
R. Sunkara
Nutrition and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal, Alabama
M. Verghese
Nutrition and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal, Alabama
L.T. Walker
Nutrition and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal, Alabama
L. Shackelford
Nutrition and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal, Alabama
Research Journal of Phytochemistry
Year: 2009 | Volume: 3 | Issue: 2 | Page No.: 25-34







ABSTRACT
The present study investigated the effect of cranberries on development of colon tumors induced by azoxymethane in Fisher 344 male rats. Fifty five rats were divided into five groups and fed with control (AIN 93) or treatment diets: cranberry meal (5, 10%) cranberry juice (2.5, 5%). Two AOM (16 mg kg-1 b.wt.) injections were given weekly for 2 weeks for induction of colon tumors. At 45 weeks of age, all rats were killed and colons were evaluated for tumor incidence, size of tumor and tumor multiplicity. Selected hepatic phase 1 (CYP2E1), phase 11 (GST) and antioxidative enzyme (catalase and SOD) activities were determined. Tumor size and tumors/tumor bearing rat were higher (p≤0.05) in the control group. Number of tumors was lower in cranberry fed rats compared to control. Administration of cranberry to rats increased (p<0.05) hepatic enzyme activities by 1.2-3.7 fold compared to control fed rats. These results indicate that feeding cranberry (meal and juice) inhibited colon tumors induced by AOM and enhanced the activity of hepatic enzymes.
PDF Abstract XML References Citation
How to cite this article
R. Sunkara, M. Verghese, L.T. Walker and L. Shackelford, 2009. Suppression of Colon Cancer Development in an Azoxymethane-Fisher 344 Rat Model by Cranberry. Research Journal of Phytochemistry, 3: 25-34.
URL: https://scialert.net/abstract/?doi=rjphyto.2009.25.34
URL: https://scialert.net/abstract/?doi=rjphyto.2009.25.34
INTRODUCTION
Colon cancer is the third most common cancer (108, 070) and second leading cause of death (49, 960) in the US (ACS, 2008). High morbidity related to cancer increases health care costs and economic burden. Thus prevention of cancer has become a prime concern for researchers. Dietary modification is one of the alternative approaches for primary prevention of cancer. It has been hypothesized that components present in certain foods may offer protection against many diseases and much research has been focused in identifying these chemopreventive compounds (Kassie et al., 2002; Fresco et al., 2006; Howells et al., 2007). However, some of these isolated agents do not yield the expected beneficial effects against diseases as they exhibited different properties at non-physiological dose (Hennekens et al., 1996; Omenn et al., 1996). The protective effect of whole foods, vegetables and fruits consumption might be due to additive or synergistic interaction of various bioactive agents.
Cranberry (Vaccinium macrocarpon Ait Ericaceae) is a native fruit of North America and has proven to have many health benefits. Prevention of urinary tract infection by the consumption of cranberry is well documented (Kandil et al., 2002). The active components present in cranberry prevent adhesion of bacteria to urinary and gastro intestinal tract and hydroxyapatite in teeth. The benefits of cranberry are attributed to the phytochemicals such as phenolics, anthocyanins, flavonols, proanthocyanidins, organic acids and fiber. Phytochemicals present in cranberry are found to contain antioxidative and in vitro anticancer properties. Extracts of cranberry showed higher antiproliferative activity (Seeram et al., 2004), reduced tumor growth (Murphy et al., 2003) and induced apoptosis (Volate et al., 2005) and exhibited anti-angiogenic and anti-inflammatory properties (Atalay et al., 2003; Bagchi et al., 2004; Roy et al., 2002) in many cell lines.
