
D. Jeena
Department of Molecular Biology and Genetic Engineering, College of Basic Sciences and Humanities, G.B. Pant University of Agriculture and Technology, Pantnagar, Uttarakhand, 263145, India
M. Baunthiyal
Department of Bioscience and Biotechnology, Banasthali University, PO Banasthali Vidyapith, Rajasthan 304 022, India
D. Pandey
Department of Molecular Biology and Genetic Engineering, College of Basic Sciences and Humanities, G.B. Pant University of Agriculture and Technology, Pantnagar, Uttarakhand, 263145, India
A.K. Gupta
Department of Molecular Biology and Genetic Engineering, College of Basic Sciences and Humanities, G.B. Pant University of Agriculture and Technology, Pantnagar, Uttarakhand, 263145, India
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 3 | Page No.: 97-112







ABSTRACT
Mitogen-Activated Protein (MAP) kinases play very important role in signal transduction during pathogen attack. We attempted to study the response of MAPK4 and MAPK6 under three different conditions i.e., with pathogen (Alternaria brassicae) only, with zeatin only and with both pathogen and zeatin. The study was performed on Arabidopsis thaliana (A. thaliana) plants. The objective was to identify the relationship between zeatin and disease resistance. To our surprise, we found an increased disease incidence in presence of pathogen at middle stage along with zeatin and also observed increased expression of MAPK4 under similar condition. MAPK6 expressions are already known to be increased during the disease incidence and in our case it has increased expression during the application of both pathogen as well as zeatin. Two-dimensional (2D) analysis of these samples revealed that the upregulated proteins were mostly in the range of 35-50 kDa range which is similar to the range of where all the MAP kinases.
PDF Abstract XML References Citation
Received: November 25, 2014;
Accepted: April 30, 2015;
Published: June 27, 2015
How to cite this article
D. Jeena, M. Baunthiyal, D. Pandey and A.K. Gupta, 2015. ATMPK4 and ATMPK6 Transcript and Protein Profiling in Arabidopsis thaliana Plants Challenged with Zeatin and Alternaria brassicae. Plant Pathology Journal, 14: 97-112.
DOI: 10.3923/ppj.2015.97.112
URL: https://scialert.net/abstract/?doi=ppj.2015.97.112
DOI: 10.3923/ppj.2015.97.112
URL: https://scialert.net/abstract/?doi=ppj.2015.97.112
INTRODUCTION
Protein phosphorylation is the most important mechanism for controlling many fundamental cellular processes in all living organisms including plants. A specific class of serine/threonine protein kinases, the Mitogen-activated Protein Kinases (MAPKs), plays a central role in the transduction of various extra and intracellular signals and is conserved throughout eukaryotes. The genome of A. thaliana contains approximately 10 MAP4K, 80 MAP3Ks, 10 MAP2Ks and 23 MAPKs. MAP kinase cascades comprise of a series of sub-families, i.e., MAP4K, MAP3K, MAP2K, MAPK, that are sequentially activated. These generally function via a cascade of networks, where MAP kinase (MAPK) is phosphorylated and activated by MAPK kinase (MAPKK or MAP2K) which itself gets activated by MAPKK kinase (MAPKKK or MAP3K). Activation of a MAP3K can occur through physical interaction or phosphorylation by the receptor itself, intermediate bridging factors or interlinking MAP4Ks. Signaling through MAP kinase cascade leads to cellular responses including cell division, differentiation as well as response to various stresses. In plants, MAP kinases are represented by multigene families and are organized into a complex network for efficient transmission of specific stimuli. Putative plant MAP kinase cascades have been postulated based on experimental analysis of in vitro interactions between specific MAP kinase components. These cascades have been tested in plants following the expression of epitope-tagged kinases in protoplasts. It is known that signaling for cell division and stress responses in plants are mediated through MAP kinases. Even auxin, abscisic acid (ABA) and possibly ethylene and cytokinin also utilize a MAP kinase pathway. Most of the biotics (pathogens and pathogen-derived elicitors) including wounding and abiotic stresses (salinity, cold, drought and oxidative) can induce defense responses in plants through MAP kinase pathways (Wrzaczek and Hirt, 2001). Average molecular mass of MAPKs ranges between 42-52 kDa (Zhang and Zhou, 2010).
Zeatin and its derivatives are the most important group of isoprenoid CKs. Zeatin occurs as two isomers; while trans-zeatin (trans-Z) has been found to be a bioactive substance, cis-zeatin (cisZ) has been reported to have a weak biological impact (Gajdosova et al., 2011). Among the different diseases caused by the genus Alternaria, blight disease is one of the most dominant one that causes average yield loss in the range of 32-57% (Conn et al., 1990). Out of 20 MAPK, three MAP kinases were characterized for disease responsive in which MAPK3 and MAPK6 are found to be as positive regulators of defense response while MAPK4 is the negative regulator of defense signaling (Pitzschke et al., 2009a) ATMPK4 and ATMPK6 are rapidly activated by low temperature, dehydration, touch, wounding and hyperosmotic stresses, but not by heat stress or ABA treatment (Ichimura et al., 2000). We focused in MAPK4 and MAPK6 in this study. We studied the expression patterns of MAPK4 and MAPK6 in the samples. Two-dimensional (2 D) separation and analysis of samples were carried out under different conditions (pathogen only, zeatin only and in presence of both pathogen and zeatin) at different time points.
