
John E. Erpelding
USDA-Agricultural Research Service, Tropical Agriculture Research Station,
2200 Pedro Albizu Campos Ave., Suite 201, Mayaguez, Puerto Rico 00680-5470, USA
Louis K. Prom
USDA-Agricultural Research Service, Southern Plains Agriculture Research Center,
2765 F and B Road, College Station, Texas 77845, USA
Plant Pathology Journal
Year: 2004 | Volume: 3 | Issue: 2 | Page No.: 65-71







ABSTRACT
Host plant resistance provides an economical approach to stabilize crop production and enhance profitability. Highly variable pathogens, such as anthracnose, will require additional sources of host resistance for crop improvement. Sorghum germplasm from Mali, West Africa was artificially inoculated with anthracnose (Colletotrichum sublineolum) to identify sources of resistance to the disease. During the dry and wet growing seasons in 2003, 270 sorghum accessions from the Mali working collection were evaluated for anthracnose resistance in Puerto Rico. A resistant response was observed for 245 accessions during the dry season and for 215 accessions in the wet season. The results would indicate that climatic conditions were more favorable for disease development during the wet season. The resistance response for 196 accessions was also effective against Texas isolates of the pathogen. The mode of inheritance for 41 accessions indicated the presence of dominant and recessive gene action. Genetic variation for resistance and resistance to multiple pathotypes of the disease would suggest that Mali sorghum germplasm would be useful for sorghum improvement.
PDF Abstract XML References Citation
How to cite this article
John E. Erpelding and Louis K. Prom, 2004. Evaluation of Malian Sorghum Germplasm for Resistance Against Anthracnose. Plant Pathology Journal, 3: 65-71.
DOI: 10.3923/ppj.2004.65.71
URL: https://scialert.net/abstract/?doi=ppj.2004.65.71
DOI: 10.3923/ppj.2004.65.71
URL: https://scialert.net/abstract/?doi=ppj.2004.65.71
INTRODUCTION
Colletotrichum sublineolum P. Henn., Kabát and Bubák is the causal organism of anthracnose specific to Sorghum species[1-3]. The pathogen is capable of infecting all above ground tissues of the sorghum plant; however, the foliar disease is most widely distributed. Foliar infection can occur at any stage of plant development, but symptoms are generally observed 40 days after seedling emergence. Characteristic symptoms on susceptible cultivars include small circular to elliptical spots or elongated lesions and as the fungus sporulates, fruiting bodies (acervuli) appear as black spots in the center of the lesions[3]. Coalescence of lesions can occur on susceptible cultivars eventually covering most of the leaf and resulting in senescence. Under conditions of high disease severity, premature defoliation can occur, which can retard plant growth and development or result in plant death prior to maturity and no seed development. Infection can also occur in the leaf midrib when little or no leaf damage is present and may be an independent response from leaf infection[3,4]. Foliar disease symptomology can be highly variable depending on the plant host, pathogen and weather conditions[4].
The stalk, panicle and grain can also be infected with anthracnose. Stalk infection, referred to as red stalk rot, can result in lodging in susceptible cultivars and may be independent from foliar disease development[5,6]. Anthracnose infection of the panicle and grain may also be independent from foliar and stalk disease development[3]. Anthracnose infection in the panicle results in death of some or all florets within the panicle with lesions and acervuli formation present on the glumes and panicle branches. In addition, infection of the peduncle can contribute to panicle breakage prior to harvest due to rotting of the interior of the stalk. Anthracnose infection on the grain appears as black streaking due to the formation of acervuli. Panicle and grain infection can result in sterility, grain abortion, premature grain ripening, reduced seed size and poor head development[3,7,8]. Infected panicles and grain can also contribute to the spread of the disease or the introduction of novel pathotypes to new regions[9].