Understanding the underlying mechanisms of action by chemopreventive agents in reducing tumors is important for the identification of potential agents against colon cancer. Specific components of foods offer protection against cancer through various mechanisms: antioxidant activity and/or trapping of oxygen radicals; induction of drug metabolizing and detoxifying enzymes, modulation of signal transduction, cell proliferation and apoptosis (Shimizu and Weinstein, 2005). Protective agents for colon cancer were characterized based on their potential to modulate biotransformation enzymes (Phase1 and II) (Van Iersel et al., 1999; Paolini et al., 1996). The chemical carcinogen used in this study was Azoxymethane (AOM), which is metabolized in the liver by phase 1 (CYP2E1) enzyme. Phase I enzymes are generally involved in activating the carcinogen by exposing the functional groups and formation of reactive electrophiles. Phase II enzymes (GST, QR, UDP Glucaronidase) facilitate conjugation of specific substrates to increase water solubility and thereby facilitate elimination. Antioxidative enzymes scavenge free radicals formed during metabolism of carcinogen.
Administration of nutritional and chemopreventive agents inhibited colon carcinogenesis at different stages using the AOM-F 344 rat model (Reddy and Maruyama, 1986; Kawamori et al., 1999; Reddy, 2000). Compounds that appear to be effective in short-term studies must further be examined in long term experiments for further conclusive protective effects. Analysis of tumor data in an end point tumor model provides a definitive picture of the chemopreventive potential of agents as there are many mechanisms by which these compounds may affect tumorigenesis. Our earlier study showed that feeding cranberry for 13 weeks reduced AOM-induced colonic ACF in F344 rats (Sunkara et al., 2008). Therefore, the objective of the present study was to determine the modulatory effects of cranberry on tumor formation in F 344 male rats induced by AOM. We also examined the effect of cranberry on selected hepatic enzymes.
MATERIALS AND METHODS
Animals, Chemicals and Diets
Fifty five fisher 344 male rats (Harlan, IN) at 3 week age were maintained in a temperature (21°C) and humidity (50%) controlled environment with a 12 h of light and dark cycles. The study was conducted in the Small Animal Laboratory in the Department of Food and Animal Sciences, Alabama A and M University (2007). Following 1 week adaptation, rats were divided into 5 different groups (11 rats/group). Control group received AIN93-G/M diets and the treatment groups received cranberry in the form of meal (5 and 10%) juice (2.5 and 5%). Modifications (isocaloric) were made to the treatment diets based on AIN 93 G and M diet. Feed and fluids were given ad libitum during the experimental period. Bi-weekly body weights and daily feed and fluid intakes were monitored. Cranberry meal and juice were prepared as described earlier (Sunkara et al., 2008). For the induction of colon tumors, azoxymethane (s/c) was administered to the rats twice at seventh and eighth week of age @ 16 mg kg-1 b.wt. At 46 weeks, all rats were killed by CO2 asphyxiation. The colons of rats from each group were removed, opened longitudinally and flushed with potassium phosphate buffer (0.1 M, pH 7.2) and prepared for examination. Total number of tumors, tumor size, location and tumors per tumor bearing rat ratio (T/TBR) were determined. Weight of cecal tissue and pH of the contents were noted.
Preparation of Liver for Enzyme Analysis
Liver was excised from rats, washed with 1.15% ice-cold potassium chloride and blotted. The samples were minced, homogenized and centrifuged at 10,000 g for 30 min. An aliquot of supernatant was stored for the analysis of catalase and SOD activities. The supernatant was further centrifuged at 100,000 g for 60 min under refrigeration. The microsomal pellet was washed, mixed with an equal volume of homogenization buffer and used for the analysis of CYP2E1 and GST activities.
Determination of Enzyme Activities
The hydroxylation of p-nitrophenol (PNP) to 4-nitrocatechol (4-NC) by CYP2E1 was determined spectrophotometrically according to Koop (1997). Reaction mixtures contained 0.2 mM PNP, 50 mM Tris-HCl, 25 mM MgCl2, (pH 7.4) and 50 μL of microsomal proteins in a total volume of 1 mL. Reactions were started by adding 50 mM NADPH and stopped by the addition of 0.6 N perchloric acid. GST activity was analyzed by measuring the conjugation of 1-chloro 2,4 dinitrobenzene (CDNB) with glutathione mediated by GST (Habig et al., 1974). Liver sample was added to the reaction mixture (phosphate buffer, glutathione and CDNB) and change in absorbance was measured for 5 min at 340 nm in a Cary 1/3 UV/VIS dual beam spectrophotometer. Liver CAT and SOD activities were analyzed by using the catalase and SOD assay kits from Caymen chemical Co (Ann Arbor, MI) according to the manufacturer’s instructions.
