
Louis K. Prom
USDA-ARS, Plains Area Agricultural Research Center, College Station, 77845, Texas
LiveDNA: 1.3546
Hugo Cuevas
USDA-ARS, Tropical Agriculture Research Station, 2200 Pedro Albizu Campos Avenue, Mayaguez, 00680, Puerto Rico
Ramasamy Perumal
Kansas State University, Agricultural Research Center, Hays, 67601, Kansas
LiveDNA: 91.1156
Thomas Isakeit
Department of Plant Pathology and Microbiology, Texas A and M University, College Station, 77843, Texas
Clint Magill
Department of Plant Pathology and Microbiology, Texas A and M University, College Station, 77843, Texas
Plant Pathology Journal
Year: 2018 | Volume: 17 | Issue: 2 | Page No.: 75-79







ABSTRACT
Background and Objective: Anthracnose incited by Colletotrichum sublineola is the most important foliar disease of sorghum worldwide. The hyper-variable nature of the pathogen requires continual evaluations of sorghum germplasm to identify sources with different resistance genes. Thus, this research was undertaken to determine the inheritance of two Sudanese lines PI570726 and PI569979 to pathotype 30 and BT×378 (Redlan) to pathotypes 30 (P30) and 35 (P35) of the anthracnose pathogen. Materials and Methods: Crosses between these three sorghum lines and a susceptible parent PI609251 were performed and the resulting F2 populations were evaluated in greenhouse. Seeds were planted in 1-gallon cans and 30 d post planting, plants were inoculated with the anthracnose pathotypes. Individual plants from each cross were scored based on a scale of 1-5, where 1 = No symptoms or chlorotic flecks on leaves; 2 = Hypersensitive reaction; 3, 4 and 5= Infected leaf lesions with acervuli formation. Results: Chi-square test for the F2 populations of BT×378×PI609251 against P30 and P35, segregated in the expected Mendelian ratio 3 (resistant): 1(susceptible) fashion, indicating that resistance in BT×378 to P30 and P35 of C. sublineola obtained from the USA is governed by a single dominant gene. Segregation analysis for the PI569979×PI609251 and PI570726×PI609251 F2 populations revealed 1:3 ratio of resistance to susceptible, indicating that resistance in PI569979 and PI570726 to P30 was conditioned by a recessive trait. Conclusion: Resistance in the sorghum line BT×378 was shown to be dominant and will therefore be more desirable for use in breeding for anthracnose resistance, especially in sorghum hybrid production.
PDF Abstract XML References Citation
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Louis K. Prom, Hugo Cuevas, Ramasamy Perumal, Thomas Isakeit and Clint Magill, 2018. Inheritance of Resistance of Three Sorghum Lines to Pathotypes of Colletotrichum sublineola, Causal Agent of Anthracnose. Plant Pathology Journal, 17: 75-79.
DOI: 10.3923/ppj.2018.75.79
URL: https://scialert.net/abstract/?doi=ppj.2018.75.79
DOI: 10.3923/ppj.2018.75.79
URL: https://scialert.net/abstract/?doi=ppj.2018.75.79
INTRODUCTION
Sorghum is one of the most indispensable crops, especially in the drier tropics. However, the crop is hampered by a number of biotic stresses, including anthracnose incited by the fungal pathogen Colletotrichum sublineola Henn1,2. In some regions, yield losses up to 50% may occur when susceptible lines are planted3-5. The pathogen infects the leaves, panicle and stalk, reducing both the yield and grain quality2. Colletotrichum sublineola reproduces asexually by producing acervuli, a fruiting body with cushion like structure containing enormous amounts of conidia. Acervuli detection in sorghum foliar is often used as a diagnostic sign for anthracnose susceptibility2,6,7.
Large number of resistant sorghum lines to anthracnose had been identified; however, the hyper-variable nature of the pathogen requires continuous evaluation to identify new sources of resistance with different gene action to the disease5,8-17. Although the sorghum gene pool is diverse, knowledge still is lacking on the distribution, inheritance and effectiveness of resistance genes in diverse environments. Effective control of sorghum diseases by breeding resistant lines will be enhanced by a more thorough understanding of the distribution and nature of both resistance genes and pathotype diversity. Over the years, studies on the inheritance of anthracnose resistance in some sorghum lines were conducted in different sorghum growing regions18-22.
However, there is limited or no knowledge on the inheritance and allelic relationships of host plant resistance and the sorghum differentials to diverse pathotypes of the anthracnose pathogen. A set of 18 sorghum differentials was compiled by Prom et al.5 and these three lines were taken from it. Currently, this set or some of the lines within the set of differentials are being used in several countries to establish C. sublineola pathotypes.
