
F. Fadakar Masouleh
Department of Fisheries Science, Faculty of Natural Resource, University of Tehran, Karaj, Iran
B. Mojazi Amiri
Department of Fisheries Science, Faculty of Natural Resource, University of Tehran, Karaj, Iran
A.R. Mirvaghefi
Department of Fisheries Science, Faculty of Natural Resource, University of Tehran, Karaj, Iran
M.A. Nemtollahi
Department of Fisheries Science, Faculty of Natural Resource, University of Tehran, Karaj, Iran
Research Journal of Environmental Toxicology
Year: 2011 | Volume: 5 | Issue: 2 | Page No.: 108-116







ABSTRACT
To study the impacts of environmentally hazardous organophosphate, Diazinon (DZN), on male reproductive system of biologically, nutritionally and economically pivotal Cyprinidae, Caspian kutums (Rutilus frisii kutum), the in vitro effects of DZN on spermatogenesis and sperm motility of maturing and mature fish investigated. The fish testes were exposed to sub-lethal concentrations of DZN, 0.01, 0.1 and 0.2 mg L-1, using in vitro tissue culture, for 3 and 6 days. In other experiment, percentage of motile spermatozoa and its total duration were measured in solutions containing 0.01, 0.1, 0.2 and 0.5 mg L-1 of DZN. Adverse effects on testes were more apparent with the increase of DZN concentration. At concentration of 0.1 and 0.2 mg L-1 of DNZ a remarkable necrotic germ cells were observed. Compared to control groups, the mean size of Spermatogonia (SPG) and Spermatocycle (SPC) significantly decreased (p<0.001) in dose-dependent manner. With the increase of DZN concentration, the number of SPCs and Spermatids (ST) decreased (p<0.001) but the number of second SPG, increased (p<0.05). With the increase of DZN concentration, percentage of motile spermatozoa and total duration of spermatozoa significantly decreased (p<0.05) but there was no significant difference between 0.01, 0.1, 0.2 mg L-1 (p>0.05). Our study supports the detrimental impacts that pollution of aquatic environment with very low levels of DZN has adverse effects on male testes and sperm quality in the Caspian kutum.
PDF Abstract XML References Citation
Received: February 27, 2011;
Accepted: April 25, 2011;
Published: June 03, 2011
How to cite this article
F. Fadakar Masouleh, B. Mojazi Amiri, A.R. Mirvaghefi and M.A. Nemtollahi, 2011. In vitro Effects of Diazinon on Male Reproductive Tissue and Sperm Motility of Caspian Kutum (Rutilus frisii kutum). Research Journal of Environmental Toxicology, 5: 108-116.
DOI: 10.3923/rjet.2011.108.116
URL: https://scialert.net/abstract/?doi=rjet.2011.108.116
DOI: 10.3923/rjet.2011.108.116
URL: https://scialert.net/abstract/?doi=rjet.2011.108.116
INTRODUCTION
Rutilus frissi kutum is one of the endemic anadromus and most important cyprinid fish living in the south coasts of Caspian Sea (the largest lake in the world). Fish spend their whole life in the sea but migrate to the rivers like Haviq, Lemir, khoshkroud, Tajan (Kavan et al., 2009) and Shiroud for spawning when they are maturing or mature in 3 or 4 year old males and 4 year old females (Yousefian and Mosavi, 2008). Caspian kutum will spend a short time in the river before spawning where they are exposed to some pollutants. It seems that many of these municipal, agricultural and industrial pollutants are suspected to have endocrine disrupting function (McLachlan, 2001). The fish, among all other vertebrates, because of their contaminated habitats may experience lifelong systemic exposure to a wide variety of Endocrine Disrupting Chemicals (EDCs) (Yang et al., 2008) and this is more important for Caspian kutums because environmental pollution is one the main reasons for decreasing in catch during the 1960s and 1970s in Iran (Valipour and Khanipour, 2006). Today a large amount of organophosphorus pesticides like DZN are used world-wide. In Iran, maximum concentration measured in some rivers just a day after use of this pesticide beside agricultural lands was 0.9-1.14 mg L-1 (Shayeghi et al., 2001; Arjmandi et al., 2010). It can cause neurotoxicity by inhibition of acetyl cholinesterase (AChe), Loosing Fish appetite (El-Sherif et al., 2009) and produce very toxic effects on birds, fish and aquatic invertebrates (USEPA, 1986; Ducolomb et al., 2009).