RESULTS
General Observations
All the animals remained healthy throughout the experimental period. Feed intake and body weights of the rats in the 5 groups did not differ throughout the experiment and no significant differences were observed at the end of the study (Table 1). Highest consumption of feed, which resulted in a higher weight gain, was observed in rats administered CJ (2.5%). Rats fed cranberry did not show any significant differences in cecal weight or pH compared to the control fed rats.
Tumor Incidence, Size and Multiplicity
Most tumors developed in the distal colon of all rats. Both proximal and distal tumor incidence was higher in rats fed the control diet compared to the rats fed cranberry. Proximal tumor incidence (%) was higher in rats fed cranberry meal (CM 5%-45.45; CM 10%-36.36) compared to those fed cranberry juice (CJ 2.5% and 5%-18.18), Whereas, distal tumor incidence (%) was higher in rats fed cranberry juice (72.72) compared to the meal fed rats. There was no dose response effect observed with feeding cranberry (meal or juice) in reduction of tumor incidence (%). Administration of cranberry as meal had a higher (45.46%) effect in reducing tumor incidence compared to cranberry juice (27.28%).
| Table 1: | Effect of cranberries on weight gain, feed intake, cecal weight and cecal pH on AOM-induced Fisher 344 male rats |
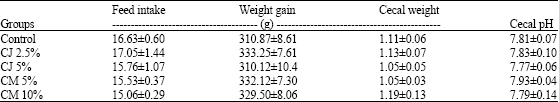 | |
| CJ: Cranberry juice, CM: Cranberry meal. Values are Means±SEM, n = 11 | |
| Table 2: | Incidence of tumors in the colon of Fisher 344 male rats fed cranberry |
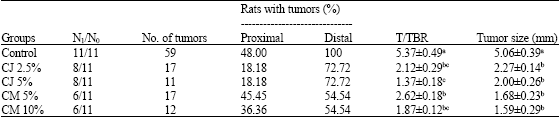 | |
| CJ: Cranberry juice, CM: Cranberry meal. N1/N0: No. of rats with tumors/No. of rats in group, T/TBR: Tumors/tumor bearing rat. Values are Mean±SEM, n = 11. abcValues not sharing a common superscript in a column are significantly different (p<0.05) with Tukey’s student range test | |
| Table 3: | Activity of selected hepatic enzymes (CYP2E1 and GST) |
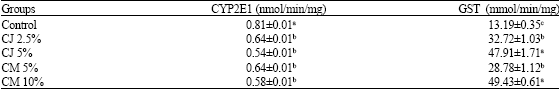 | |
| CJ: Cranberry juice, CM: Cranberry meal, CYP: Cytochrome, GST: Glutathione-S-Transferase. Values are Mean±SEM, n = 6. Values not sharing a common superscript in a column are significantly different (p<0.05) with Tukey’s student range test | |
Rats fed cranberry had significantly (p<0.05) lower T/TBR and total number of tumors compared to the control fed rats (Table 2). Among the treatment groups, number of rats with tumors/group was higher (8) in the cranberry juice fed group compared to the meal fed (6) group. Tumor numbers were highest in rats fed the control diet (59). Similar number of tumors were observed in rats fed cranberry at lower concentrations (CJ 2.5 and CM 5%- 17) and higher concentrations (CJ 2.5%-11 and CM 10%-12). Highest reductions in tumor multiplicity or T/TBR were observed in rats fed CJ 5% (74%) followed by CM 10% (62.47%), CJ 2.5% (60.22%) and CM 5% (46.90%). Rats fed higher concentrations of cranberry, CM 10% (1.87) and CJ 5% (1.37) had lower TBR ratios compared to their counterparts (CM 5%-2.62; CJ 2.5%-2.12), indicating a possible dose response effect.