MATERIALS AND METHODS
Plant material and treatments: Seeds of A. thaliana were collected from Indian Agriculture and Research Institute, Delhi. Arabidopsis thaliana ecotype Col-0 plants were sown in autoclaved soil in pots and plants were grown under conditions favorable for incidence of the disease i.e., maintaining the humidity around 85% at mean air temperature of 18-23°C and 14 h light cycle. Spore suspension was prepared for inducing the infection to A. thaliana plants. Suspension was prepared by suspending 75 spores in 100 μL of autoclaved distilled water. Infected leaves of Brassica juncea cv. Varuna exhibiting typical symptoms of Alternaria leaf spot of A. brassicae were collected from Crop Research Centre, G.B. Pant University of Agriculture and Technology, Pantnagar. It was made sure that the suspension should only contain Alternaria brassicae spore. Leaves challenged with Alternaria spores and 1 ppm zeatin phytohormone and the inoculated plants were stored in humid chamber for three to four days. At the onset of disease, the inoculated plants were transferred from humid chamber to growth chamber for proper growth and measurement of disease. Four weeks old plants were taken for this experiment. Plant samples challenged with pathogen or hormone or both were collected and stored at -80°C for RNA isolation. Expression profiling of MAPK4 and MAPK6 were performed for each type of experimental inoculation at different time intervals i.e., early stage (seven days after inoculation (DAI), 12 DAI), middle stage (17 DAI, 22 DAI) and late stage (27 DAI) for RNA isolation (Table 1).
Reverse transcriptase (RT)-PCR analysis of Arabidopsis MAPK4 and MAPK6: For RT-PCR analysis, leaves samples collected at different time intervals were subjected for NA isolation using Invitrogen kit (Invitrogen, http://www.invitrogen.com) as per the manufacturer instructions. The primers used for amplification of MAPK4 and MAPK6 were designed by using primer 3 software (Pitzschke et al., 2009a). Both MAPK and actin genes were co-amplified under the following PCR conditions: Reverse transcription at 50°C for 30 min followed by 35 cycles of amplification (94°C for 1 min, 58.2°C for 1 min and 72°C for 1 min) with final extension at 72°C for 7 min.
Densitometry analysis: After the completion of reverse transcription polymerase chain reaction (RT-PCR), the amplicons were analyzed by electrophoresing them in 1.8% agarose gel followed by quantification using the spot densitometry tool of AlphaImager software.
Two-dimensional (2D) electrophoresis: Isoelectric focusing (IEF) of protein extracts in the first dimension as well as separation by SDS-PAGE in the second dimension was performed following reported method (Garfin and Heerdt, 2000). Approximately 300 mg of protein in 300 mL of rehydration/sample buffer (8 M urea, 2% CHAPS, 40 mM dithiothreitol (DTT), 0.2% Bio-Lyte and 2 mM TATA-binding protein (TBP)) was used to passively hydrate 17 cm immobilized pH gradient (IPG) strips (pH 4-7 linear; Bio-Rad). Isoelectric focusing was performed using a Bio-Rad PROTEAN IEF unit programmed to provide an optimum, maximum field strength of 600 V cm-1 and a 50 mA limit/IPG strip at 10,000 V for 60,000 V-h. Prior to the focusing step, the strips were held at 250 V for 15 min to remove charged contaminants and after focusing, the strips were held at 500 V for 3 h to eliminate artifacts due to over and under-focusing. Prior to second dimension SDS-PAGE, in order to solubilize focused proteins and allow SDS binding, the focused IPG strips were equilibrated in buffers containing SDS and DTT for the reduction of sulfhydryl groups followed by a second incubation in buffer containing iodoacetamide which alkylates the reduced sulfhydryl groups.
| Table 1: | Characteristics of symptoms of disease developed at different DAI of pathogen in Arabidopsis thaliana under controlled condition |
 | |
Second dimension electrophoresis was carried out on 13% polyacrylamide gels (20×20 cm×1 mm thick gel) using a PROTEAN II xi system (Bio-Rad) at 45 V/gel until the dye front reached the bottom of gel and it took 2 h to reach. Gels were stained with silver using Silver Stain Plus kit (Bio-Rad) and images of the stained gels were acquired using the GS-800 calibrated densitometer (Bio-Rad). For each time-point, images from at least three gels obtained from three independent inoculation experiments were compared.
Image analysis: Two-dimensional gels from the protein extracts of control and inoculated leaf tissues were analyzed using the PDQuest software (Bio-Rad). Gels from three independent biological replicates were used to make the match-sets and individual spots were matched (added or deleted) using software tools. Each set of gels from three replicates were analyzed simultaneously using Student’s t-test feature of this software in order to identify protein spots which indicate statistically significant differences in levels as a result of pathogen challenge. Spot intensities of control and inoculated gels at different time points were determined using the spot quantification tool and the fold changes from controls were calculated. These spots were excised from the gels using a sterile scalpel and the proteins were identified using ESI-Q-ToFMS/MS.
RESULTS
Inoculation of Arabidopsis thaliana with pathogen and phytohormone: One month old healthy Arabidopsis plants were treated with the pathogen and phytohormone individually and then with 1 ppm Zeatin followed by inoculation with A. brassicae pathogen after 24 h by spraying the 10 mL spore suspension over the leaves with the help of an atomizer. First symptom of infection was seen in the form of very small necrotic spot after seven days of inoculation which gradually increased to give rise chlorotic and necrotic lesions at the later stages. These symptoms resembled to the typical symptoms as seen in case of pathogenesis of Alternaria blight in Brassica. When infected leaf samples were crushed in distilled water to prepare suspension and checked under compound microscope, viable Alternaria brassicae spores. Inoculated plants were incubated for 24 h in humid chamber at 80% RH at 22-24°C for the development of symptoms.
