
Nayer Bahavar
Department of Agronomy and Plant Breeding, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
Ali Ebadi
Department of Agronomy and Plant Breeding, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
Ahmad Tobeh
Department of Agronomy and Plant Breeding, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
Shahzad Jamaati-E-Somarin
Young Researchers Club, Department of Agronomy, Faculty of Agriculture, Islamic Azad University, Ardabil Branch, Ardabil, Iran
Research Journal of Environmental Sciences
Year: 2009 | Volume: 3 | Issue: 3 | Page No.: 332-338







ABSTRACT
This experiment was conducted to determine water use efficiency of chickpea (cv. Arman) under water deficit condition and different mineral N application. The experiment was arranged in growth chamber of Mohaghegh Ardabili University, Ardabil, Iran, in 2007, with three nitrogen levels (25, 50 and 75 kg N ha-1 as urea) and five irrigation regimes (soil water content of 25, 45, 65, 85 and 100% field capacity). The experiment was laid out in randomized complete block design (factorial) experiment with three replications. Results showed that water stress affect Water Use Efficiency (WUE), Relative Water Content (RWC), Leaf Area (LA), leaf number after stress induction, biomass (above ground tissues) and root/shoot ratio (R/S) traits. Application of Mineral nitrogen increased water use efficiency, RWC, leaf area and biomass. Maximum amount of WUE, RWC, biomass and LA obtained from 75 kg N ha-1 under water deficit condition. This experiment revealed that nitrogen fertilizer could reduce many detrimental effects of water stress. These results also suggest that using of nitrogen fertilizer in water deficit conditions (such as dry land farming areas) increase tolerance of plants to stress.
PDF Abstract XML References Citation
How to cite this article
Nayer Bahavar, Ali Ebadi, Ahmad Tobeh and Shahzad Jamaati-E-Somarin, 2009. Effects of Mineral Nitrogen on Water Use Efficiency of Chickpea (Cicer arietinum L.) under Water Deficit Condition. Research Journal of Environmental Sciences, 3: 332-338.
DOI: 10.3923/rjes.2009.332.338
URL: https://scialert.net/abstract/?doi=rjes.2009.332.338
DOI: 10.3923/rjes.2009.332.338
URL: https://scialert.net/abstract/?doi=rjes.2009.332.338
INTRODUCTION
Drought or water deficit is the most important environmental stress that limits agricultural production and reduced drought regions out-put (Soltani et al., 2001). Water stress limits leaf expansion and reduces cell turgidity and therefore, stomata closer that reduce CO2 uptake and the photosynthesis. Water stress also disrupts the balance between roots and shoot ratio (Shangguan et al., 2004). Water use efficiency also reduced by increasing root per shoot ratio. Benjamin and Nielsen (2006) reported that shoot dry weight of water stressed reduced relative to root dry weight. Water deficit decreased nitrogen fixation in bean (Verdoy et al., 2004). Water deficit reduce nitrogen fixation, because the nitrogenase is more sensitive to the water shortage. The ability of plants root to deep distribution in soil for water gain is a drought avoidance mechanism. Chickpea roots grow deeply in soil at water deficit conditions based on weather and available water and occupied bigger soil per root unit weight (Benjamin and Nielsen, 2006). Water stress reduces chlorophyll and relative water content (Chandrasekar et al., 2000; Costa-Franca et al., 2000). Water stress significantly decreased relative water content and membrane stability (Sairam and Srivastava, 2001). Saneoka et al. (2004) found that nitrogen fertilizer application at primary growth stage, reduced osmotic potential, with increasing nitrogen fertilizer to optimum level and suppose to increase the relative water content. They showed positive significant correlations between drought period and reduction of plant height, leaf area and leaf dry weight. Drought stress affects nutrients absorption which may be due to the: (i) reduction of transport from soil to the rizosphere, (ii) selective absorption of ions by roots, (iii) changes in shoot and root minerals requirement, (iv) transport reduction of elements into the plants and (v) deficiency or association of ions that cause to metabolic disorders (Alizadeh, 2002). In water deficit conditions fertilizer application increased photosynthesis and growth (Ghosh et al., 2004). Shangguan (2004) showed that the shoot dry weight and leaf gas exchange increased by increasing nitrogen availability and reduced by less nitrogen availability, whereas root growth had a negative correlation by nitrogen level. Water stress causes that leaf water potential decreased, leaves lost turgor and leaf area was reduced dramatically due to wilting in Faba bean (Mwanamwenge et al., 1999). When stress period was long, leaf water potential reduced more severely. Goicoechea et al. (1997) reported that nitrogen application increased accumulation of compatible solutes. Increase in accumulation of compatible solutes such as proline and soluble carbohydrates in water stress conditions may maintain cell turgor, RWC and LWP. Antolin et al. (1995) reported that nitrogen supplemented in an equal water potential level had higher relative water content which resulted more yield and WUE. This may result by role of nitrate in nitrogen supply, its function on osmo-regulation and increase leaf area and stomata behaviors in water stress conditions. Zhang et al. (2004) showed that water stress decreased yield, biomass and evapotranspiration. In view of these facts the present study was undertaken to evaluate. The aim this study evaluation of the effects mineral nitrogen on water use efficiency and some other traits of chickpea (cv. Arman) under water deficit condition.