Anthracnose is presently found in many sorghum growing regions of the world and was first identified in Togo, West Africa in 1902[3,10]. In the United States, the disease was first observed in Texas in 1912[7]. Anthracnose stalk rot was not reported until 1943. Anthracnose is considered one of the most destructive diseases of sorghum where warm, humid climatic conditions are prevalent[3,9]. Anthracnose was a major factor limiting the production of sorghum in the Southeastern United States[7]. The disease is also a major constraint to sorghum production in West and Central Africa and is widely distributed in the Sahel, Sudan and Guinea zones of West Africa, which includes the sorghum producing regions of Mali[4,8,11,12]. During severe epiphytotics, grain yield losses of 50% or greater have occurred for susceptible cultivars from the foliar phase of the disease[3,7,13]. Under experimental conditions in Nigeria, yield losses as high as 47% have been reported[4]. Grain yield losses of 41 to 67% have also been reported in experimental evaluations of susceptible cultivars infected with anthracnose in Mali and leaf death was observed prior to physiological maturity[8]. A reduction in grain weight of 18 to 36% was also observed due to anthracnose infection of susceptible cultivars in Mali. In comparison, no yield losses were reported in Mali for cultivars with moderate resistance to sorghum anthracnose. Grain yield losses due to panicle infection of susceptible cultivars have ranged from 30-50% during severe epidemics[3]. Grain yield losses are generally associated with a reduction in kernel number and size[3,8].
Although sorghum anthracnose can be controlled through the use of fungicides and cultural practices, the use of resistant cultivars provides a more economical and environmentally safe alternative. Variation in the pathogen has been a major obstacle in the development and utilization of resistant cultivars. Numerous studies have indicated that the pathogen is highly variable for cultural and spore morphology, sporulation and growth rate, symptom type, pathogenicity, virulence and aggressiveness[4,10,14]. Molecular genetic analyses have also demonstrated variability at the DNA level for the pathogen[15,16]. In addition, pathotype variation is extensive within and between regions for the disease[9,10,17,18]. In contrast, limited information is available on plant host resistance. Most published studies have evaluated a limited number of sorghum cultivars, which were selected based on known disease response. Information on the mode of inheritance of host resistance is also lacking. Thakur and Mathur[3] indicated that known sources of resistance to anthracnose harbor multiple resistance genes and that tissue specific infection may be controlled by different genes. The mode of inheritance of anthracnose resistance has been determined for only a limited number of cultivars[5,9,20] and no information is available on allelic variation.
The development of anthracnose resistant cultivars will require the identification of additional sources of resistance. Therefore, a study was conducted to evaluate sorghum genetic resources from Mali that are maintained by the US National Plant Germplasm System. The Mali sorghum working collection was selected for evaluation in order to sample a genetically diverse collection of germplasm from a region where anthracnose is considered problematic due to favorable climatic conditions. Disease pressure may contribute to variation for resistance in the host providing an opportunity to identify novel sources of resistance that could be incorporated into sorghum improvement programs.
MATERIALS AND METHODS
The US National Plant Germplasm System maintains 2,343 sorghum accessions collected from Mali, West Africa. In 2000 and 2001, quarantine seed increases were conducted at the USDA-ARS Germplasm Introduction and Research Unit in St. Croix, US Virgin Islands for 1,774 sorghum accessions from Mali. The Mali working collection was established from this increase with 273 accessions selected. Sorghum accessions were selected based on phenotypic diversity, agronomic performance and breeding potential. The selection process involved visual comparisons between accessions by several sorghum researchers to identify accessions for the working collection representing approximately 10% of the collection. Seed samples were obtained from the USDA-ARS Plant Genetic Resources Conservation Unit in Griffin, Georgia from the collection for sorghum anthracnose evaluation. Four of the 273 accessions were not available at the time of this study. Multiple seeds lots were available for three accessions and were included in the study as duplicate samples. Two of the duplicate accessions are part of the Mali working collection. In total, 270 Malian sorghum accessions were included in the anthracnose evaluation.