Arabidopsis were tested for reaction against the pathogen Alternaria blight and the phytohormone zeatin individually as well as in presence of both the pathogen and hormone together under controlled condition (glass house). The observation was based on the following parameters:
Study of symptoms: At four DAI, first symptoms in the form of black minute spots were observed in pathogen inoculated and Pathogen+Zeatin inoculated plants while symptoms were not found in zeatin inoculated plants (Fig. 1, Table 2). At seven DAI, the spots were looked somewhat round in pathogen treated leaves and the pathogen+zeatin treated leaves, spots looked somewhat bigger in comparison to the former. However, symptoms were still not found in leaves challenged only with zeatin. At 12 DAI while round medium spots were observed in pathogen treated leaves, round bigger spots were appeared in pathogen+zeatin treated leaves. Zeatin treated leaves still did not show any infection even after 12 DAI. At 17 DAI, clear concentric rings were observed in pathogen+zeatin treated leaves while of pathogen treated leaves showed no such concentric ring; no appearance of infection in zeatin treated leaves. At 22 DAI while large spots and well-developed concentric rings appeared in pathogen+Zeatin treated leaves, spots were collapsed and yellow halo formation appeared in pathogen treated leaves; no infection appeared in zeatin treated leaves. It was found that in pathogen treated plant the shape of spots gradually became irregular due to fusion of spots and there was yellow halo formation and onset of chlorosis at 27 DAI in pathogen treated while. In pathogen+zeatin treated plants we found were collapsed and yellow halo formation appeared. Therefore, Arabidopsis showed development of characteristic symptoms of Alternaria blight while inoculated with pathogen alone and pathogen and zeatin together but zeatin treated Arabidopsis plants did not develop any such characteristic symptoms (Table 3, 4 and 5).
Isolation of total RNA and expression profiling of MAP kinases: Total RNA was isolated from the leaves of the host plants at different stages of infection using invitrogen plant reagent. Isolated RNA was treated with DNase and the resulting RNA was electrophoresed in 1.2% denaturing agarose gel. Three RNA bands with smearing indicated isolation of good quality of total RNA. Concentration of each sample was determined spectrophotometrically at 260 nm. The ratio of absorbances at 260 and 280 nm (OD260/OD280) is close to 2.0.
Expression profiling of MAPK-4 and MAPK-6 genes: RT-PCR was performed using isolated RNA from infected leaves of the host plants. The primer set of MAPK-4 and MAPK-6 partial gene product (243 and 161 bp) were designed to amplify MAP kinases and to perform expression analysis. For expression analysis of MAPK4 and MAPK6, semi-quantitative RT-PCR was performed on total RNA, isolated from leaves at different stages of disease progression, using actin gene as the internal standards. The RT-PCR yielded expected amplicons of 243 and 161 bp for MAPK-4 and MAPK-6, respectively. Actin was also co-amplified along with the amplicon of expected size (172 bp) at 58.2°C after semi-quantitative RT-PCR product was checked with 1.8% agarose gel. Earlier our group showed that A. thaliana acts as host for the disease and expression of MAPK-4 and MAPK-6 are inversely related during pathogenesis of Alternaria blight. It has also been shown that MAPK-6 mutant has a compromised disease resistance, whereas MAPK4 mutant has increased resistance to pathogens. The amplified product sequence was obtained by direct sequencing using the automated DNA sequencing.
 | |
| Fig. 1(a-l): | Healthy leaves, pathogen, pathogen with zeatin and alone zeatin treated leaves. (a-d) Health leaves, (e-h) Pathogen (Alternaria brassicae) treated leaves and (i-l) Pathogen (Alternaria brassicae)+1 ppm zeatin treated leaves |
| Table 2: | Description of stages of leaves having different grades of infection |
 | |
| Table 3: | Average size of Alternaria spots (cm) on leaves of genotype Arabidopsis thaliana inoculated with pathogen and pathogen+zeatin under controlled conditions |
 | |
| *Average of 30 leaves (10 leaves in each replication) | |
| Table 4: | Average number of Alternaria spots/9 cm2 leaf area of genotype Arabidopsis thaliana inoculated with pathogen and pathogen+zeatin under controlled conditions |
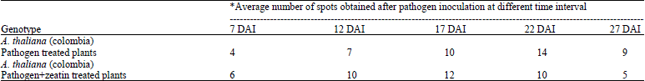 | |
| *Average of 30 leaves (10 leaves in each replication) | |
The obtained sequences were subjected to homology search using blast. It was found that sequence having 99% identity with A. thaliana mRNA, MAPK-4 and MAPK-6.
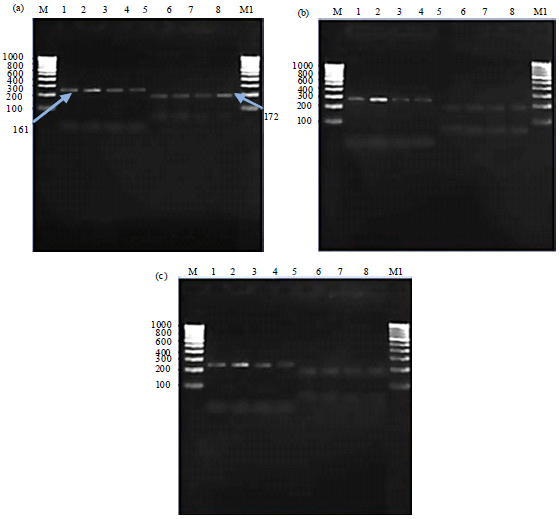 | |
| Fig. 2(a-c): | RT-PCR analysis of MAPK4 under different treatments, (a) Pathogen, (b) Pathogen+zeatin and (c) Zeatin |
| Table 5: | Determination of concentration of protein in different leaves samples of host species of A. thaliana after the challenge of A. brassicae, brassicae with zeatin and zeatin alone |
 | |
Expression profiling of MAPK-4 from pathogen and phytohormone challenged leaves at different stages: Temporal expression analysis of MAPK-4 was performed during pathogenesis of Alternaria blight alone, Alternaria in combination of phytohormone (zeatine) and phytohormone (zeatine) alone. For this, RT-PCR was performed on total RNA, isolated from Arabidopsis leaves having constant disease severity but harvested at different DAI, using actin gene as the internal standard. Actin showing a constitutive expression with all the samples indicated that there was no difference in the quantity of RNA in different samples. However, the gene MAPK-4 was found to be downregulated in infected leaf samples compared to healthy samples (Fig. 2). This may be triggered in response to signaling pathway by A. brassicae pathogen to negate the infection process.