It is believed that DZN is one of the most harmful organophosphates that could be as EDCs and suppress reproductive activities with endogenous hormonal disruption (Dutta and Arends, 2003; Xue et al., 2005; Mlambo et al., 2009). It also could have direct effects on gonads, disrupting sexual behaviors, gametes quality and spawning process. Histology is an important tool to investigate the endocrine disrupting effects of DZN on fish. Some reports showed that during spermatogenesis a number of histopathological changes appeared in fish when they were exposed to EDCs and also it is approved that, there is a direct relationship between water quality and released sperm viability and fertilization capability (Abascal et al., 2007; Catarino et al., 2008). To evaluate sperm quality several biomarkers such as spermatocrit, pH, osmolarity and composition of seminal plasma, enzymatic activity and Adenosine Triphosphate (ATP) concentration are being used (Rurangwa et al., 2004) but sperm motility and its duration are the most well known and important factors on successful fertilization so far (Gage et al., 2004; Aral et al., 2007; Rosengrave et al., 2009). Because of kutum’s significant role in economical activities of the people living near the south coast of Caspian Sea (Lasheidani et al., 2008) and its importance in their nutrition, good quality gametes and reproduction efficiency have been considered in recent decades. This study was conducted to investigate: 1) the direct effects of DZN on Caspian kutum spermatogenesis when fish were passing last few days of maturing process and 2) the effect of this chemical on sperm motility of this unique fish.
MATERIALS AND METHODS
In vitro tissue culture: During the early period of fish migration for spawning to the river (February, 2008), four wild maturing male Caspian kutums were catched using trap net and kept in 13 ppt salinity and 12°C tanks until transport to the lab. After anesthetized using 200 ppm clove powder with sterile condition fresh testes were removed, cut into small pieces (10-15 mg) and placed in each wells of 24-well culture dishes (NUNCE, Denmark) containing 1.5 mL Leibovitz (L-15, Gibco, USA) culture medium supplemented with 0.5% bovine serum albumin fraction V (Sigma, USA), 1 mg L-1 bovine insulin (Miura et al., 2005) and 10 mM Hepes, 100 units mL-1 penicillin (Gibco, USA), 100 μg mL-1 streptomycin (Gibco, USA) and adjusted to pH 7.5. DZN was filtered by micro filter 0.2 μm (Wattman) and added with the concentration of 0.01, 0.1 and 0.2 mg L-1 to the wells and incubated in humidified air at 12°C for 3 and 6 days. The 3rd day groups were incubated separately from 6th day groups and the medium was changed after 3 days. After the culture, testicular fragments were collected and fixed in buin’s solution, then were prepared using histotechniques (Humason, 1972) and stained with hematoxylin-eosin (Pearse, 1985). Prepared sections photographed on a light microscope using a digital camera (Nikon coolpix p6000) and examined for histopathological lesions. The taken data analyzed by image tools software (3.00) for quantitative results. The number and size of cells were evaluated in four replicated views (0.3 mm2) per histological sections at 1000x magnifications and mean values were used for statistical analysis. Dead cells were omitted and only intact cells were measured in present analysis.
Sperm motility: The semen of eight wild male Caspian kutums were collected separately during their migrating into the river (March, 2009) and brought to the laboratory for analysis. Sperm motility was evaluated for total duration of motility (Alavi et al., 2004) when 99% of spermatozoa were immotile (in seconds) and the percentage of motile spermatozoa (Cosson and Linhart, 1991) after activation. To induce initiation of sperm motility, 0.5 μL of semen placed on a glass slide and then 1 μL of DZN solution with 0 (as control group), 0.01, 0.1, 0.2 and 0.5 mg L-1 diluted it. All experiments were performed in five repeated manner using a light microscope under 400 magnifications and simultaneously was recorded with digital camera. Semen was stored at 4°C during motility analysis at room temperature 15-17°C.