The anthracnose evaluation of the Mali working collection was conducted in 2003 at the USDA-ARS Tropical Agriculture Research Station in Isabela, Puerto Rico. The experiment was planted in a partially balanced lattice design on 28 March and 29 July 2003. The first planting date was selected to correspond to the dry season with the second planting associated with the wet season. Sixteen inbred sorghum breeding lines were selected as control samples to evaluate variation in infection response within and between experiments. Infection response for the majority of the control samples is unknown for the current pathotypes of sorghum anthracnose in Puerto Rico, but several lines were selected based on published disease response at other locations. The control samples and Malian accessions were planted in single rows, 1.8 m in length with 0.9 m row spacing and replicated three times. Border rows of the anthracnose susceptible germplasm line ‘Dorado’ surrounded the experiments. Fertilizer was applied at a rate of 560 kg ha-1 (15-5-10 NPK) at planting. Weeds were controlled by hand hoeing and mechanical tillage. Supplemental irrigation was only applied for stand establishment and no irrigation was applied after disease inoculation.
The experiments were artificially inoculated approximately 30 days after planting with anthracnose infected leaf tissue. Leaves infected with anthracnose were collected from numerous sorghum accessions in 2002 and 2003, including Dorado and BTx623, in order to represent the pathotypic variation present at the Tropical Agriculture Research Station in Puerto Rico. Anthracnose infected leaves were also collected from susceptible Malian accessions, control samples and border rows from the first experiment and used as an additional source of inoculum for the second experiment to maintain the pathotype profile between experiments. The inoculum was prepared by grinding infected leaves in a Thomas-Wiley cutting mill (A.H. Thomas Co., Philadelphia, PA) using a 1 mm screen. Plants were inoculated by placing approximately 0.5 g of infected leaf tissue in the leaf whorl. Disease evaluations of inoculated leaves were conducted approximately 30, 40 and 60 days after inoculation. Anthracnose disease assessment was based on a 1 to 5 scale modified from Pande[10], where; 1 = no symptoms or chlorotic flecks on leaves emerging from the inoculated whorl; 2 = hypersensitive reaction (reddening or red spots) on inoculated leaves with no acervuli formation; 3 = chlorotic lesions on inoculated leaves with acervuli in the center; 4 = elongated necrotic lesions with acervuli on infected leaves and spreading to other leaves and 5 = coalescence of lesions with abundant acervuli resulting in leaf death and most leaves infected including the flag leaf. Highly resistance accessions were rated as 1, resistant accessions as 2, moderately susceptible accessions as 3, susceptible accessions as 4 and highly susceptible as 5. The rating scale based on distinct disease response phenotypes was selected for accuracy, since the germplasm collection was highly variable for many traits that could contribute to inconsistent quantitative variation for disease response. The sorghum landraces in the collection were heterogeneous for anthracnose infection response; therefore, the disease phenotype that represented the majority of the plants in the row was used to rate the disease response for the Malian accessions.
The disease response at 60 days was used to conduct the statistical analysis of the data. At 60 days after inoculation, over 50% of the accessions were approaching physiological maturity and further disease progression would not influence seed development. Additionally, no changes in disease progression were observed after 60 days. Analysis of variance was conducted with Statistix software (Analytical Software, Tallahassee, FL) to determine whether disease response was significantly different for the Malian sorghum accessions.