 | |
| Fig. 3(a-c): | Densitometry analysis of MAPK4 gene under different conditions, (a) Pathogen, (b) Pathogen+zeatin and (c) Zeatin |
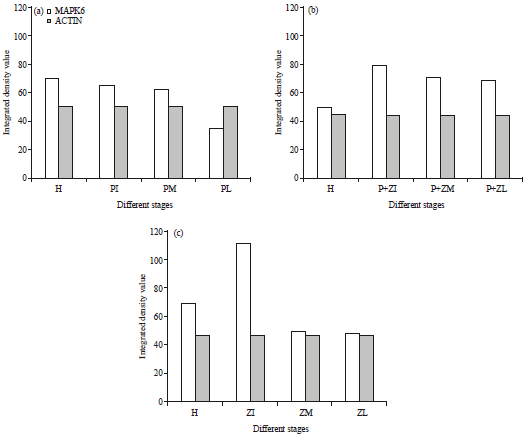
When the host plant was treated A. brassicae together with zeatin, the MAPK expression varied at different stages. MAPK-4 was expressed at early, middle, late and also in control. Densitometry analysis showed that MAPK-4 expression was found maximum in P+ZI, the early stage and thereafter, expression decreased till P+ZL, the late stage (Fig. 3).
MAPK-4 was found expressed in control, early, middle and late when A. thaliana was inoculated with zeatin. The densitometry analysis clearly demonstrated that the expression level of MAPK-4 was varied in all the treated samples and found the maximum at ZI, the early stage after zeatin treatment. Thereafter, the expression decreased at ZM and gradually became constant. The expression levels, ZM and ZL, are same but low compared to ZI and control.
Expression profiling of MAPK-6 from pathogen and phytohormone challenged leaves at different stages: The transcript profiling of MAPK-6 during different stages of disease progression, we performed RT-PCR analysis on RNA samples isolated from leaves of Arabidopsis at different stages of disease progression. During different stages of disease progression, the expression of MAPK-6 gradually increased from initial to late stage (Fig. 4). The present results suggest that the pathogen facilitates the fungal colonization by continuous signaling to upregulate MAPK-6 during pathogenesis. This was demonstrated by the experiment in which expression of the pathogen-inducible gene vegetative storage protein1 (VSP1) in MPK6-silenced plants was several fold lower than in control plants. In A. thaliana plant, MAPK- 6 amplification was found at different stages (A. brassicae/A. brassicae with Zeatin/Zeatin challenged) including in the controls (healthy) while in all the cases, actin was amplified and showed band of expected size i.e., 172 bp.
In A. thaliana, MAPK-6 was expressed in control, early, middle and late infection. Densitometry analysis clearly showed that the expression of MAPK-6 increased after pathogen (A. brassicae) infection. MAPK-4 expression maximum was observed at early stage of pathogen PI. Thereafter, it decreased at middle stage of pathogen PM; however, at PL, the late stage of pathogen, it again increased further. In control, very little expression of gene was seen.
In A. thaliana plant, the MAPK-6 expression was found at early, mid and late stages of disease progression. The expression of MAPK-6 was found to increase in pathogen with zeatin treated leaves at different stages. The minimum expression was found in control HL; however, maximum at P+ZL, the pathogen with zeatin late stages. The densitometric analysis done by alpha imager was demonstrated that actin level remained almost same at all time intervals (Fig. 5).
In zeatin treated leaves of A. thaliana plant showed different pattern of MAPK-6 expression. MAPK-6 expression was found at early, mid and late stages of disease progression. In this case, the expression was found maximum in zeatin late stage (ZL) and the minimum expression was found in zeatin middle stage.
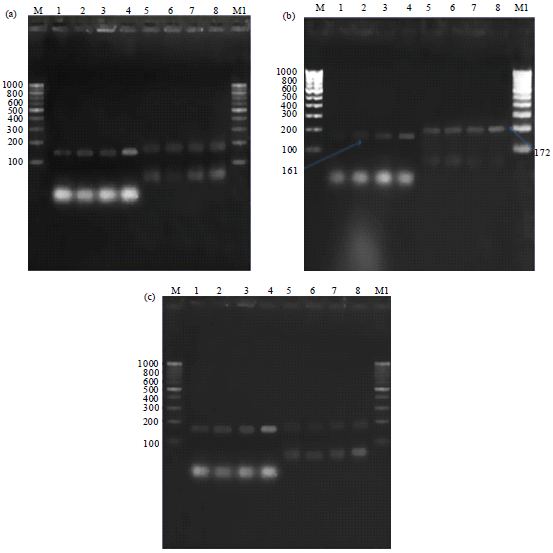 | |
| Fig. 4(a-c): | RT-PCR analysis of MAPK6 under different treatments, (a) Pathogen, (b) Pathogen+zeatin and (c) Zeatin |
Isolation of total protein from the host plant (A. thaliana): Total protein was extracted from the leaves of A. thaliana plant at healthy (control) and subsequently, at different stages of disease progression under different condition. The amount of protein was estimated by Bradford method in each case. The concentration of protein obtained from different leaf samples is given in Table 5.
Alteration of protein profiling during different stages of infection: In order to study the differential expression of proteins induced in response to pathogenesis, proteins isolated from infected leaves at different stages (early, mid, late) were subjected to SDS-PAGE (12.5%) electrophoresis (Fig. 6).
In pathogen treated samples, total 16 bands were found at late stage while total 17 bands in control. A 22 kD band appeared during early, mid and late stages of disease (marked by circle). Bands at 43 and 22 kDa disappeared during late stages of disease. Band at 45 kDa was only found in control sample. The bands at 36 and 27 kDa band were present only pathogens treated samples but absent in control. The band at 43 kDa was absent at middle stage.
Total 17 bands were found in samples treated with pathogen and zeatin together at late stage compared to control. The bands at 28 and 22 kDa appeared during early, mid and late stages of disease (marked by circle). These bands showed decrease in disease progression. Two bands at 33 and 25 kDa were absent in control.
Total 15 bands were found in the zeatin treated samples at late stage compared to control healthy samples. A band at 22 kD band appeared during early, mid and late stages of disease (marked by circle). At the mid and late stages of disease progression, two bands at 28 and 25 kDa appeared (marked by circle) in only zeatin treated sample but absent in control. However, bands at 21 kDa were absent in zeatin treated sample.