Among the treatment groups, rats fed CM had smaller tumors compared to the rats fed CJ. Although there were no significant differences in average tumor size (mm) among the rats fed cranberry (juice and meal), they were lower significantly (p<0.05) compared to the rats fed the control diet. Reductions (%) in tumor size was highest in rats fed CM 10% (68.57), followed by CM 5% (66.79), CJ 5% (60.47), CJ 2.5% (55.13), compared to the rats fed the control diet.
Activity of Hepatic Phase I and Phase II Enzymes
CYP 2E1 enzyme activity was higher (p<0.05) (0.81±.01 nmol/min/mg) in rats fed the control diet compared to the rats fed cranberry diets (Table 3). GST activity (mmol/min/mg) was significantly (p<0.05) lower (13.19±0.35) in rats fed the control compared to the treatment diets. Among the treatment groups, highest GST activity was observed in rats fed higher concentrations of cranberry, CM 10% (49.43±0.61) and CJ 5% (47.91±1.71) compared to their counterparts (CM 5%-28.78±1.12; CJ 2.5%-32.72±1.03). GST activities were 3.74 (CM 10%), 3.63 (CJ 5%), 2.48 (CJ 2.5%) and 2.18 (CM 5%) fold higher than in control fed rats.
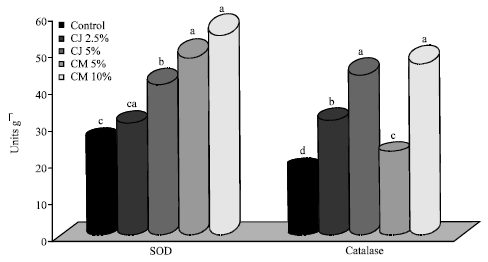 | |
| Fig. 1: | Activity of hepatic antioxidative enzymes. CJ: Cranberry juice, CM: Cranberry meal, CAT: Catalase, SOD: Superoxide dismutase. Values are means, n = 6. Bars not sharing a common superscript are significantly different (p<0.05) with Tukey’s student range test |
Activity of Antioxidative Enzymes
In order to explore the effect of antioxidant defense mechanisms in reducing the incidence, number and size of colon tumors, hepatic antioxidative enzymes (SOD and CAT) were analyzed (Fig. 1). Antioxidative enzyme activities in rats were significantly (p<0.05) increased following administration of cranberry (except CJ 2.5% in SOD activity) compared to the control. SOD activity was significantly (p<0.05) higher in the rats fed CM 10% (2.15 fold), followed by CM 5% (1.89 fold), CJ 5% (1.61 fold), compared to the rats fed the control diet. Among the treatment groups, SOD activity (units/g) was significantly (p<0.05) higher in the meal fed groups (CM 10%-58.52; CM 5%-51.54) compared to the juice fed groups (CJ 5%-43.72; CJ 2.5%-31.65). CAT activity (U/g) was significantly (p<0.05) higher in rats fed the higher concentrations of cranberry (CM 10%-49.51; CJ 5%-46.33) compared to the groups fed lower dose counterparts (CM 5%- 23.12; CJ 2.5%- 32.86). A 1.2-2.6 fold increase in CAT activity was observed in treatment groups compared to the control.
DISCUSSION
Consumption of fruits and vegetables has been implicated in reducing the risk of developing colon cancer. Plant foods consist of many bioactive (non-nutritive) and nutritive compounds which include minerals and vitamins. Including whole foods, fruits and vegetables in the diet may offer greater protection against cancer development due to the additive or synergistic action of phytochemicals (Hemaiswarya and Mukesh, 2006; Liu et al., 2002; Mertens-Talcott and Percival, 2005). Much focus has been given in recent years in the identification of fruits and vegetables that possess chemopreventive efficacy in reducing colon cancer. Dark fruits and vegetables are rich in many phytochemicals with a predominant concentration of anthocyanins. Red colored fruits such as cranberry are rich in anthocyanins and may exhibit chemopreventive effects against colon cancer.