Thus, this study was conducted to determine the inheritance of anthracnose resistance in three lines [PI569979, PI579726 and BT×378 (Redlan)] selected from the set of sorghum differentials for pathotype differentiation.
MATERIALS AND METHODS
Plant materials: The two Sudanese lines PI569979 and PI579726 and BT×378 were crossed to a susceptible parent (PI609251) to obtain the F2 populations.
Pathotype: Two pathotypes (P30 and P35) of C. sublineola were selected for this study. The pathotypes were based on the response of 18 sorghum differentials compiled by Prom et al.5. The 18 differentials included PI570726 which was resistant to P30 and PI569979 which susceptible to this pathotype. BT×378 was resistant to both P30 and P35 while PI570726 and PI569979 were susceptible to P35. Pathotype 30 was obtained from Texas and P35 was from Georgia.
Greenhouse evaluation: Seeds of the parental lines and F2 from the crosses with a susceptible parent PI609251 were planted at a rate of 4 seeds per 1-gallon cans (Hummert International) containing metro mix 200 (BWI) amended with osmocote classic fertilizer 17-7-12 (O.M. Scott and Sons Company). On a bi-weekly basis, plants were fertilized with Peters excel multi-purpose 15-5-15 (O.M. Scott and Sons Company).
Inoculum and inoculation protocol: The inoculum preparation, inoculation technique and disease assessment method were as described by Prom et al.7. Two inoculation methods were utilized in this study. In the colonized grain method, 10 C. sublineola-colonized sorghum seeds were dropped into the leaf whorl of each plant at 30 days after planting. In the spore suspension method, each C. sublineola pathotype was grown separately in Petri plates containing half-strength potato dextrose agar medium and incubated at 25°C for 10 days under a 12 h photoperiod provided by fluorescent (F40CW/RS/EW-11) and blacklight (emits visible light and UV light). Conidia for the different pathotypes were harvested by flooding the plates with 10 mL sterilized water and scraping the agar surface to dislodge them. The conidial suspensions were filtered through two layers of cheesecloth into separate Erlenmeyer flasks and diluted with sterile water to obtain final concentrations of 1×106conidia mL–1. In each flask, 0.5 mL Tween 80 added per liter. Plants were spray inoculated with approx. About 3-5 mL of conidial suspension and allowed to dry for 30 min. Post-inoculation, plants were misted daily for 30 sec at 45 min intervals for 8 h–1 for 20 days to provide ideal conditions for infection and disease development.
Disease assessment method: Disease assessment was conducted 21 day-post-inoculation using the 1-5 scale as described by Prom et al.7, where 1: No symptoms or chlorotic flecks on leaves, 2: Hypersensitive reaction (reddening or red spots) on inoculated leaves but no acervuli formation and no spreading to other leaves, 3: Lesions on inoculated and bottom leaves with acervuli in the center, 4: Necrotic lesions with acervuli on the bottom and middle leaves and 5: Most leaves dead due to infection with infection on the flag leaf containing abundant acervuli.
 | |
| Fig. 1: | Anthracnose infected leaves: in the susceptible response there is the presence of acervuli (dark fruiting bodies) on the infected leaves, which indicates successful reproduction of the pathogen. Under favorable environmental conditions, masses of spores are produced and splash unto other leaves |
| Table 1: | Chi-square analysis for the crosses evaluated against Colletotrichum sublineola pathotypes 30 |
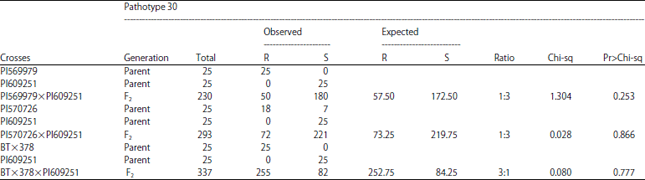 | |
Total: Total number of plants, R: Resistant response, S: Susceptible response | |
Symptom types were then categorized into two reaction classes, resistant rated as 1 or 2 and susceptible rated as 3, 4 or 5. Figure 1 shows a susceptible response.
Statistical analysis: Chi-square test (SAS version 9.4, SAS Institute, Cary, NC) was performed to determine the goodness-of-fit [Mendelian 3 (resistant):1 (susceptible) or 1(resistant):3 (susceptible) ratio] for the number of gene(s) segregating in three F2 populations derived from crosses of BT×378, PI569979 and PI579726 with a susceptible parent PI609251.