Statistical analysis: The results are presented as Mean±SD. The normal distribution of the data was tested using the kolmogrov-smirnov test. Statistical analysis was done by one-way Analysis of Variance (ANOVA). All statistical analyses were carried out using SPSS, 15. Dunkan’s Multiple Comparison Test was used to determine significant difference against the control.
RESULTS
In vitro tissue culture
Qualitative analysis: Morphological analysis from control groups showed that testes had a normal histological structure. In 3rd day groups, lobules contained germinal cells at all stages of spermatogenesis and in 6th day groups, clusters of ST and spermatozoa were more visible (Fig. 1a). In the 6th day DZN exposed groups, lumens showed disorganized structures compare to 3rd day and controls. In 0.2 mg L-1, necroses were clearly visible in SPCs, ST (Fig. 1b) and spermatozoa cells (Fig. 1c). In some lumens germ cells were completely disrupted and in a few sections with no distinct reason lifting of the basal membrane was distinguished.
Quantitative analysis
Size of germ cells: The size of second SPG and second SPC after exposure to different concentrations of DZN became smaller in dose dependent manner compared to controls (p<0.001, Fig. 2). The mean area of SPG in 3rd and 6th day groups were 0.444±0.07 and 0.416±0.04 mm2, respectively.
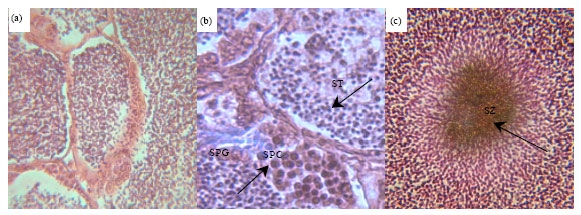 | |
| Fig. 1: | Representative histological sections of kutum testes (a) control normal clusters of spermatozoa (SZ) 400X (b) necrotic areas covering spermatocytes (SPC) and spermatids (ST) and (c) necrotic spermatozoa (arrowhead) 1000X |
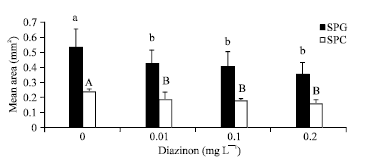 | |
| Fig. 2: | Mean area of germ cells±SD exposed to different concentrations of Diazinon (p<0.001; n = 36), SPG: Spermatogonia II, F value: 175.418***, df: 3, SPC: Spermatocyte, F value: 23.381***, df: 3 |
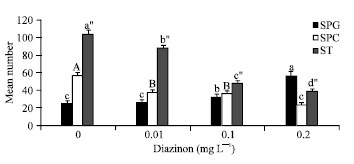 | |
| Fig. 3: | Mean numbers of germ cells±SD exposed to different concentrations of DZN (p<0.001, n = 36), SPG: Spermatogonia (II) F value: 50.427***, df: 3; SPC: Spermatocyte F value: 46.135***, df: 3, ST: spermatid F value: 54.298***, df: 3 |
The mean areas of SPC in those days were 0.198±0.01 and 0.177±0.01 mm2, respectively. There were no significant interaction between DZN and time on SPG and SPC size (p>0.05).
Number of germ cells: The mean numbers of counted SPC and ST decreased (p<0.001, Fig. 3) but the mean number of SPG increased (p<0.001) time dependently. They were 32.333±1.20 and 37.062±4.24 in 3rd and 6th day groups, respectively. The mean numbers of SPC were 46.533±3.37 and 31.083±2.04 and the mean counted number of ST were 74.458±6.76 and 65.333±3.87 in 3rd and 6th day groups, respectively. There were no significant interactions between DZN and time on cells Number (p>0.05).