Weight gain and feed intake did not differ among the groups fed cranberry (meal/juice) suggesting that none of the diets were either more palatable or toxic. As the cecal weight and cecal pH did not differ significantly among the groups, there may not have been any protective effects of fiber in reducing tumor incidence, by its fermentation mechanism associated with production of Short-Chain Fatty Acids (SFCA).
Tumor incidence and number of tumors were higher in control fed rats, whereas, feeding treatment diets reduced tumor incidence by 25-45% in rats. These results are comparable to the significantly lower number of tumors/rat/colon following administration of red wine polyphenols after induction of colon cancer with DMH in F344 rats (Dolara et al., 2005). Similarly, Harris et al. (2001), reported a decrease in tumor burden by 28, 42 and 75% in rats fed 2.5, 5 and 10% black raspberries compared to the control fed AOM-induced rats.
Most of the tumors in this study were located in the distal region of the rat colon in all the groups. Similar distribution of tumors has been observed in humans and rats (Holt et al., 1996) with colon cancer. Reduction in tumors/tumor bearing rat (T/TBR) was seen in rats fed cranberry. The TBR is the best indicator for assessing the efficacy of chemopreventive compounds in a chemically-induced neoplasm model as it shows the reduction of tumors in rats that developed tumors. Reduction (p<0.05) of T/TBR in the colon of rats fed cranberry meal (5 and 10%) and juice (2.5 and 5%) compared to the control fed rats indicated the tumor suppressing activity of cranberry. These results are consistent with other studies (Harris et al., 2001; Bartoli et al., 2000) where tumor multiplicity was reduced by 42, 45 and 71% after feeding 2.5, 5 and 10% black raspberries.
Rats injected with the colon carcinogen and fed the control diet might have higher degree of proliferation and lower induction of apoptosis, which led to higher tumor incidence and multiplicity. Effect of phytochemicals on cellular responses such as apoptosis and proliferation may contribute to the chemopreventive effect against colon cancer at later stages of carcinogenesis. Antiproliferative and apoptotic activities of cranberry might reduce tumor burden, size and incidence in cranberry fed groups. Hydroxycinnamates isolated from cranberries reduced the proliferation in breast, cervical and prostate cancer cell lines (Murphy et al., 2003). Anthocyanins from purple sweet potatoes, red cabbage and purple corn (at 5% in the diet), significantly reduced colorectal carcinogenesis by 48, 63 and 89%, respectively, in rats treated with 1, 2-dimethylhydrazine (Hagiwara et al., 2002). Commercial Satsuma mandarin (Citrus unshiu Marc.) juice rich in β cryptoxanthin and hesperidin, suppressed cyclin D1 overexpression and induced apoptosis in AOM-induced colon tumors male F344 rats (Tanaka et al., 2000). A monoterpene perillyl alcohol in cherry and spearmint induced apoptosis in colonic tumors and inhibited AOM-induced colon carcinogenesis (Reddy et al., 1997). A non-steroidal anti-inflammatory drug, sulindac, inhibited colon carcinogenesis by inducing apoptosis in rats (Boolbol et al., 1996).
Several plausible explanations may explain the observed effect, including the presence of various phytochemicals and their combined effect along with the fiber content of the cranberries. Polyphenols present in cranberry may also decrease tumor promotion by their anti-inflammatory activity. Several chemopreventive phytochemicals (curcumin, epigallocatechin gallate and resveratrol) have been shown to inhibit COX-2 and iNOS expression by blocking improper nuclear factor-κB activation. Whole cranberry polyphenolic extracts (Neto et al., 2006) in prostate cell line and Ursolic acid (Kondo et al., 2004) in fibrosarcoma cells inhibited the expression of extracellular matrix metaloproteins (MMP-2 and MMP-9) thereby reduced the tumor invasion and metastasis.