RESULTS
In this study, the inheritance of anthracnose resistance in three sorghum lines PI569979, PI579726 and BT×378 (Redlan) against pathotypes, P30 and P35 of C. sublineola was determined. The results of the chi-square test for the F2 populations of BT×378×PI609251 against P30 and P35, segregated in the expected Mendelian ratio 3 (resistant): 1 (susceptible) fashion, indicating that resistance in BT×378 to the two pathotypes of C. sublineola obtained from the USA was governed by a single dominant gene (Table 1, 2). Segregation analysis for the PI569979×PI609251 F2 population revealed 1:3 ratio of resistance to susceptible, indicating that resistance in PI569979 to P30 was conditioned by a single recessive gene (Table 1).
| Table 2: | Chi-square analysis for the crosses evaluated against Colletotrichum sublineola pathotypes 35 |
 | |
| Total: Total number of plants, R: Resistant response, S: Susceptible response | |
PI570726 was shown to be susceptible to P30 by Prom et al.5. When the segregating F2 population derived from PI570726×PI609251 was challenged with P30, the chi-square test also revealed a Mendelian 1:3 ratio of resistance to susceptible, suggesting a recessive trait (Table 1).
DISCUSSION
The durability and effectiveness of controlling sorghum diseases by breeding resistant lines will require a more exhaustive knowledge of the distribution and nature of resistance genes. However, the hyper-variable nature of C. sublineola makes the understanding of the inheritance of anthracnose resistance more challenging5,8-12,16,19. Resistance for BT×378 against P30 and P35 was shown to be governed by a single dominant gene. Similarly, Erpelding20 also noted that anthracnose resistance in BT×378 was governed by a single dominant gene when F2 populations derived from BT×378×PI609251 were evaluated using Puerto Rico isolates in Isabela, Puerto Rico. Over the years, studies on the inheritance of anthracnose resistance in other sorghum lines have been reported18-22. Boora et al.18 noted that resistance in SC326-6 to anthracnose when crossed with the susceptible line BT×623 segregated as a simple recessive trait, while Da Costa et al.21 found that resistance in SC283 was also controlled by a recessive gene. Resistance in SC137, SC166, SC991, SC120 and SC748-5 were reported by Mehta et al.19 to be controlled by a single dominant gene. Also, segregating F2 populations of SC112-14×PI609251 evaluated against a number of C. sublineola pathotypes indicated that resistance in SC112-14 is governed by a single dominant gene22. Thakur and Mathur2 noted that anthracnose resistance is also influenced by other genes based on the fact that resistant F1 sorghum hybrids are often found in progenies of crosses between two susceptible parents. A dominant gene (Cg1), derived from cross between SC748 and BT×623, was mapped at the distal region of chromosome five in sorghum23. Cuevas et al.22 also mapped the resistance in SC112-14 in chromosome 5 but in different locations to the Cg1.
CONCLUSION
BT×378 could be a candidate in breeding for anthracnose resistance. However, additional evaluation with pathotypes from other sorghum growing regions is needed before mapping the resistance gene in this line.
SIGNIFICANCE STATEMENTS
This study has shown that the anthracnose resistance in sorghum line BT×378 was dominant, whereas resistance in PI569979 and PI570726 was recessive. Therefore BT×378 will be more desirable for use in breeding for anthracnose resistance, especially in sorghum hybrid production because at least one of the parents will exhibit a dominant allele. In addition to identifying a sorghum line BT×378 that could be used in breeding programs, the three lines are part of a set of sorghum differentials to delimit the C. sublineola pathotypes. Understanding the inheritance of resistance for these lines and other lines within the set of differentials, we will be able to determine whether the interaction between the pathogen with sorghum follows a gene-for-gene pattern.
ACKNOWLEDGMENT
The research was supported by the Department of Agriculture, Agricultural Research Service, United States, Project No. 3091-22000-034-00-D. We thank Roxanne Jacobsen for her technical assistance.
REFERENCES
- Prom, L.K., J. Erpelding, R. Perumal, T. Isakeit and H. Cuevas, 2012. Response of sorghum accessions from four African countries against Colletotrichum sublineolum, causal agent of sorghum anthracnose. Am. J. Plant Sci., 3: 125-129.
CrossRefDirect Link - Ngugi, H.K., S.B. King, G.O. Abayo and Y.V.R. Reddy, 2002. Prevalence, incidence and severity of sorghum diseases in Western Kenya. Plant Dis., 86: 65-70.
CrossRefDirect Link - Prom, L.K., R. Perumal, S.R. Erattaimuthu, C.R. Little and E.G. No et al., 2012. Genetic diversity and pathotype determination of Colletotrichum sublineolum isolates causing anthracnose in sorghum. Eur. J. Plant Pathol., 133: 671-685.