RESULTS AND DISCUSSION
The disease response to sorghum anthracnose was obtained for 270 germplasm accessions from the Mali working collection. Table 1 presents the anthracnose disease response for each accession during the dry and wet growing seasons with the accessions organized by disease severity. During the dry season, data was only obtained from one replication of the control sample, SC727, as no seed germinated in the other two replications. Three rows in the third replication were lost to rodents for the experiment conducted during the wet season. Analysis of variance indicated highly significant (P=0.01) differences for anthracnose disease response for the Malian sorghum accessions (data not shown). Thus, anthracnose resistant accessions could be readily differentiated from susceptible accessions. The response to infection with anthracnose was generally uniform between experiments for both the accessions and control samples. Fifteen accessions evaluated in the dry season showed variation for anthracnose disease response. These accessions showed a uniform susceptible response during the wet season. The disease response for two accessions during the wet season was variable and showed a resistant response for the experiment in the dry season. These results would indicate a genotype by environment influence. During the dry season, 245 accessions were resistant compared to 215 accessions during the wet season (Table 2). Ten accessions were susceptible in the dry season compared to 53 accessions in the wet season. These results would suggest that climatic conditions influence the host and pathogen, probably arresting disease development during the dry season. Neya and Le Normand[11] also observed variation in disease response based on the climatic conditions at three locations in Burkina Faso, West Africa with the wetter locations associated with greater disease severity. Pande[12] recommended field screening during the rainy season as the most effective with disease development enhanced through the use of irrigation to increase humidity.
Disease development was also hindered for some of the control samples during the dry season.
| Table 1: | Anthracnose disease response under field conditions for 270 sorghum accessions from Mali, West Africa and 16 control samples inoculated with Colletotrichum sublineolum during the dry and wet growing seasons in Puerto Rico during 2003 |
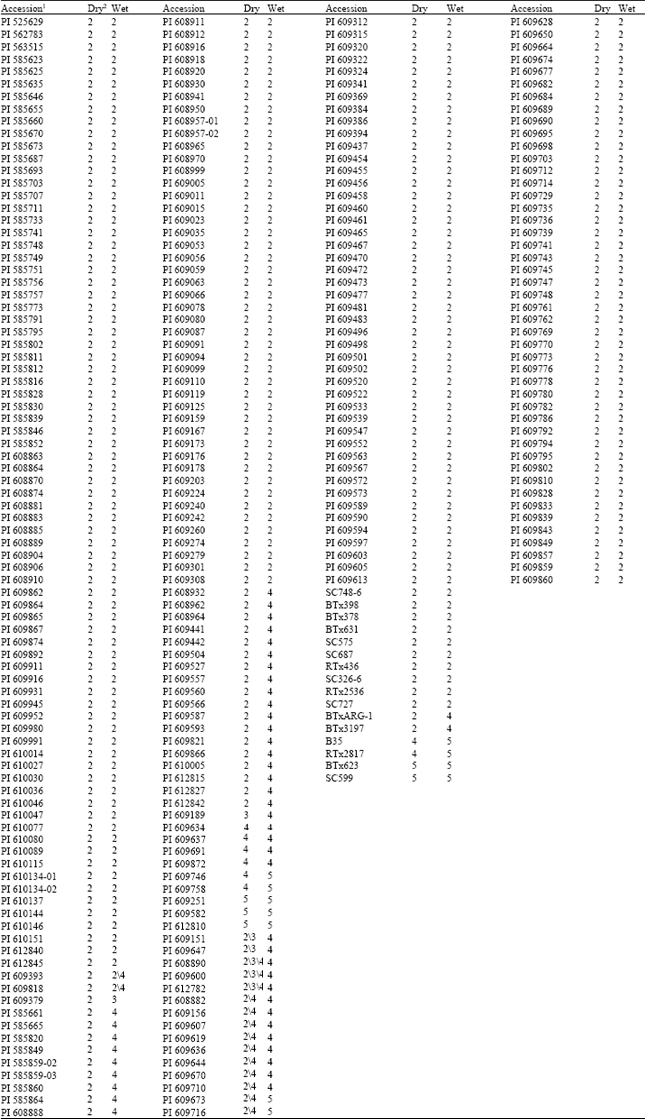 | |
| 1Accessions are listed by anthracnose disease response from resistant to susceptible. Two-digit extension indicates a duplicate accession. 2Rows with a single value indicate no variation within an experiment. An accession with more than one disease response value indicates variation within a growing season. Scale: 1 = no symptoms or chlorotic flecks on leaves emerging from the inoculated whorl; 2 = hypersensitive reaction (reddening or red spots) on inoculated leaves with no acervuli formation; 3 = chlorotic lesions on inoculated leaves with acervuli in the center; 4 = elongated necrotic lesions with acervuli on infected leaves and spreading to other leaves; and 5 = coalescence of lesions with abundant acervuli resulting in leaf death and most leaves infected including the flag leaf. | |
| Table 2: | Number of Mali sorghum accessions for each anthracnose disease rating based on disease response observed for 270 accessions during the dry and wet growing seasons in Puerto Rico during 2003 |
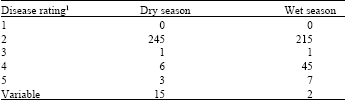 | |