MATERIALS AND METHODS
A factorial experiment (using RCB design) including three nitrogen levels (25, 50 and 75 kg N ha-1 as potassium nitrate) and five irrigation schedules (25, 45, 65, 85 and 100% of field capacity) with three replications was conducted in 2007 in a growth chamber of Mohaghegh Ardabili University, Ardabil, Iran. Seeds of Arman (a dry land chickpea cultivar in Iran) planted in 25x20 cm dimension pots. Day-night temperatures and light period were 25, 18 and 16 °C, 8 h, respectively. The soil texture was loamy with pH about 7.76. Seeds inoculated with bacteria (Mesorhizobium cicer) before planting. Mineral nitrogen was applied after water deficit induction. Pots were fertilized with 337.5 mg ammonium phosphate and 112.5 mg potassium sulfate for each pot at planting time (April, 2007). Field Capacity (FC) of soil calculated as gravimetrically. After plant establishment they thinned for three plants per pot. Irrigation regimes conducted by daily weighing of the pots to keep of expected moisture. Leaf RWC estimated from following equation (Clavel et al., 2006):
RWC = {(Fw -Dw) / (Tw -Dw)}x100 |
Leaf material was weighed (1.0 g) to determine fresh weight (Fw) and placed in double-distilled water for 4 h under dark conditions, then turgid weight (Tw) was recorded. Subsequently the samples were kept in a hot air oven at 65 °C for 48 h and then dry weight (Dw) were recorded. LA and root area were measured by leaf area meter (model SCANMAN Ci-202, 2006). Root and shoot dried at 75 °C in oven for 48 h and then weighed by 0.001 g balance. WUE calculated as follow (Alizadeh, 2002):
WUE = Dry matter (g)/Water lost (kg) |
Number of leaves that emerged and expanded after water stress induction determined the leaf number after water stress. Leaf Water Potential (LWP) was measured by means of a pressure chamber (model ELE, England). At final harvest, the three plants from each pot were harvested and oven dried at 75 °C for 48 h, then weighed by 0.001 g balance. Leaves, stems and roots were weighed separately. Data was subjected to a standard analysis of variance using SAS statistical software. Least significant differences (Duncan) tests were done at the (p<0.05) confidence level and Microsoft® Excel software used to represent the figure.
RESULTS AND DISCUSSION
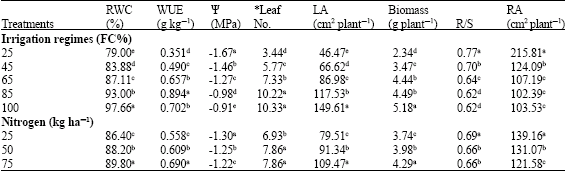
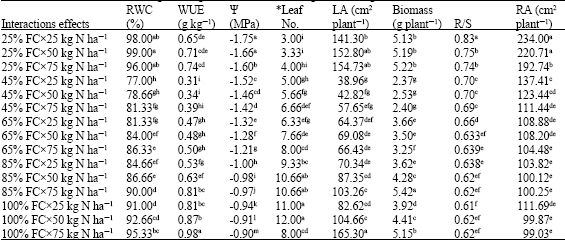
Mineral nitrogen enhanced water use efficiency (Table 2) and increasing nitrogen application to 50 and 75 kg ha-1 increased WUE by 15 and 25% (respectively) relative to 25 kg N ha-1 (Table 1). There was significantly (p<0.05) interaction between irrigation regimes and N application (Table 2). The highest WUE obtained from 85% FC and 75 kg N ha-1. By application of 25 kg N ha-1 in sever stress condition (25% FC), WUE reduced to 0.316 g kg-1, but WUE had no significant differences between 100% and 65 FC (Table 1). Reduction in soil water availability reduced water use efficiency. Although reduction of water availability from 85-65, 65-45 and 45-25% FC caused a small reduction of 5.8, 3.2 and 4.8% (respectively) in RWC; but it reduced WUE for 26.9, 24.5 and 28.5%, respectively. It shows that WUE is more susceptible to decrease relative water content because stomata opening depending on leaf water status. This reduction also can be related to the shoot growth reduction resulted from decreasing leaf area under water deficit condition.