Determination of total protein by 2D gel electrophoresis: Two dimensional gel electrophoresis of total protein of host spp of A. thaliana was performed for the leaf samples collected at different stages of disease progression. The details of samples sent for custom services of 2D analysis are listed in Table 6. After 2D gel electrophoresis, gels were analyzed using PD quest software.
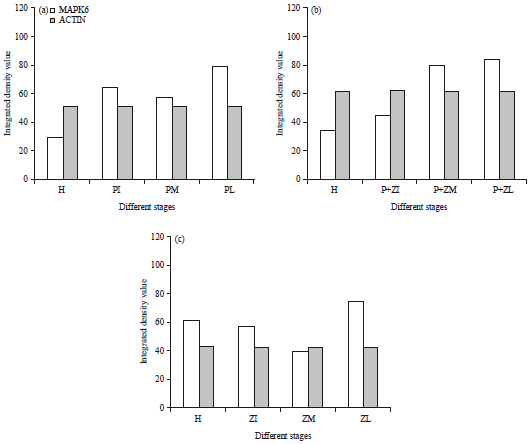 | |
| Fig. 5(a-c): | Densitometry analysis of MAPK gene under different conditions, (a) Pathogen, (b) Pathogen+zeatin and (c) Zeatin |
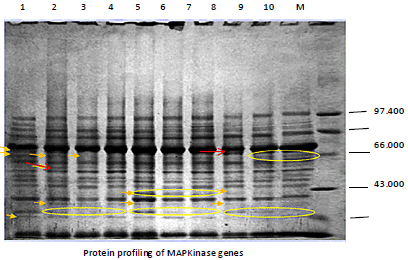 | |
| Fig. 6: | Determination of concentration of protein in different leaves samples of host spp. of A. thaliana challenged with A. brassicae alone, A. brassicae with zeatin and zeatin alone, Lane 1: Healthy leave, Lane 2: Pathogen early leave, Lane 3: Pathogen middle leave, Lane 4: Pathogen late leave, Lane 5: Pathogen+zeatin early leave, Lane 6: Pathogen+zeatin middle leave, Lane 7: Pathogen+zeatin late leave, Lane 8: Zeatin early, Lane 9: Zeatin middle leave, Lane 10: Zeatin late leave, M: Middle range molecular marker |
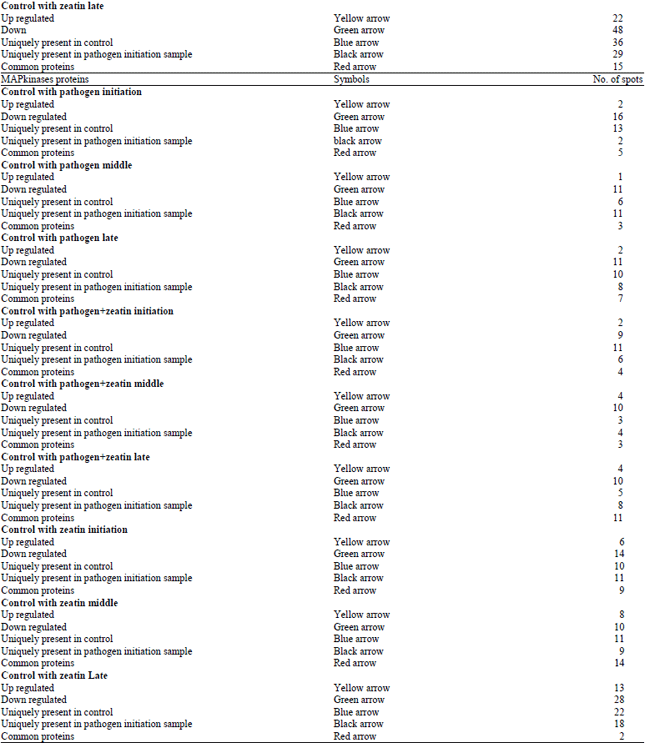
By PD quest, scattered plots of different compared pair was plotted to determine the number of matched proteins down regulated /up regulated as well as the number of proteins remained constant. Finally, the differentially expressed proteins were marked in gel by arrows of different colors: Green arrow for down regulated proteins, yellow arrow for up regulated proteins, blue arrow for proteins present only in control, black arrow for proteins uniquely expressed in treated sample and red arrow for protein present the same expression in both control and treated samples (Table 7).
| Table 6: | 2D gel electrophoresis of proteins isolated from Alternaria brassicae challenged and phytohormone zeatin treated leaves at different stages of infection |
 | |
| Table 7: | All the kinases range between 32-52 kD so protein spots were observed in this region 32-52 kD was observed and represented by black circle in the gel |
  | |
2D gel analysis of pathogen (A. brassicae) treated samples: The different set of gels which was compared is listed in Table 7.
Comparison between HL-PI: Results of PD quest analysis clearly demonstrated that there was difference in protein expression at all the stages in comparison to control.
 | |
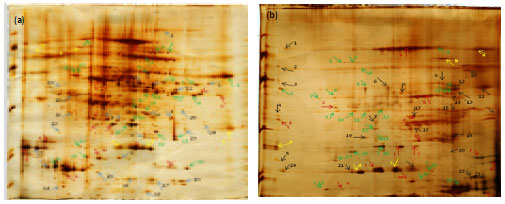
| Fig. 7(a-b): | Comparison between HL-PI |
 | |
| Fig. 8(a-b): | Comparison between HL-PM |
A clear difference in protein pattern was found upon comparing control HL at early stage of pathogen treated samples (PI). By comparing visible spots, 16 proteins were found upregulated (A-F yellow arrow), 40 were downregulated (1-40 green arrow), 40 were present only in control (1-40 blue arrow), 15 were uniquely appeared in PI after pathogen challenged (1-15 black arrow) and 17 were the same expressed protein (A-Q red arrow). Because all the kinases ranges between 35-52 kD, therefore, protein spots in this region was observed and represented by purple circle in gel. We also found that in 32-52 kD range, two proteins were upregulated protein, 16 were downregulated, 13 proteins were only present in control while two proteins appeared after pathogen challenge and five same expressed proteins. One cluster of protein (marked by dark blue circle) was found uniquely appeared in PI after pathogen challenge and the other cluster of downregulated proteins was found (Fig. 7).
