant peroxidase and its potential for the decolorization of the reactive dye remazol turquoise G 133%. Water Sci. Technol., 65: 669-675.
CrossRefDirect Link - Ward, W.W. and G. Swiatek, 2009. Protein purification. Curr. Anal. Chem., 5: 85-105.
CrossRefDirect Link - Bania, I. and R. Mahanta, 2012. Evaluation of peroxidases from various plant sources. Int. J. Scient. Res. Public., 2: 2250-2253.
Direct Link - Nwanguma, B.C. and M.O. Eze, 1995. Heat sensitivity, optimum pH and changes in activity of sorghum peroxidase during malting and mashing. J. Inst. Brewing, 101: 275-276.
CrossRefDirect Link - Matto, M. and Q. Husain, 2007. Decolorization of direct dyes by salt fractionated turnip proteins enhanced in the presence of hydrogen peroxide and redox mediators. Chemosphere, 69: 338-345.
CrossRefDirect Link - Yazdi, M.T., S. Khaleghparast and H.R. Monsef, 2002. Purification and some partial characterization of peroxidase isoenzyme from Brassica oleracea capitata L. J. Sci. Islamic Republic Iran, 13: 107-112.
Direct Link - Hu, Y., J. Wu, P. Luo and Y. Mo, 2012. Purification and partial characterization of peroxidase from lettuce stems. Afr. J. Biotechnol., 11: 2752-2756.
Direct Link - Thongsook, T. and D.M. Barrett, 2005. Purification and partial characterization of broccoli (Brassica oleracea Var. Italica) peroxidases. J. Agric. Food Chem., 53: 3206-3214.
Direct Link