Nitrogen application increased WUE by affecting on plant growth. By application of 25 kg ha-1 nitrogen (starter nitrogen) WUE was 0.558 g dry matter per kg used water, whereas WUE increased 25% by increasing mineral nitrogen to 75 kg ha-1 and reached to 0.690 g kg-1. Application nitrogen increased water use efficiency (Antolin et al., 1995; Athar and Johnson, 1996). Reca et al. (2001) also showed that yield reduction under water deficit condition leads to low water use efficiency.
| Table 1: | Effect of nitrogen application and water stress on measured traits of chickpea |
 | |
| Column with same letter(s) have no significant differences to each other. *Remaining leaves number after stress onset | |
| Table 2: | Interaction effects of Irrigation regimes (FC %) and Nitrogen levels (kg ha-1) on measured traits. |
 | |
| Column with same letter(s) have no significant differences to each other. *Remaining leaves number after stress onset | |
Relative Water Content (RWC) reduced significantly by water deficiency. Relative water content of 25% FC was 14.4% lower than 100% FC (Table 1), but mineral nitrogen applying improved RWC, so RWC of 75 kg N ha-1 treatment was 3.4% higher than 25 kg N ha-1. It seems that nitrogen application in water deficit condition not only supplies plant N demands, but it has an efficient roll in maintaining water status by mechanisms such as osmotic adjustment. There was significant (p<0.05) interaction between water shortage and rate of nitrogen fertilizer. The highest relative water content (98%) obtained from 100% FC with 75 kg N ha-1 (Table 2) whereas 25% FC and 25 kg N ha-1 had the least RWC (77%). Sairam and Srivasta (2001) reported that water stress significantly decreased relative water content in leaves of wheat cultivars. LRW content decreased with increased water stress (Rosales-Serna et al., 2004). Also Chandrasekar et al. (2000) observed the highest Relative Water Content (RWC) at all of the growth stages under irrigated conditions. Antolin et al. (1995) found that nitrate fed alfalfa plants in a similar water stress had higher RWC than nitrogen fixing ones.
N application (75 kg ha-1) increased leaf water potential (Table 1). There were significantly differences (p<0.01) between Leaf Water Potential (LWP) of different nitrogen amounts and nitrogen schedules, where the maximum LWP (Namely, -0.9 MPa) obtained from 75 kg N ha-1 and 100% FC. Application of 25 kg N ha-1 and 25% field capacity had the lowest (-1.75 MPa) leaf water potential ratio. Also nitrogen increased the accumulation of compatible solutes which increased leaf water potential (Antolin et al., 1995). In water stress conditions decreased leaf water potential (Grzesiak et al., 1997).
Root per shoot ratio (R/S) affected from nitrogen level and irrigation schedule (Table 1). The highest R/S ratio (0.77 and 0.69) obtained from severs stress (irrigation at 25% field capacity) and application of 25 kg N ha-1, respectively. There were no significantly differences between 85 and 100% field capacity treatments. Threshold turgor potential need for root growth is lower than shoot tissues; and thus water stress reduced shoot dry weight rather than roots. There was significantly (p<0.01) interaction between water deficiency and nitrogen fertilizer (Table 2). Application of 75 and 50 kg N ha-1 had the lowest (0.66) R/S ratio. The shoot dry weight increased with the increase of nitrogen supply, but decreased when nitrogen supply reached a certain level. The root growth was negatively correlated with the increase of nitrogen supply. The water use efficiency decreased with increasing ratio of R/S, while leaf photosynthetic rate tended to increase initially and then decrease. The increase in root/shoot was unfavorable to increase water use efficiency (Shangguan et al., 2004). Kafi et al. (2005) reported that the ratio of root to shoot (R/S) is an index that reflects growth and dry matter accumulation between the root and shoot. These results showed that water stress affect shoot growth rather than root, i.e., growth of above ground tissues are more susceptible to water deficits. Shoot dry weight decreased by increasing water shortage but nitrogen supply ameliorate stress effects on shoot growth (Shangguan et al., 2004).