| 1Scale: 1 = no symptoms or chlorotic flecks on leaves emerging from the inoculated whorl; 2 = hypersensitive reaction (reddening or red spots) on inoculated leaves with no acervuli formation; 3 = chlorotic lesions on inoculated leaves with acervuli in the center; 4 = elongated necrotic lesions with acervuli on infected leaves and spreading to other leaves and 5 = coalescence of lesions with abundant acervuli resulting in leaf death and most leaves infected including the flag leaf. Variable = anthracnose disease response varied from resistant to susceptible within an experiment | |
Two controls, BTxARG-1 and BTx3197, were rated as resistant during the dry season compared to a susceptible response during the wet season. The anthracnose disease response was uniform for the other 14 control samples with only the degree of susceptibility varying for the controls rated as susceptible. Three control samples (SC326-6, SC748-6, RTx2536) reported to have anthracnose resistance at other locations[9,21,22] were also resistant to the anthracnose pathotypes in Puerto Rico. Susceptible disease responses were observed in Puerto Rico for two control samples (B35, BTx623) that have been reported as susceptible at other locations[21,22]. The anthracnose disease response for three control samples (BTx398, BTx378, SC599) was different in Puerto Rico as compared to reported results. SC599 was susceptible in this study compared to a resistant response reported by Tenkouano[22] for anthracnose isolates from Texas and Georgia. BTx398 and BTx378 showed a resistant response to the Puerto Rican anthracnose pathotypes used in this study. In contrast, Cardwell[9] reported a susceptible disease response for BTx398 and BTx378 to anthracnose isolates from Puerto Rico, Texas and Georgia. These results could indicate that different pathotypes are present in Puerto Rico.
The influence of environmental conditions on disease response is also supported by the observation that some of the susceptible accessions during the wet season were rated as resistant 30 days after inoculation and, with disease progression, were given a susceptible rating 40 days after inoculation; whereas, no change occurred in disease response during the dry season indicating no further disease progression. The delay in disease response was also observed for susceptible accessions in the dry season as the disease rapidly infected the inoculated leaves resulting in senescence with little or no infection in non-inoculated leaves. In comparison, extensive infection was present on inoculated and non-inoculated leaves of the same accessions during the wet season. In general, disease progression proceeded rapidly from 30 to 40 days with little or no change in the disease ratings after 40 days. From 40 to 60 days, changes in disease ratings were only observed for susceptible accessions where the disease continued to progress until infection was observed in the flag leaf. Within-accession variation for disease response was observed in the dry and wet seasons for approximately 10% of the accessions. Since the sorghum germplasm collection from Mali contains landraces, within-accession variation is expected to occur for many traits. Phenotypic variation was observed for several traits including anthracnose disease response. In most cases, fewer than 20% of the plants in a row represented a different disease phenotype. Therefore, the anthracnose disease phenotype that represented the majority of the plants in the row was used to rate the disease response for the accessions and this was not a source of the variation observed within and between experiments.