AOM, the colon carcinogen used in this study, is metabolized in the liver to methylazoxymethanol by CYP 2E1 enzyme and further metabolized in the liver and colon to a highly reactive electrophile, methyldiazonium ion, which can methylate cellular nucleophiles, including DNA (Fiala, 1977). Phase II detoxification enzymes such as GST and QR catalyzes the conjugation reactions to neutralize the electrophilic sites of carcinogen thus making the carcinogen more water soluble and readily excreted in bile or urine. Since the alkylation of nucleic acids is considered to be the initial step for induction of colon tumors by AOM, changes in the metabolic activation of AOM will influence the outcome of colon tumors. The decrease in the activity of phase I CYP2E1 activity and increase in hepatic phase II GST activity in this study contributes to the inhibition of colon tumors by modulating the metabolism of AOM in cranberry treated rats. Contrary to the short term study, feeding rats with cranberry for 41 wk significantly reduced phase 1 (CYP2E1) enzyme activity compared to the rats fed control diet. Constant and long-term availability of flavanoids and anthocyanins in cranberry might act as bifunctional inducers of drug metabolizing (phase I and phase II) enzymes and thereby provide benefits as cancer chemopreventing agents. Oral intake of theaflavins for four weeks, inhibited the protein level of CYP2E1 in rat intestinal microsomes (Catterall et al., 2003). Feeding citrus lemonoids, limonin and nomilin (1, 2, 5 and 10 mg day-1) for 10 days induced GST activity in liver and small intestine of wister rats in a dose dependent manner, while having no effect in induction of CYP2E1 enzyme (Kelly et al., 2003).
Reactive electrophiles developed by the conversion of AOM causes damage to DNA, proteins and other macromolecules. These free radicals also promote tissue damage and progression of cancer. One of the antioxidant defense mechanisms in the body is the enhanced activity of antioxidative enzymes such as catalase, superoxide dismutase and glutathione peroxidase. The enhanced activity (1.2-2.6 fold) of antioxidative enzymes, CAT and SOD in the liver of cranberry fed rats might be one of the mechanisms for reducing the tumor burden and size. Shih et al. (2007) reported that anthocyanins activated antioxidant response element (ARE) upstream of genes that code for antioxidative and phase II enzymes, thereby protecting cells against oxidative stress. Drinking cranberry juice at 27 and 45% for 4 months increased plasma antioxidant capacity and SOD activity in orchidectomized rats (Deyhim et al., 2007). Hepatic activities of SOD, CAT, Glutathione Peroxidase (GPx) and Glutathione Reductase (GR) were significantly increased with administration of luteolin (1.2 mg kg-1) in mice induced with AOM (Ashokkumar and Sudhandiran, 2008).
CONCLUSION
The present study showed that administration of cranberry as meal and juice significantly inhibited the development of colon tumors in F344 rats. To the best of our knowledge, no studies have been conducted to evaluate the effect of cranberry (meal and juice) on an end point colon tumor model induced by AOM in Fisher 344 rats. Although the exact mechanisms remain unclear, the modulation of colon carcinogenesis by cranberry is associated with the induction of detoxification and antioxidative enzymes. Further mechanistic and extensive studies should be conducted using cranberry to elucidate molecular mechanisms of action.
REFERENCES
- Ashokkumar, P. and G. Sudhandiran, 2008. Protective role of luteolin on the status of lipid peroxidation and antioxidant defense against azoxymethane-induced experimental colon carcinogenesis. Biomed. Pharmacother., 62: 590-597.
CrossRefPubMedDirect Link - Atalay, M., G. Gordillo, S. Roy, B. Rovin, D. Bagchi, M. Bagchi and C.K. Sen, 2003. Anti-angiogenic property of edible berry in a model of hemangioma. FEBS. Lett., 544: 252-257.
PubMedDirect Link - Bartoli, R., F. Fernandez-Banares, E. Navarro, E. Castella and J. Mane et al., 2000. Effect of olive oil on early and late events of colon carcinogenesis in rats: Modulation of arachidonic acid metabolism and local prostaglandin E2 synthesis. Gut, 46: 191-199.