Comparison between HL-PM: In this case five proteins (a-e yellow arrow) were upregulated, 23 (1-23 green arrow) were downregulated, 25 proteins (1-25 blue arrow) were only present in control, 24 proteins (1-24 black arrow) were uniquely appeared at early stage of infection and 14 proteins (a-n red arrow) the same expression. A cluster of proteins (marked by dark blue circle) was found uniquely appeared in PI after pathogen challenge. In the 35-52 kD range (marked by purple circle), 11 proteins were found downregulated, six were only present in control; however, 11 proteins were appeared after pathogen challenge while three were the same expression (Fig. 8).
Comparison between HL-PL: In this case, five proteins (a-e yellow arrow) were upregulated, 22 proteins (1-22 green arrow) were downregulated, 22 proteins (1-22 blue arrow) were present only in control, 28 proteins (1-28 black arrow) were appeared uniquely in PL i.e., at late stage of disease and 17 (a-q red arrow) proteins expressed the same expression. It was found that out of total protein spots present in the range of 35-52 kD (marked by purple circle), one protein was upregulated, 11 were downregulated, 10 proteins present in control, six were present uniquely in PL and seven were the same expressed proteins as marked by purple circle (Fig. 9).
Comparison between HL-P+ZI: The results of comparison between HL-P+ZI demonstrated that 10 proteins (a-j yellow arrow) were upregulated, 14 proteins (1-14 green arrow) were downregulated, 25 proteins (1-25 blue arrow) only present in control and 26 proteins (1-26 black arrow) were uniquely expressed in P+ZI after combined pathogen and phytohormone challenge while only (1 red arrow) one protein showed the same expression. Two clusters of proteins (marked by dark blue circle) were found uniquely appeared in P+ZI after combined pathogen and zeatin challenges.
 | |
| Fig. 9(a-b): | Comparison between HL-PL |
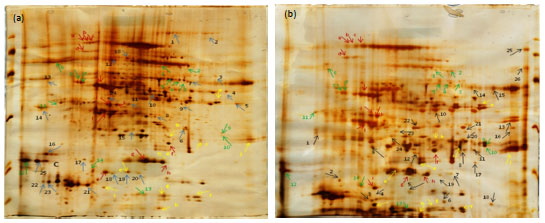 | |
| Fig. 10(a-b): | Comparison between HL-P+ZI |
We also found that out of total protein spots present in the range of 35-52 kD (marked by purple circle), one protein was found upregulated, nine were downregulated, 12 proteins were only found in control, five proteins were uniquely expressed in P+ZI and four showed the same expression present in the range between 35-52 kD (Fig. 10).
Comparison between HL-P+ZM: In this case, we found that six proteins (a-f yellow arrow) were upregulated, 25 proteins (1-25 green arrow) were downregulated, 14 proteins (1-14 blue arrow) were present only in control; however, 21 proteins (1-21 black arrow) uniquely expressed in P+ZM pathogen and zeatin treatment while 14 proteins (a-n red arrow) showed same expression in both HL and P+ZM. Among the total protein spots present in the range of 35-52 KD (marked by purple circle), four proteins were upregulated, 10 were downregulated, three proteins were only found in control, three were uniquely expressed protein in P+ZM and three showed the same expressed in both control and combined pathogen and zeatin challenged samples (Fig. 11).
Comparison between HL-P+ZL: The results of this pair indicated that nine proteins (a-i yellow arrow) were upregulated, 12 proteins (1-12 green arrow) were downregulated, seven proteins (1-7 blue arrow) were only present in control, 18 proteins (1-18 black arrow) were uniquely expressed in pathogen with zeatin treatment; however, 14 proteins (a-n red arrow) were the same expression in both control and pathogen with zeatin treated samples. Out of total protein spots, three were upregulated proteins, 10 were downregulated protein, five were present in control, four were uniquely present in P+ZL and 11 showed the same expression in both HL and P+ZL in the ranges between 32-52 kD (marked by purple circle) (Fig. 12).
Comparison between HL-ZI: In this case, 20 proteins (a-t yellow arrow) were upregulated, 24 (1-24 green arrow) were downregulated, 22 proteins (1-22 blue arrow) were only present in control, 17 proteins (1-17 black arrow) were uniquely appeared in zeatin treated samples and 16 proteins (a-p red arrow) were uniquely appeared at HL-ZI after zeatin treatment. In the range of 35-52 kD, (marked purple circle) we found that two proteins were upregulated, 14 were downregulated, 10 were only present in control, 11 proteins were appeared after zeatin treatment; however, nine were uniquely expressed in both HL-ZI (Fig. 13).
Comparison between HL-ZM: A clear difference in protein pattern was found while comparing HL with ZM.
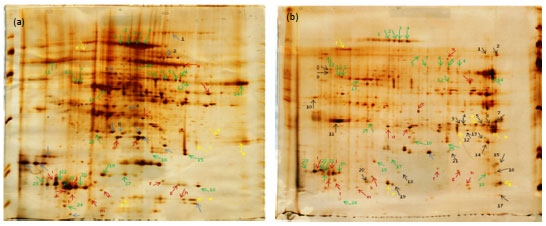 | |
| Fig. 11(a-b): | Comparison between HL-P+ZM |
 | |
| Fig. 12(a-b): | Comparison between HL-P+ZL |
 | |
| Fig. 13(a-b): | Comparison between HL-ZI |
By comparing visible spots, we found that 29 proteins were upregulated (a-z yellow arrow), 32 were downregulated (1-32 green arrow), 21 were present only in control (1-21 blue arrow), 36 (1-36 black arrow) were uniquely appeared in ZM and 29 (a-z red arrow) proteins uniquely expressed in both HL-ZM. A cluster of down regulated proteins marked by dark blue color circle was uniquely expressed in ZM after zeatin treatment. Since all the kinases ranges between 35-52 kD, the protein spots observed in this region are represented by purple circle in gel. We found, in the 32-52 kD range, eight upregulated proteins, 10 downregulated proteins, 11 proteins were only present in control while nine proteins appeared in ZM and 14 proteins in HL-ZM (Fig. 14).