It was observed that Root Area (RA) also affected from water deficit and hence increasing soil water from 25% FC to 45, 65, 85 and 100% decreased root area (Table 1). There were significant differences (p<0.05) between Root Area (RA) of different nitrogen amounts and irrigation schedules, where the highest RA (234 cm2 plant-1) obtained from 25 kg N ha-1 and 25% FC. Application of 75 kg N ha-1 had the lowest (99.03 cm2 plant-1) root area ratio. There were no significantly differences between root area of 100, 85 and 65% field capacity (Table 2). Benjamin and Nielsen (2006) also reported that water deficit stress resulted in a greater proportion of chickpea roots to grow deeper in the soil. Also in water stress conditions, chickpea occupy large surface area per unit root weight. In other study showed that the greatest root growth in an average year followed by lesser root growth in the dry year and the least root growth in the wettest year (Merril et al., 2002). The plant root growth decreased with the increase nitrogen application was reported by Shangguan et al. (2004).
There were significantly differences (p<0.01) between biomass of different nitrogen amounts and irrigation schedules (Table 1). The maximum biomass (5.226 g plant-1) obtained from 100% FC with 75 kg N ha-1 and the lowest (2.371 g plant-1) belongs to sever stress (25% FC) with minimum nitrogen (25 kg N ha-1) application (Table 2). Water stress affected biomass and seed yield more severely and secondary branches were more susceptible than primary ones (Leport et al., 2006). Increasing water availability (25-45% FC) increased biomass up to 48%, but biomass rising up to 45% FC was lower (27%) than (25-45% FC). Athar and Johnson (1996) also concluded that reducing soil water potential form -0.3 to -1 MPa decreased total dry matter by 65%. In a greenhouse study with one cultivar of chickpea both the creation and abortion of pods reduced by water deficit imposed after the commencement of pod set (Behboudian et al., 2001). There were significantly differences between mineral nitrogen and biomass production and the highest biomass (4.29 g plant-1) obtained from 75 kg ha-1 (Table 1). Unless biomass of 25 and 100% FC there was significant differences by nitrogen amounts between 85-45% FC, where consumption of 25 kg N ha-1 at 85% FC was equal to biomass of 75 kg N at 45% FC (Table 2). These results showed that nitrogen deficiency in water restricted condition is the most limiting factor of biomass production.
Leaf Area (LA) also affected from water deficit (Table 1). In this study increasing soil water from 25-45, 45-65 and 65-85%, increased leaf area 30, 23.6 and 26, respectively. Reduction of leaf area on insufficient soil water results from both little leaf expansion and enhancing older leaf senescence. Reducing soil water from 45-25% decreased leaf number from 5.77-3.44 leaves per plant. Decreasing leaf area and leaf number reduced dry matter production by lowering carbon assimilation. Nitrogen application caused to increase the leaf area. There was significantly (p<0.01) interaction between water deficiency and nitrogen fertilizer, where the highest LA (155.3 cm2 plant-1) obtained from 75 kg N ha-1 and 85% FC. The lowest leaf area (38.96 cm2 plant-1) obtained from sever stress (25% field capacity) with 25 kg N ha-1 (Table 2). This experiment showed that leaf area growth is more sensitive to the water deficit. Pagter et al. (2005) reported that water deficit decreased leaf number and leaf area. In water stress conditions increased nitrogen fertilizer causes increased leaf area (Lindmann and Glover, 2003).