The 270 Mali sorghum accessions were also evaluated for anthracnose resistance in College Station, Texas during 2003 in a non-replicated evaluation (unpublished results). A resistant response was observed for 204 accessions with 196 corresponding to accessions identified as resistant in Puerto Rico. One accession that was resistant in Puerto Rico was highly susceptible in Texas; whereas, the other 18 accessions were rated as moderately susceptible in Texas. In contrast, seven accessions rated as susceptible in Puerto Rico were resistant in Texas. The majority of the susceptible accessions observed in Texas were rated as moderately susceptible (36 accessions) compared to a susceptible or highly susceptible response in Puerto Rico (34 accessions). These results could indicate that variation in pathotypes occur between the locations. Extensive pathotypic variation has been reported to occur within and between locations for C. sublineolum[4,10,17,18]. Several studies have indicated that anthracnose isolates collected from Texas and Puerto Rico represent different pathotypes[9,15,17]. The presence of different pathotypes at each location would explain the occurrence of susceptible responses to anthracnose at one location compared to resistant responses at the other location. The anthracnose pathotypes present in Puerto Rico were considered more virulent than pathotypes in Texas by Cardwell[9], which would explain the higher susceptibility observed for 59 Malian accessions evaluated in Puerto Rico. Presently, no information is available on the virulence of the pathotypes occurring at each location. Climatic conditions may also be a factor in the variation observed. The greater susceptibility observed in Puerto Rico for 34 accessions could be attributed to the tropical climate on the island as compared to the semi-arid climate of Texas.
The inheritance of host plant resistance may also contribute to the disease response variation observed in the experiments. A preliminary evaluation of the mode of inheritance for anthracnose resistance was conducted for 41 Malian accessions identified as resistant during the evaluations (data not shown). The accessions were crossed to the anthracnose susceptible, male sterile line ATx623 and resulting hybrids were inoculated with anthracnose infected leaf tissue to evaluate infection response. A resistant response was observed for 20 hybrids indicating anthracnose resistance for the Malian parental accessions was conditioned by dominant gene action. The other 21 hybrids were susceptible suggesting recessive gene action for resistance. Dominant and recessive genes conditioning anthracnose resistance have been identified in sorghum[5,19,20], but no information is available on the genetic diversity of host resistance or allelic variation of individual disease resistant loci. The results would suggest that multiple genes for anthracnose resistance occur within the Malian sorghum germplasm collection. Additional information is need on the inheritance of anthracnose resistance in order to effectively utilize resistant germplasm.
The results show that Malian sorghum genetic resources maintained in the US National Plant Germplasm System would be a valuable source of host resistance for sorghum anthracnose. Nearly 80% of the sorghum accessions in the Mali working collection showed a uniform anthracnose resistant disease response in both the dry and wet seasons in Puerto Rico. The majority of these anthracnose resistant accessions showed a resistant disease response when evaluated in Texas, which would indicate that the germplasm should provide resistance to multiple pathotypes of the disease. Additionally, both dominant and recessive genetic factors appear to condition host resistance in different germplasm lines suggesting that genetic variation for resistance occurs within the collection that would be useful for the development of anthracnose resistant sorghum cultivars. The limited variation that was observed within and between experiments suggests that large scale, non-replicated evaluations could be effectively conducted during the dry and wet seasons in Puerto Rico as a quick screening method to identify sorghum germplasm with potential resistance to anthracnose. Selected genotypes could then be subjected to more rigorous, replicated trials and multi-location evaluations to select germplasm for sorghum improvement.
Disclaimer: Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
REFERENCES
- Freeman, S., M. Pham and R.J. Rodriguez, 1993. Molecular genotyping of Colletotrichum species based on arbitrarily primed PCR, A+T-rich DNA and nuclear DNA analyses. Exp. Mycol., 17: 309-322.
Direct Link - Sherriff, C., M.J. Whelan, G.M. Arnold and J.A. Bailey, 1995. rDNA sequence analysis confirms the distinction between Colletotrichum graminicola and C. sublineolum. Mycol. Res., 99: 475-478.
CrossRefDirect Link - Marley, P.S., R.P. Thakur and O. Ajayi, 2001. Variation among foliar isolates of Colletotrichum sublineolum of sorghum in Nigeria. Field Crops Res., 69: 133-142.