Comparison between HL-ZL: Comparison between HL and ZL revealed that 22 (a-v yellow arrow) proteins were upregulated, 48 proteins (1-48 green arrow) were downregulated, 36 proteins (1-36 blue arrow) were present only in control, 29 (1-29 black arrow) were appeared uniquely in ZL i.e., at late stage of disease and 15 proteins (a-o red arrow) were uniquely expressed proteins in both HL-ZL.
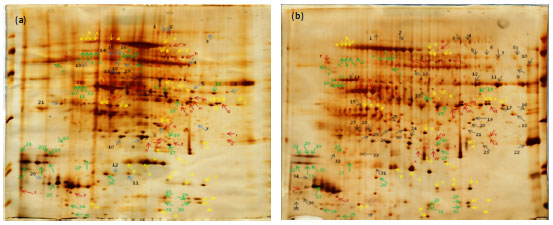 | |
| Fig. 14(a-b): | Comparison between HL-ZM |
 | |
| Fig. 15(a-b): | Comparison between HL-ZL |
Two clusters of down regulated proteins, marked by dark blue color circle, were uniquely expressed in HL. Another one cluster of upregulated proteins and one cluster of downregulated proteins were expressed in ZL after zeatin treatment marked by dark blue color circle. Out of total protein spots present in the range of 35-52 kD, 13 proteins were up regulated, 28 were down regulated, 22 proteins present in control, 18 were present uniquely in ZL and only one protein was uniquely expressed in both HL-ZL as marked by black circle (Fig. 15).
DISCUSSION
We found that phytohormone zeatin only failed to induce any infection in the host plant, A. thaliana at any stage. However, the incidence of disease gradually increased with time in the plants inoculated with only pathogen as well as with pathogen together with the zeatin (Table 3). Interestingly, the DI is high in the plants inoculated with pathogen and zeatin together. Novak et al. (2013) reported that hypersensitive response is mediated through cytokinins. In this case hypersensitivity was created only in combination of both cytokinin and pathogen high levels of cytokinins to trigger fast cell death without any intervening of chlorosis -a hallmark of the hypersensitive response. The results suggest that chloroplastic hydrogen peroxide orchestrates the molecular responses underpinning the hypersensitive-like response, including the inhibition of photosynthesis, elevated levels of stress hormones, oxidative membrane damage and stomatal closure (Novak et al., 2013).
We also found expression of MAPK-4 was downregulated in plants challenged with only pathogen, whereas the expression of MAPK-4 was high in case of zeatin alone as well as pathogen and zeatin together. It has earlier been reported that MPK-4 is required for cytokinin synthesis. In addition to participation in the defense responses, the activity of MAPK4 responds to various external stimuli such as infection by pathogens and treatment with the bacterial elicitor flagellin (Meszaros et al., 2006), cold stress and salt stress (Teige et al., 2004). In response to flagellin, the MAPKK Mkk1 gets involved in the activation of MPK4 while MAPKK Mkk2 directly activates MPK4 in the response to cold stress and salt stress. Ichimura et al. (2006) found that activation of MPK4 requires the MAPKKK Mekk1 which negatively regulates a specific type of cell death. Reverse genetic analysis of the Arabidopsis MPK-4 knockout mutant revealed its importance in regulating systemic acquired resistance, including the ability to accumulate salicylic acid (Petersen et al., 2000).
Pathogen Associated Molecular Patterns (PAMP)-triggered immunity (PTI) constitutes the first layer of plant immunity that restricts pathogen proliferation MPK4 and its upstream MAP kinase kinases (MKK1 and MKK2) and the MAP kinase kinase kinase (MEKK1) form a cascade that negatively regulates defenses in Arabidopsis, because loss-of-function mutations in this cascade results in constitutive activation of defenses and dwarfed plants (Ichimura et al., 2006; Meszaros et al., 2006; Suarez-Rodriguez et al., 2007; Gao et al., 2008; Qiu et al., 2008a; Pitzschke et al., 2009b). How MPK4 regulates plant immunity remains largely unknown. Nonetheless, MPK4 has been reported to interact with its substrate MKS1; the latter interacts with WRKY transcription factors, WRKY25 and WRKY33 (Andreasson et al., 2005). MPK4, MKS1 and WRKY33 form a complex in the nucleus and the flg22-induced MPK4 activation has been shown to release WRKY33 from the complex. This enables WRKY33 to directly activate the transcription of PAD3 which encodes a cytochrome P450 involved in camalexin biosynthesis (Qiu et al., 2008b). However, the functions of MKS1 andWRKY33 do not appear to account for the dramatic phenotype of the loss-of-function MPK4 mutants. Other substrates or interacting proteins mediating MPK4 function remain to be identified. A recent study showed that the P. syringae effector AvrB interacts with and stimulates the activity of MPK4, thereby perturbing hormone signaling and enhancing plant susceptibility in the absence of cognate R proteins RPM1 and TAO1 (Cui et al., 2010).
The activation of the MPK4 pathway by PAMPs suggests that certain PAMP-activated processes might play a strategic role to repress defenses in favor of the pathogen or to reduce detrimental effects for the plant. The hypothesis that the activation of the MPK4 pathway benefits the pathogen is also corroborated by the finding that the P. syringae effector avrB directly activates MPK4 and thereby increases Arabidopsis susceptibility to Pst DC3000 infection (Cui et al., 2010). Overall, these results contradict the common belief that PAMP-induced signaling enhances resistance against microbial infection and show that certain PAMP-induced immune signaling events are aimed at dampening plant immunity. The expression of MAPK-6 is different from that of MAPK-4. MAPK-6 is always induced in disease condition and also previously reported in several cases. In addition, harpin and H2O2 activate AtMPK4/6 and AtMPK6, respectively, in leaves in a similar manner to that seen with suspension cultures (Desikan et al., 2001).