CrossRefDirect Link - Prom, L.K., J.E. Erpelding and N. Montes-Garcia, 2007. Chinese sorghum germplasm evaluated for resistance to downy mildew and anthracnose. Commun. Biometry Crop Sci., 2: 26-31.
Direct Link - Prom, L.K., R. Perumal, J.E. Erpelding, T. Isakeit, N. Montes-Garcia and C.W. Magill, 2009. A pictorial technique for mass screening of sorghum germplasm for anthracnose (Colletotrichum sublineolum) resistance. Open Agric. J., 3: 20-25.
Direct Link - Ali, M.E.K. and H.L. Warren, 1987. Physiological races of Colletotrichum graminicola on sorghum. Plant Dis., 71: 402-404.
Direct Link - Cardwell, K.F., P.R. Hepperly and R.A. Frederiksen, 1989. Pathotypes of Colletotrichum graminicola and seed transmission of sorghum anthracnose. Plant Dis., 73: 255-257.
CrossRefDirect Link - Pande, S., L.K. Mughogho, R. Bandyopadhyay and R.I. Karunakar, 1991. Variation in pathogenicity and cultural characteristics of sorghum isolates of Colletotrichum graminicola in India. Plant Dis., 75: 778-783.
Direct Link - Casela, C.R., A.S. Ferreira and R.E. Schaffert, 1992. Physiological Races of Colletotrichum graminicola in Brazil. In: Sorghum and Millets Diseases: A Second World Review, De Milliano, W.A.J., R.A. Frederiksen and G.D. Bengston (Eds.). International Crops Research Institute for the Semi-Arid Tropics, India, pp: 209-212.
- Marley, P.S., R.P. Thakur and O. Ajayi, 2001. Variation among foliar isolates of Colletotrichum sublineolum of sorghum in Nigeria. Field Crops Res., 69: 133-142.
CrossRefDirect Link - Erpelding, J.E. and L.K. Prom, 2004. Evaluation of malian sorghum germplasm for resistance against anthracnose. Plant Pathol. J., 3: 65-71.
CrossRefDirect Link - Da Costa, R.V., C.R. Casela, L. Zambolim, F.G. Santos and F.X. do Vale, 2005. Evaluation of genetic mixtures of sorghum lines for anthracnose resistance management. Fitopatol. Bras., 30: 525-526.
CrossRefDirect Link - Thakur, R.P., V.P. Rao, B.M. Wu, K.V. Subbarao and K. Mathur et al., 2007. Genetic resistance to foliar anthracnose in sorghum and pathogenic variability in Colletotrichum graminicola. Indian Phytopathol., 60: 13-23.
Direct Link - Moore, J.W., M. Ditmore and D.O. TeBeest, 2008. Pathotypes of Colletotrichum sublineolum in Arkansas. Plant Dis., 92: 1415-1420.
CrossRefDirect Link - Prom, L.K., R. Perumal, H. Cuevas, G. Radwan, S. Katile, T. Isakeit and C. Magill, 2016. Assessing the vulnerability of sorghum converted lines to anthracnose and downy mildew infection. J. Agric. Crops, 2: 101-106.
Direct Link - Boora, K.S., R. Frederiksen and C. Magill, 1998. DNA-based markers for a recessive gene conferring anthracnose resistance in sorghum. Crop Sci., 38: 1708-1709.
Direct Link - Mehta, P.J., C.C. Wiltse, W.L. Rooney, S.D. Collins and R.A. Frederiksen et al., 2005. Classification and inheritance of genetic resistance to anthracnose in sorghum. Field Crops Res., 93: 1-9.
CrossRefDirect Link - Erpelding, J.E., 2007. Inheritance of anthracnose resistance for the sorghum cultivar redlan. Plant Pathol. J., 6: 187-190.
CrossRefDirect Link - Da Costa, R.V., L. Zambolim, L.V. Cota, D.D. da Silva, J.A.S. Rodrigues, F.D. Tardin and C.R. Casela, 2011. Genetic control of sorghum resistance to leaf anthracnose. Plant Pathol., 60: 1162-1168.
CrossRefDirect Link - Cuevas, H.E., L.K. Prom and J.E. Erpelding, 2014. Inheritance and molecular mapping of anthracnose resistance genes present in sorghum line SC112-14. Mol. Breed., 34: 1943-1953.
CrossRefDirect Link - Ramasamy, P., M.A. Menz, P.J. Mehta, S. Katile and L.A. Gutierrez-Rojas et al., 2009. Molecular mapping of Cg1, a gene for resistance to anthracnose (Colletotrichum sublineolum) in sorghum. Euphytica, 165: 597-606.
CrossRefDirect Link