CrossRefDirect Link - Harris, H.B., B.J. Johnson, J.W. Dobson Jr. and E.S. Luttrell, 1964. Evaluation of anthracnose on grain sorghum. Crop Sci., 4: 460-462.
Direct Link - Thomas, M.D., I. Sissoko and M. Sacko, 1996. Development of leaf anthracnose and its effect on yield and grain weight of sorghum in West Africa. Plant Dis., 80: 151-153.
Direct Link - Cardwell, K.F., P.R. Hepperly and R.A. Frederiksen, 1989. Pathotypes of Colletotrichum graminicola and seed transmission of sorghum anthracnose. Plant Dis., 73: 255-257.
CrossRefDirect Link - Pande, S., L.K. Mughogho, R. Bandyopadhyay and R.I. Karunakar, 1991. Variation in pathogenicity and cultural characteristics of sorghum isolates of Colletotrichum graminicola in India. Plant Dis., 75: 778-783.
Direct Link - Neya, A. and M. le Normand, 1998. Responses of sorghum genotypes to leaf anthracnose (Colletotrichum graminicola) under field conditions in Burkina Faso. Crop Prot., 17: 47-53.
CrossRefDirect Link - Pande, S., R.P. Thakur, R.I. Karunakar, R. Bandyopadhyay and B.V.S. Reddy, 1994. Development of screening methods and identification of stable resistance to anthracnose in sorghum. Field Crops Res., 38: 157-166.
CrossRefDirect Link - Ngugi, H.K., A.M. Julian, S.B. King and B.J. Peacocke, 2000. Epidemiology of sorghum anthracnose (Colletotrichum sublineolum) and leaf blight (Exserohilum turcicum) in Kenya. Plant Pathol., 49: 129-140.
Direct Link - Thomas, M.D. and R.A. Frederiksen, 1995. Dynamics of oval and falcate conidium production of Colletotrichum graminicola from sorghum. Mycologia, 87: 87-89.
CrossRefDirect Link - Guthrie, P.A.I., C.W. Magill, R.A. Frederiksen and G.N. Odvody, 1992. Random amplified polymorphic DNA markers: A system for identifying and differentiating isolates of Colletotrichum graminicola. Phytopathology, 82: 832-835.
Direct Link - Rosewich, U.L., R.E. Pettway, B.A. McDonald, R.R. Duncan and R.A. Frederiksen, 1998. Genetic structure and temporal dynamics of a Colletotrichum graminicola population in a sorghum disease nursery. Phytopathology, 88: 1087-1093.
CrossRefDirect Link - Ali, M.E.K. and H.L. Warren, 1987. Physiological races of Colletotrichum graminicola on sorghum. Plant Dis., 71: 402-404.
CrossRef - Boora, K.S., R. Frederiksen and C.C. Magill, 1998. DNA-based markers for a recessive gene conferring anthracnose resistance in sorghum. Crop Sci., 38: 1708-1709.
Direct Link - Lo, S.C.C., K. de Verdier and R.L. Nicholson, 1999. Accumulation of 3-deoxyanthocyanidin phytoalexins and resistance to Colletotrichum sublineolum in sorghum. Physiol. Mol. Plant Pathol., 55: 263-273.
CrossRefDirect Link - Tenkouano, A., F.R. Miller, G.E. Hart, R.A. Frederiksen and R.L. Nicholson, 1993. Phytoalexin assay in juvenile sorghum: An aid to breeding for anthracnose resistance. Crop Sci., 33: 243-248.
Direct Link - Casela, C.R., A.S. Ferreira and R.E. Schaffert, 1992. Physiological Races of Colletotrichum graminicola in Brazil. In: Sorghum and Millets Diseases: A Second World Review, De Milliano, W.A.J., R.A. Frederiksen and G.D. Bengston (Eds.). International Crops Research Institute for the Semi-Arid Tropics, India, pp: 209-212.