In case of 2D analysis, we found all the kinases ranged between 35-52 kD protein spots. Hence, when we considered those spots along the lines of upregulation in pathogen only case at three temporal stages (i.e., early, middle and late), the upregulated proteins were 2, 5 and 22, respectively; in case of zeatin only at three temporal stages, the upregulated proteins were 2, 8 and 13, respectively; in case of pathogen together with zeatin at three temporal stages, the upregulated proteins were 1, 4 and 3, respectively. This is in contrast to gene expression at protein level. The proteins ranging between 35- 52 kD were found to be more in pathogen only treated leaves, whereas very low presence of those protein was found in case of pathogen+zeatin treated leaves. Therefore, the combination of pathogen together with phytohormone zeatin proved to make plant more susceptible to disease which was yet to be known.
Further investigations are required to elucidate the relationship between the phytohormone zeatin and MAP kinases mutant. Mass spectrometry analysis can shed more light on the proteins which are responding to different conditions.
ACKNOWLEDGMENT
The authors are thankful to the University for their Infrastructural Support to carry out this study.
REFERENCES
- Andreasson, E., T. Jenkins, P. Brodersen, S. Thorgrimsen and N.H. Petersen et al., 2005. The MAP kinase substrate MKS1 is a regulator of plant defense responses. EMBO J., 24: 2579-2589.
CrossRefDirect Link - Conn, K.L., J.P. Tewari and R.P. Awasthi, 1990. A disease assessment key for Alternaria blackspot in rapeseed and mustard. Can. Plant Dis. Surv., 70: 19-22.
Direct Link - Cui, H., Y. Wang, L. Xue, J. Chu and C. Yan et al., 2010. Pseudomonas syringae effector protein AvrB perturbs Arabidopsis hormone signaling by activating MAP kinase 4. Cell Host Microbe, 7: 164-175.
CrossRefDirect Link - Desikan, R., J.T. Hancock, K. Ichimura, K. Shinozaki and S.J. Neill, 2001. Harpin induces activation of the Arabidopsis mitogen-activated protein kinases AtMPK4 and AtMPK6. Plant Physiol., 126: 1579-1587.
Direct Link - Gajdosova, S., L. Spichal, M. Kaminek, K. Hoyerova and O. Novak et al., 2011. Distribution, biological activities, metabolism and the conceivable function of cis-zeatin-type cytokinins in plants. J. Exp. Bot., 62: 2827-2840.
CrossRefDirect Link - Gao, M., J. Liu, D. Bi, Z. Zhang, F. Cheng, S. Chen and Y. Zhang, 2008. MEKK1, MKK1/MKK2 and MPK4 function together in a mitogen-activated protein kinase cascade to regulate innate immunity in plants. Cell Res., 18: 1190-1198.
CrossRefDirect Link - Ichimura, K., C. Casais, S.C. Peck, K. Shinozaki and K. Shirasu, 2006. MEKK1 is required for MPK4 activation and regulates tissue-specific and temperature-dependent cell death in Arabidopsis. J. Biol. Chem., 281: 36969-36976.
CrossRefDirect Link - Ichimura, K., T. Mizoguchi, R. Yoshida, T. Yuasa and K. Shinozaki, 2000. Various abiotic stresses rapidly activate Arabidopsis MAP kinases ATMPK4 and ATMPK6. Plant J., 24: 655-665.
CrossRefDirect Link - Meszaros, T., A. Helfer, E. Hatzimasoura, Z. Magyar and L. Serazetdinova et al., 2006. The Arabidopsis MAP kinase kinase MKK1 participates in defence responses to the bacterial elicitor flagellin. Plant J., 48: 485-498.
CrossRefDirect Link - Novak, J., J. Pavlu, O. Novak, V. Nozkova-Hlavackova and M. Spundova et al., 2013. High cytokinin levels induce a hypersensitive-like response in tobacco. Ann. Bot., 112: 41-55.
CrossRefDirect Link - Petersen, M., P. Brodersen, H. Naested, E. Andreasson and U. Lindhart et al., 2000. Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance. Cell, 103: 1111-1120.
CrossRefDirect Link - Pitzschke, A., A. Schikora and H. Hirt, 2009. MAPK cascade signalling networks in plant defence. Curr. Opin. Plant Biol., 12: 421-426.
CrossRefPubMedDirect Link - Qiu, J.L., B.K. Fiil, K. Petersen, H.B. Nielsen and C.J. Botanga et al., 2008. Arabidopsis MAP kinase 4 regulates gene expression through transcription factor release in the nucleus. EMBO J., 27: 2214-2221.
CrossRefDirect Link - Qiu, J.L., L. Zhou, B.W. Yun, H.B. Nielsen and B.K. Fiil et al., 2008. Arabidopsis mitogen-activated protein kinase kinases MKK1 and MKK2 have overlapping functions in defense signaling mediated by MEKK1, MPK4 and MKS1. Plant Physiol., 148: 212-222.
CrossRefPubMedDirect Link - Suarez-Rodriguez, M.C., L. Adams-Phillips, Y. Liu, H. Wang and S.H. Su et al., 2007. MEKK1 is required for flg22-induced MPK4 activation in Arabidopsis plants. Plant Physiol., 143: 661-669.
Direct Link - Teige, M., E. Scheikl, T. Eulgem, R. Doczi and K. Ichimura et al., 2004. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis. Mol. Cells, 15: 141-152.
CrossRefPubMedDirect Link - Wrzaczek, M. and H. Hirt, 2001. Plant MAP kinase pathways: How many and what for? Biol. Cell, 93: 81-87.
CrossRefDirect Link - Zhang, J. and J.M. Zhou, 2010. Plant immunity triggered by microbial molecular signatures. Mol. Plant, 3: 783-793.
CrossRefDirect Link