Also, with the increase of DZN concentration, diameter of seminiferous tubules in testes decreased (p<0.001, Fig. 4). There were neither significant correlation between time and tubules diameter nor significant interaction between DZN and time on diameter of seminiferous tubules (p>0.05).
Sperm motility: Total duration of spermatozoa motility decreased with DZN increase but there was no statistically significant differences between 0.01, 0.1, 0.2 mg L-1 (p>0.01; Fig. 5a). Comparison of the mean motility duration of different percentages of motile spermatozoa that affected by different concentrations of DZN indicated that the observed decreased had significant correlation between DZN concentrations and decreased motility duration of 10, 40 and 80% of motile spermatozoa (p<0.05) and when spermatozoa were in higher motility (80%), their observed significant differences were more apparent than other percentages of motile spermatozoa (Fig. 5b).
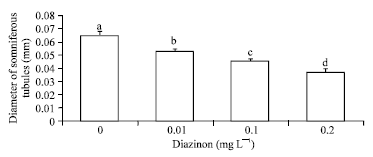 | |
| Fig. 4: | Mean diameter of seminiferous tubules±SD exposed with different concentration of Diazinon (p<0.001, n = 27), F value: 60.648***, df: 3 |
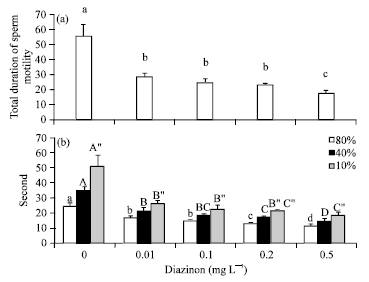 | |
| Fig. 5: | Mean total duration of spermatozoa motility±SD (a) percentage of motile spermatozoa±SD (b) exposed to different concentration of diazinon |
DISCUSSION
DZN is a harmful organophosphate that can be considered as EDCs to adversely influence reproductive activities with endogenous hormonal disruption (Dutta and Maxwell, 2003; Mlambo et al., 2009). DZN has also direct effect on gonads to make low quality gametes or harmful affecting sexual activity and spawning. Significant reduction levels of reproductive steroids in Atlantic salmon (Salmo salar) and zebrafish (Danio rerio) after exposure of sub lethal doses of DZN were reported (Moore and Wairing, 1996). In mammals, DZN and Malathion have destructive effects on enzymes involving in spermatogenesis process (Ducolomb et al., 2009). The observation of this experiment agrees with studies that have shown several adverse effects of toxic pollutants on testis structure and spermatogenesis process in rats and fish (Yamaguchi et al., 2007; Jorsaraei et al., 2010; Orlu and Gabriel, 2011). Associated with disorganized changes in testes, qualitative analysis of the histological sections indicated that at least one degenerative effect was seen in each section. With the increase of dose exposure, the adverse effects on testes were more distinctive and damage tissues with necrotic areas were obviously visible in 0.2 mg L-1. The lumens structure was disordered in some parts. Histopathological alterations observed in this study are nearly similar to those found in Mozambique tipalia (Oreochromis mossambicus) (Mlambo et al., 2009) and black goby (Gobius niger) (Louiz et al., 2009) affected by organic pollutants like DDT.
In quantitative analysis, there were significant differences between germ cell numbers and size exposed to DZN and the controls. So DZN concentrations were sufficient to cause disorganization on testes structure. In fact, the concentrations of DZN applied in this experiment were lower than LC50 values and those observed in the rivers where kutum migrated. In vivo experiment using lepomis machrochirus demonstrated the inhibitory effect of DZN on testicular development by decreasing diameter of spermatogonia, lumen and seminiferous tubules following exposure of 60 μg L-1 DZN (Dutta and Maxwell, 2003). Also, these present data shows that DZN lessened the size of germ cells in all treatments. As during the exposure, increase of DZN concentrations and also length of exposure caused decrease in SPG and SPC size, suggesting direct correlation between germ cells size, dose and length of exposure. As well as germ cells size, the number of SPCs and ST decreased, too. Large number and great size cells were related to control groups that can show DZN dose-dependent inhibitory effects on the spermatogenesis. With the increase of DZN, the number of SPGs increased because of declining or stoppage effects on cell division. The number of ST because of low SPG and SPC proliferation rate, decreased and diameter of the seminiferous tubules had direct correlation with DZN doses as well. In control groups germ cells were larger and ST were commonly recognized and also clusters of spermatozoa were obviously seen in the lumens. Damage cells were more visible in 0.1 and 0.2 mg L-1 and necrotic areas were more distinctive. In male rats injected intraperitoneally by sub lethal doses of DZN, significant reduction in seminiferous tubule size observed in 25 mg kg-1 compare to controls and the number of spermatocytes, Leydig and germinal cells were significantly decreased (Jorsaraei et al., 2010).