CrossRefPubMedDirect Link - Boolbol, S.K., A.J. Dannenberg, A. Chadburn, C. Martucci and X.J. Guo et al., 1996. Cyclooxygenase-2 overexpression and tumor formation are blocked by sulindac in a murine model of familial adenomatous polyposis. Cancer Res., 56: 2556-2560.
PubMed - Catterall, F., N.J. McArdle, L. Mitchell, A. Papayanni, M.N. Clifford and C. Ioannides, 2003. Hepatic and intestinal cytochrome P450 and conjugase activities in rats treated with black tea theafulvins and theaflavins. Food Chem. Toxicol., 41: 1141-1147.
CrossRefPubMedDirect Link - Fiala, E.S., 1977. Investigations into the metabolism and mechanism of action of the colon carcinogens 1,2-dimethylhydrazine and azoxymethane. Cancer, 40: 2436-2445.
PubMed - Fresco, P., F. Borges, C. Diniz and M.P. Marques, 2006. New insights on the anticancer properties of dietary polyphenols. Med. Res. Rev., 26: 747-766.
CrossRefPubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Hagiwara, A., H. Yoshino, T. Ichihara, M. Kawabe and S. Tamano et al., 2002. Prevention by natural food anthocyanins, purple sweet potato color and red cabbage color, or 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine(PhIP)-associated colorectal carcinogenesis in rats initiated with 1,2-dimethylhydrazine. J. Toxicol. Sci., 27: 57-68.
CrossRefPubMedDirect Link - Harris, G.K., A. Gupta, R.G. Nines, L.A. Kresty and S.G. Habib et al., 2001. Effects of lyophilized black raspberries on azoxymethane-induced colon cancer and 8-hydroxy-2′-deoxyguanosine levels in the Fischer 344 rat. Nut. Cancer, 40: 125-133.
PubMed - Hemaiswarya, S. and D. Mukesh, 2006. Potential synergism of natural products in the treatment of cancer. Phytother. Res., 20: 239-249.
CrossRefPubMedDirect Link - Hennekens, C.H., J.E. Buring, J.E. Manson, M. Stampfer and B. Rosner et al., 1996. Lack of effect of long-term supplementation with beta carotene on the incidence of malignant neoplasms and cardiovascular disease. N. Engl. J. Med., 334: 1145-1149.
Direct Link - Holt, P.R., A.O. Mokuolu, P. Distler, T. Liu and B.S. Reddy, 1996. Regional distribution of carcinogen-induced colonic neoplasia in the rat. Nutr. Cancer, 25: 129-135.
CrossRefPubMedDirect Link - Kandil, F.E., M.A.L. Smith, R.B. Rogers, M.F. Pepin, L.L. Song, J.M. Pezzuto and D.S. Seigler, 2002. Composition of a chemopreventive proanthocyanidin-rich fraction from cranberry fruits responsible for the inhibition of 12-O tetradecanoyl phorbol-13-acetate (TPA)-induced ornithine decarboxylase (ODC) activity. J. Agric. Food Chem., 50: 1063-1069.
CrossRefPubMedDirect Link - Kassie, F., S. Rabot, M. Uhl, W. Huber and H.M. Qin et al., 2002. Chemoprotective effects of garden cress (Lepidium sativum) and its constituents towards 2-amino-3-methyl-imidazo[4,5-f]quinoline (IQ)-induced genotoxic effects and colonic preneoplastic lesions. Carcinogenesis, 23: 1155-1161.
CrossRefDirect Link - Kawamori, T., R. Lubet, V.E. Steele, G.J. Kelloff, R.B. Kaskey, C.V. Rao and B.S. Reddy, 1999. Chemopreventive effect of curcumin a naturally occurring anti-inflammatory agent, during the promotion/progression stages of colon cancer. Cancer Res., 59: 597-601.
PubMedDirect Link - Kelly, C., C. Jewell and N.M. O'Brien, 2003. The effect of dietary supplementation with the citrus limonoids, limonin and nomilin on xenobiotic-metabolizing enzymes in the liver and small intestine of the rat. Nutr. Res., 23: 681-690.