Leaf number affected from soil water deficiency and mineral nitrogen (Table 1). There was significantly (p<0.01) interaction between water shortage and nitrogen application in response to the leaf number. After stress imposing the maximum leaf number (12) obtained from 85 and 100% field capacity, whereas it was three fold greater than the leaf number of 25% FC (Table 2). There were no significantly differences between leaf number of 100 and 85% FC. Nitrogen application up to 50 kg N ha-1 increased leaf number, but effects of 50 and 75 kg N was similar (Table 2). Oweis et al. (2004) observed that water deficit decreases green leaf area, photosynthesis rate, stomatal size, dry matter production and seed yield of Faba bean. Mwanamwenge et al. (1999) also reported that water deficit decreased rates of leaf expansion, specific leaf area, leaf shedding, shoot dry matter production but increased root dry matter of faba bean. In water stress conditions, growth and yield increased by nitrogen application (Yang et al., 2000).
CONCLUSION
Water stress affects on some growth and physiological aspects of plants. Reduction of nutrient availability in root zone is a growth limiting factor resulted from water deficits. In legumes that their N demand only supplied with N fixing mechanism, suffer more damage from water stress because this phenomenon (N fixing) is more susceptible to water deficit. Under these conditions, not only plant N demand increase but also some surviving phenomenon (such as osmotic adjustment via proline and other amino acid accumulation) depending on nitrogen supplemented. Results of this study showed that increasing mineral nitrogen from 25 (starter N) to 50 and 75 kg ha-1 increased RWC about 3.4%, leaf area 29.96%, LWP 0.08%. Such positive effects of mineral nitrogen application caused water use efficiency increased for 25%. Mineral N suppressed photoassimilate partitioning to roots and increased it to above ground organs. By such mechanisms water use efficiency increased. These results also suggest that using of nitrogen fertilizer in water deficit conditions (such as dry land farming areas) increase tolerance of plants to stress.
ACKNOWLEDGMENTS
This study was supported by the Central Laboratory of Agricultural Faculty, University of Mohaghegh Ardabili. Valuable experimental support by Aziz Jamaati-E-Somarin and Rogayyeh Zabihi-E-Mahmoodabad is greatly appreciated.
REFERENCES
- Antolin, M.C., J. Yoller and M. Sachez-Diaz, 1995. Effects of temporary drought on nitrate-fed and nitrogen-fixation alfalfa plants. Plants Sci., 107: 159-165.
CrossRef - Athar, M. and D.A. Johnson, 1996. Nodulation biomass production and nitrogen fixation in alfalfa under drought. J. Plant Nutr., 19: 185-199.
Direct Link - Behboudian, M.H., Q. Ma, N.C. Turner and J.A. Palta, 2001. Reactions of chickpea to water stress: Yield and seed composition. J. Sci. Food Agric., 81: 1228-1291.
CrossRefDirect Link - Benjamin, J. and D. Nielsen, 2006. Water deficit effects on root distribution of soybean, field pea and chickpea. J. Field Crops Res., 97: 248-253.
Direct Link - Clavel, D., O. Diouf, J.L. Khalfaoui and S. Braconnier, 2006. Genotypes variations in fluorescence parameters among closely related groundnut (Arachis hypogaea L.) lines and their potential for drought screening programs. J. Field Cops Res., 96: 296-306.
CrossRef - Ghosh, P.K., K.K. Ajay, M.C. Bandyopadhyay, K.G. Manna and A.K. Mandal et al., 2004. Comparative effectiveness of cattle manure, poultry manure, phosphocompost and fertilizer-NPK on three cropping system in vertisols of semi-arid tropics. Dry matter yield, nodulation. Chlorophyll content and enzyme activity. Bioresour. Technol., 95: 85-93.
CrossRef - Goicoechea, N., M.C. Antolin and M. Sanchez-Diaz, 1997. Influence of arbuscular mycorrhizae and rhizobium on nutrient content and water relations in drought stressed alfalfa. J. Plant Soil, 192: 261-268.
CrossRefDirect Link - Grzesiak, S., M. Iijima, Y. Kono and A. Yamauchi, 1997. Differences in drought tolerance between cultivars of field bean and field pea. A comparison of drought- resistant and drought-sensitive cultivars. Acta. Physiol. Plantarum., 3: 349-357.
CrossRefDirect Link - Hubick, K.T., 1990. Effects of amount and source of nitrogen and water limitation on growth, transpiration efficiency and carbon-isotope discrimination in peanut cultivars. Aust. J. Plant Physiol., 17: 413-430.
Direct Link - Leport, L., N.C. Turner, S.L. Davies and K.H.M. Siddique, 2006. Variation in pod production and abortion among chickpea cultivars under terminal drought. Eur. J. Agron., 24: 236-246.