Toxicity of DZN for fish species depends on different absorption, inhibition of acetyl cholinesterase and detoxification (Oh et al., 1991). On the other hand sensitivity of different fish following exposure to chemicals might be sex dependent (Sole et al., 2003; Louiz et al., 2009). So sometimes testes are more sensitive than ovary to chemicals and the opposite is too. Therefore, the study of kutums sex dependency to DZN toxicity must be reviewed.
DZN caused reduction of milt producing, sex behaviors and also gene damage in male salmon fish (Cox, 2000). The data from sperm motility analysis demonstrated that, passing the time, DZN increasing has adverse effect on the percentage of motile spermatozoa. In addition, total duration of spermatozoa motility declined. Several intra cellular components such as energy contents, plasma membrane mediators for ionic exchange and axoneme structure and composition are responsible for sperm movement (Bobe and Labbe, 2010). Total duration of spermatozoa motility following exposure to distilled water was nearly 56 sec. When spermatozoa exposed to even low dose of DZN, its total duration decreased slightly but there was no statistically significant difference between 0.01, 0.1, 0.2 mg L-1 except in 0.5 mg L-1, as in this concentration, duration of 80% of motile spermatozoa declined to less than 10 sec. It is remarkable, as in Fig. 5, that spermatozoa in their greater percentage of movement are more sensitive than spermatozoa in low motility percentages because with the increase of DZN concentration they have more significant difference to control group than others in low percentages. In other words, in early seconds after initiation of motility, spermatozoa were more sensitive. These results indicate that there is relationship between DZN concentration and sperm quality with regard to motility and its total duration. Sufficient time to fertilize eggs is 15 sec after initiation of sperm cells motility (Gage et al., 2004), so each toxicant with decreasing effect on sperm movements, can have an adverse effect on fertilization efficiency.
Addition to fishes, organophosphates would have implications for humans and laboratory animals’ fertility and reproduction via adverse affecting morphology, motility and sperm viability (Pina-Guzman et al., 2005; Fattahi et al., 2009). Organophosphates effect on sperm motility by disturbing synthesis of ATP or energy pooling and significant increase in percentage of broken sperms (Okamura et al., 2009). In this study, reduced sperm motility and total duration were relevant to spermatotoxicity markers of DZN as well as reduced sperm production in testes. Not only DZN could have adverse effect on gamete quality and fertility rate (Dutta and Arends, 2003) but also its adverse effects on hatching success have been confirmed already by Hamm and Hinton (2000) and Aydin and Koprucu (2005). Nonetheless, in aquatic ecosystems, transitive levels of toxic elements like DZN act as reproductive toxicant either interfering with endocrine system, hormonal regulation and sperm production or spermatozoa activity after releasing in water environments. Along with endangered reproduction effects on male fish, DZN can affect on females too (Maxwell and Dutta, 2005). Indisputably, bad quality gametes produce less viable offspring and finally affect on fish populations. Significant effects of low levels of DZN (0.25 mg L-1) is observed on common carp (Cyprinus carpio) populations in aquatic environment (Aydin and Koprucu, 2005).