CrossRefDirect Link - Kondo, M., T.L. Lamoureaux, C.C. Neto, R.A.R. Hurta and S. Curtis et al., 2004. Proanthocyanidins, anthocyanins and triterpenoids from cranberry fruits: Antitumor activity and effects on matrix metalloproteinase expression. J. Nutr., 134: 3521S-3547S.
Direct Link - Koop, D.R., B. Klopfenstein, Y. Iimuro and R.G. Thurman, 1997. Gadolinium chloride blocks alcohol-dependent liver toxicity in rats treated chronically with intragastric alcohol despite the induction of CYP2E1. Mol. Pharmacol., 51: 944-950.
PubMed - Murphy, B.T., S.L. MacKinnon, X. Yan, G.B. Hammond, A.J. Vaisberg and C.C. Neto, 2003. Identification of triterpene hydroxycinnamates with in vitro antitumor activity from whole cranberry fruit (Vaccinium macrocarpon). J. Agric. Food Chem., 51: 3541-3545.
CrossRefPubMedDirect Link - Neto, C.C., C.G. Krueger, T.L. Lamoureaux, M. Kondo and A.J. Vaisberg et al., 2006. MALDI-TOF MS characterization of proanthocyanidins from cranberry fruit (Vaccinium macrocarpon) that inhibit tumor cell growth and matrix metalloproteinase expression in vitro. J. Sci. Food Agric., 86: 18-25.
CrossRef - Omenn, G.S., G. Goodman, M. Thornquist, S. Barnhart and J. Balmes et al., 1996. Chemoprevention of lung cancer: the beta-Carotene and Retinol Efficacy Trial (CARET) in high-risk smokers and asbestos-exposed workers. IARC. Sci. Publ., 136: 67-85.
PubMed - Paolini, M., R. Mesirca, L. Pozzetti, Sapone and G. Cantelli-Forti, 1996. Molecular non-genetic biomarkers related to fenarimol cocarcinogenesis: Organ and sex-specific CYP induction in rat. Cancer Lett., 101: 171-178.
PubMed - Reddy, B.S. and H. Maruyama, 1986. Effect of different levels of dietary corn oil and lard during the initiation phase of colon carcinogenesis in male F344 rats. J. Natl. Cancer Inst., 77: 815-822.
PubMed - Reddy, B.S., C.X. Wang, H. Samaha, R. Lubet, V. Steele, G. Kelloff and C.V. Rao, 1997. Chemoprevention of colon carcinogenesis by dietary perillyl alcohol. Cancer Res., 57: 420-425.
PubMed - Roy, S., S. Khanna, H.M. Alessio, J. Vider, D. Bagchi, M. Bagchi and C.K. Sen, 2002. Anti-angiogenic property of edible berries. Free Radic. Res., 36: 1023-1031.
CrossRefDirect Link - Seeram, N.P., L.S. Adams, M.L. Hardy and D. Heber, 2004. Total cranberry extract versus its phytochemical constituents: Antiproliferative and synergistic effects against human tumor cell lines. J. Agric. Food Chem., 52: 2512-2517.
CrossRefPubMedDirect Link - Shimizu, M. and I.B. Weinstein, 2005. Modulation of signal transduction by tea catechins and related phytochemicals. Mutat. Res., 591: 147-160.
PubMed - Sunkara, R., M. Verghese, V. Panala, R. Field, J. Boateng, L.A. Shackelford and L.T. Walker, 2008. Chemopreventive potential of cranberries on azoxymethane induced aberrant crypt foci in fisher 344 male rats. Int. J. Cancer Res., 4: 52-60.
CrossRefDirect Link - Volate, S.R., D.M. Davenport, S.J. Muga and M.J. Wargovich, 2005. Modulation of aberrant crypt foci and apoptosis by dietary herbal supplements (quercetin, curcumin, silymarin, ginseng and rutin). Carcinogenesis, 26: 1450-1456.
CrossRefPubMedDirect Link