CrossRef - Merril, S.D., D.L. Tanaka and J.D. Hanson, 2002. Root length growth of eight crop species in Haplustoll soils. Soil Sci. Soc. Am. J., 66: 913-923.
Direct Link - Mwanamwenge, J., S.P. Loss, K.H.M. Siddique and P.S. Cocks, 1999. Effect of water stress during floral initiation, flowering and podding on growth and yield of faba bean (Vicia faba L.). Eur. J. Agron., 11: 1-11.
Direct Link - Oweis, T., A. Hachum and M. Pala, 2004. Lentil production under supplemental irrigation in a Mediterranean environment. Agric. Water Manage., 68: 251-265.
CrossRef - Pagter, M., C. Bragato and H. Brix, 2005. Tolerance and physiological responses of phragmites Australis to water deficit. Aquat. Bot., 81: 285-299.
CrossRefDirect Link - Reca, J., J. Roldan, M. Alcaide, R. Lopez and E. Camacho, 2001. Optimization model for water allocation in deficit irrigation systems. I. Description of the model. Agric. Water Manage., 48: 103-116.
CrossRefDirect Link - Rosales-Sernaa, R., J. Kohashi-Shibataa, J.A. Acosta-Gallegosb, C. Trejo-Lo Peza and J. Ortiz-Cereceresc et al., 2004. Biomass distribution, maturity acceleration and yield in drought-stressed common bean cultivars. Field Crops Res., 85: 203-211.
CrossRef - Sairam, R.K. and G.C. Srivastava, 2001. Water stress tolerance of wheat (Triticum aestivum L.): Variations in heydrogen peroxide accumulation and antioxidant activity in tolerant and susceptible genotypes. J. Agron. Crop Sci., 186: 63-70.
Direct Link - Saneoka, H., R.E.A. Moghaieb, G.S. Premachandra and K. Fujita, 2004. Nitrogen nutrition and water stress effects on cell membrane stability and leaf water relation in Agrostis palustris hunds. Environ. Exp. Bot., 52: 131-138.
CrossRef - Shangguan, Z.P., M.A. Shao, S.J. Ren, L.M. Zhang and Q. Xue, 2004. Effect of nitrogen on root and shoot relations and gas exchange in winter wheat. Bot. Bull. Acad. Sin., 45: 49-54.
Direct Link - Soltani, A., F.R. Khooie, K. Ghassemi-Golezani and M. Moghaddam, 2001. A simulation study of chickpea crop response to limited irrigation in a semiarid environment. Agric. Water Manage., 49: 225-237.
CrossRefDirect Link - Verdoy, D., M.M. Lucas, E. Manrique, A.A. Covarrubias and M.R. Defelipe et al., 2004. Differential organ-specific response to salt stress and water deficit in nodulated bean (Phaseolus vulgaris). Plant Cell Environ., 27: 757-767.
Direct Link - Yang, J., J. Zahang, Z. Huang, Q. Zhu and L. Liu, 2001. Water deficit induced senescence and its relationship to the remobilization of ore stored carbon in wheat during grain filling. Agron. J., 93: 196-206.
Direct Link - Yang, J., J. Zahang, Z. Huang, Q. Zhu and L. Wang, 2000. Remobilization of carbon reserves is improved by controlled soil drying during grain filling of wheat. Crop Sci., 40: 1645-1655.
Direct Link - Zhang, Y., E. Kendy, Y. Qiang, L. Changming, S. Yanjun and S. Hongyong, 2004. Effect of soil water deficit on evapotranspiration, crop yield and water use efficiency in the North China Plain. Agric. Water Manage., 64: 107-122.
CrossRefDirect Link - Costa-Franca, M.G., A.T. Pham Thi, C. Pimentel, R.O. Pereyra Rossiello, Y. Zuily-Fodil and D. Laffray, 2000. Differences in growth and water relations among Phaseolus vulgaris cultivars in response to induced drought stress. Environ. Exp. Bot., 43: 227-237.
CrossRefDirect Link - Chandrasekar, V., R.K. Sairam and G.C. Srivastava, 2000. Physiological and biochemical responses of hexaploid and tetraploid wheat to drought stress. J. Agron. Crop Sci., 185: 219-227.
CrossRefDirect Link