CONCLUSION
These data represent that organophosphates like DZN are testicular toxicant in male kutum, human (Bustos-Obregon and Ricardo Hartley, 2008) and other laboratory animals and it has been subjected to disturbed reproduction functions. Furthermore additional pollution studies will be necessary in order to distinguish the dysfunction of kutums reproduction system and other DZN harmful effects studies covering in vivo experiments and hormonal regulation of kutums should be done to express the exact harmful reproductive effects.
REFERENCES
- Abascal, F.J., J. Cosson and C. Fauvel, 2007. Characterization of sperm motility in sea bass: The effect of heavy metals and physicochemical variables on sperm motility. J. Fish. Biol., 70: 509-522.
CrossRef - Alavi, S.M.H., J. Cosson, M. Karami, B.M. Amiri and M.A. Akhoundzadeh, 2004. Spermatozoa motility in the Persian Sturgeon, Acipenser persicus: Effect of pH, dilution rate, ions and osmolality. Reproduction, 128: 819-828.
CrossRefDirect Link - Aral, F., E. Sahinoz and Z. Dogu, 2007. A study on the milt quality of Oncorhynchus mykiss (Walbaum, 1972) and Carasobarbus luteus (Heckel, 1843) in Ataturk Dam Lake, Southeastern Turkey. Turk. J. Fish. Aquat. Sci., 7: 41-44.
Direct Link - Arjmandi, R., M. Tavakol and M. Shayeghi, 2010. Determination of organophosphorus insecticide residues in the rice paddies. Int. J. Environ. Sci. Tech., 7: 175-182.
Direct Link - Aydin, R. and K. Koprucu, 2005. Acute toxicity of diazinon on the common carp (Cyprinus carpio) embryos and larvae. Pestic. Biochem. Phys., 82: 220-225.
CrossRef - Bustos-Obregon, E. and B. Ricardo Hartley, 2008. Ecotoxicology and testicular damage (environmental chemical pollution). A review. Int. J. Morphol., 26: 833-840.
CrossRefDirect Link - Catarino, A.I., H.N. Cabral, K. Peeters, P. Pernet, U. Punjabi and P. Dubois, 2008. Metal concentrations, sperm motility and RNA/DNA ratio in two echinoderm species from a highly contaminated fjord (The Sorfjord, Norway). Environ. Toxic. Chem., 27: 1553-1560.
CrossRef - Ducolomb, Y., E. Casas, A. Valdez, G. Gonzalez, M. Altamirano-Lozano and M. Betancourt, 2009. In vitro effect of malathion and diazinon on oocytes fertilization and embryo development in porcine. Cell. Biol. Toxicol., 25: 623-633.
CrossRef - Dutta, H.M. and D. Arends, 2003. Effects of endosulfan on brain acetylcholinesterase activity in juvenile bluegill sunfish. Environ. Res., 91: 157-162.
CrossRefDirect Link - Dutta, H.M. and L.B. Maxwell, 2003. Histological examination of sublethal effects of Diazinon on ovary of Bluegill, Lepomis macrochirus. Environ. Pollut., 121: 95-102.
CrossRef - El-Sherif, M.S., M.T. Ahmad, M.A. El-Danasoury and N.H.K. El-Nwishy, 2009. Evaluation of diazinon toxicity on nile tilapia fish (O. niloticus). J. Fish. Aquat. Sci., 4: 169-177.
CrossRefDirect Link - Fattahi, E., K. Parivar, S.G.A. Jorsaraei and A.A. Moghadamnia, 2009. The effects of diazinon on testosterone, FSH and LH levels and testicular tissue in mice. Iran J. Rep. Med., 7: 59-64.
Direct Link - Gage, M.J.G., C.P. Macfarlane, S. Yeates, R.G. Ward, J.B. Searle and G.A. Parker, 2004. Spermatozoal traits and sperm competition in Atlantic salmon: relative sperm velocity is the primary determinant of fertilization success. Curr. Biol., 14: 44-47.
CrossRef - Hamm, J.T. and D.E. Hinton, 2000. The role of development and duration of exposure to the embryotoxicity of diazinon. Aqua. Toxicol., 48: 403-418.
CrossRef - Jorsaraei, S.G.A., A. Firoozjaee, Y.Y. Pasha, E.T. Marzony and E. Sarabi, 2010. Histopathological effects of single dose treatment of diazinon on testes structure in rat. Yakhteh Med. J., 12: 39-42.
Direct Link - Kavan, L.S. S.R. Gilkolaei, G. Vossoughi, S.M.R. Fatemi, R. Safari and S. Jamili, 2009. Population genetic study of Rutilus frisii kutum (Kamansky 1901) from the Caspian Sea; Iran and Azerbaijan regions, using microsatellite markers. J. Fish. Aquat. Sci., 4: 316-322.
CrossRefDirect Link - Lasheidani, M.F., S.N. Balouchi, A. Keyvan, S. Jamili and K. Falakrou, 2008. Effects of butachlor on density, volume and number of abnormal sperms in caspian kutum (Rutilus frisii kutum, Kamenskii 1901). Res. J. Environ. Sci., 2: 474-482.
CrossRefDirect Link - Louiz, I., M. Ben-Attia and O.K. Ben-Hassine, 2009. Gonadosomatic index and gonad histopathology of Gobius niger (Gobiidea, Teleost) from Bizerta lagoon (Tunisia): Evidence of reproduction disturbance. Fish. Res., 100: 266-273.
CrossRefDirect Link - McLachlan, J.A., 2001. Environmental signaling: What embryos and evolution teach us about endocrine disrupting chemicals. Endocrine Rev., 22: 319-341.
CrossRefPubMedDirect Link - Mlambo, S.S., J.H.J. van Vuren, I.E.J. Barnhoorn and M.S. Bornman, 2009. Histopathological changes in the reproductive system (ovaries and testes) of Oreochromis mossambicus following exposure to DDT. Environ. Toxicol. Pharmacol., 28: 133-139.
CrossRef - Moore, A. and C.P. Wairing, 1996. Sublethal effects of the pesticide diazinon on olfactory function in mature male atlantic salmon parr. J. Fish Biol., 48: 758-775.
CrossRef - Pearse, A.G.E., 1985. Histochemistry: Theoretical and Applied. 4th Edn., Churchill-Livingstone, London, Pages: 614.
Direct Link - Rosengrave, P., H. Taylor, R. Montgomerie, V. Metcalf, K. McBride and N.J. Gemmell, 2009. Chemical composition of seminal and ovarian fluids of chinook salmon (Oncorhynchus tshawytscha) and their effects on sperm motility traits. Comparative Biochem. Physiol. Mol. Integrative Physiol., 152: 123-129.
CrossRefPubMed - Rurangwa, E., D.E. Kime, F. Ollevier and J.P. Nash, 2004. The measurement of sperm motility and factors affecting sperm quality in cultured fish. Aquaculture, 234: 1-28.
CrossRefDirect Link - Shayeghi, M., S.J. Shahtaheri and M. Selsele, 2001. Phosphorous insecticides residues in Mazandaran River Waters, Iran. Iran. J. Public. Health, 30: 115-118.
Direct Link - Yamaguchi, S., C. Miura, A. Ito, T. Agusa and H. Iwata et al., 2007. Effects of lead, molybdenum, rubidium, arsenic and organochlorines on spermatogenesis in fish: Monitoring at Mekong Delta area and in vitro experiment. Aquat. Toxicol., 83: 43-51.
CrossRefDirect Link - Yousefian, M. and H. Mosavi 2008. Spawning of South Caspian Kutum (Rutilus frisii kutum) in most migratory river of South Caspian Sea. Asian J. Anim. Vet. Adv., 3: 437-442.
CrossRefDirect Link - Orlu, E.E. and U.U. Gabriel, 2011. Effect of sublethal concentrations of aqueous extract of Lepidagathis alopecuroides on spermatogenesis in the fresh water catfish Clarias gariepinus. Res. J. Environ. Toxicol., 5: 27-38.
CrossRefDirect Link