
Soad Hassan Taha
Department of Dairy Science, Faculty of Agriculture, Cairo University, El Gamaa Street, 12613 Giza, Egypt
LiveDNA: 20.29699
Ibrahim Mohamed El-Sherbiny
Nanomaterials Lab, Center of Material Science, Zewail City of Science and Technology, 6th of October, 12588 Giza, Egypt
Aida Soliman Salem
Department of Dairy Technology, Animal Production Research Institute, Agricultural Research Center, Giza, Egypt
Mahmoud Abdel-Hamid
Department of Dairy Science, Faculty of Agriculture, Cairo University, El Gamaa Street, 12613 Giza, Egypt
LiveDNA: 20.30040
Ali Hussein Hamed
Department of Virus and Phytoplasma Research, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
Ghada Abady Ahmed
Department of Dairy Technology, Animal Production Research Institute, Agricultural Research Center, Giza, Egypt
Pakistan Journal of Biological Sciences
Year: 2019 | Volume: 22 | Issue: 12 | Page No.: 614-622







ABSTRACT
Background and Objective: Potato is one of the world’s leading vegetable crops. Potato viral diseases cause adversely effects on the agricultural sector. Recently there is a growing interest to control plant viruses using spices and herbs (including curcumin). Poor solubility of curcumin in water limited its applications. Therefore, the main objective of the present study was to evaluate the effect of antiviral activity of curcumin-milk proteins nanoparticles against potato virus Y (PVY). Materials and Methods: Curcumin-milk proteins nanoparticles were prepared via ionic gelation method. The antiviral activity of the resultant nanoparticles against PVY was evaluated at different concentrations (500, 1000 and 1500 mg/100 mL). Chlorophyll content as well as the activity of peroxidase (POX) and polyphenol oxidase (PPO) was examined. Results: Curcumin-milk proteins nanoparticles showed inhibitory effect on PVY in a concentration dependent manner. Conclusion: Curcumin-milk proteins nanoparticles displayed a successful tool to control the PVY under green house conditions.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Soad Hassan Taha, Ibrahim Mohamed El-Sherbiny, Aida Soliman Salem, Mahmoud Abdel-Hamid, Ali Hussein Hamed and Ghada Abady Ahmed, 2019. Antiviral Activity of Curcumin Loaded Milk Proteins Nanoparticles on Potato Virus Y. Pakistan Journal of Biological Sciences, 22: 614-622.
DOI: 10.3923/pjbs.2019.614.622
URL: https://scialert.net/abstract/?doi=pjbs.2019.614.622
DOI: 10.3923/pjbs.2019.614.622
URL: https://scialert.net/abstract/?doi=pjbs.2019.614.622
INTRODUCTION
Potato (Solanum tuberosum L.) is one of the most cultivated crop worldwide1. In Egypt, potato has a significant position among all vegetable crops, where about 20% of the total area devoted for vegetable production is cultivated with potato2. Egypt is the 15th largest potato producer in the world, producing3 4,325,480,00 t in 2017. Therefore, infection of potato with numerous pathogens causes negative effects on the Egyptian quantitative and qualitative potato yield4.
Plant viral diseases represent the second largest plant diseases after plant fungal diseases, leading to crop loss and great damage to agricultural industry5. Potato virus Y (PVY) is one of the most common and dangerous potato viruses that infecting potato worldwide leading to rejection of seed lots from certification programs6,7. Losses from this viral disease reach about 10-80% depending on the variety as well as its effects on the product quality8. Recently, biopesticides have gained more attention and exhibited great development potential. Many researches focused on developing and assessment of new biogenic plant antiviral materials including proteins, polysaccharides and small molecules such as alkaloids, flavonoids, phenols and essential oils from various sources9.
Milk proteins, especially whey proteins, have showed a good potential as antibacterial and antiviral modifiers10,11. Lactoferrin, for instance, has been studied extensively for its antiviral properties12 and its antiviral activity against potato virus X was reported by Taha et al.13. Also esterified whey proteins had antiviral activity against tomato curl viruses14.
Chitosan is a nontoxic biodegradable polysaccharide that possesses various biological activities, including antibacterial, antiviral, antitumor and anticholesterol, therefore it is wide used in medicine and pharmacy15. Recently, researches have focused on the ability of chitosan to increase plant resistance to viruses16,17. It was reported its suppressive effect on many plant viruses18,19.
Curcumin, is a natural compound derived from turmeric (Curcuma longa), widely used as spice and coloring agent in food20 and displays many pharmacological activities21. Recently, curcumin extract was used to control alternaria solani in tomato22. Despite its bioactivities, its application is limited due to its poor solubility in water and limited absorption23. Therefore, many recent attempts were carried out to overcome these problems using different kinds of food-grade polymer materials24-26. It is worth noting that few studies have been carried out on curcumin to control virus populations. Therefore the aim of the present study was to develop and evaluate a new series of curcumin -milk proteins nanoparticles to control the PVY.
MATERIALS AND METHODS
Materials: Sodium caseinate (SC) was obtained from Friesland Campina DMV, Nederland. Whey protein concentrate (WPC 80%) was kindly attained from Onalaska Wisconsin, USA. Whey protein isolate (WPI), α-lactalbumin (α-La 97.46% protein) and β-lactoglobulin (β-lg 97.8% protein) were kindly provided by Davisco Foods International, USA. Chitosan (Cs) of low molecular weight of 100,000-300,000 and sodium tripolyphosphate (TPP) were purchased from Acros Organics, New Jersey, USA. Curcumin (Cur) (>96.1% purity) was obtained from Kolorjet Chemicals Pvt Ltd., Mumbai, Maharashtra, India. Potato tubers cultivar Spunta was obtained from Brown Root project (Ministry of Agr. Egypt). Potato virus (PVY) was obtained from Virus and Phytoplasma Department, Plant Pathology Research Institute, Agricultural Research Center, Giza. ELISA Kits were purchased from LOEWE Biochemica, GmbH, DSMZ (Germany). All other chemicals were of analytical grade. This study was done between December, 2016 to April, 2018.
METHODS
Preparation of chitosan/milk protein nanocomposite and curumin-loaded chitosan/milk proteins nanoparticles (NPs): Nanocomposites of milk proteins-chitosan were prepared by ionotropic gelation method according to Calvo et al.27 with minor modification. Briefly, 1 mL of protein solution (10%) was added to 50 mL chitosan solution (0.2% prepared in 1% acetic acid) under mild stirring. Then, aqueous sodium tripolyphosphate solution (TPP) (0.5 %) was added drop wise to the mixture. The overall mixture was stirred for 30 min at 25°C. The resulting NPs were collected by centrifugation at 25,000 rpm for 30 min at 4°C and washed twice by distilled water and then freeze dried.
About 100 mg of curcumin was dissolved in absolute ethanol till the solution became clear, then added to the previously prepared fresh chitosan/protein nanosuspension and stirred for 30 min at 25°C, followed by centrifugation at 25,000 rpm for 30 min at 4°C. The resulted curcumin-loaded NPs were washed twice with distilled water and then freeze dried and kept at room temperature.
Characterizations of nanoparticles: Particle size, zeta potential, fourier-transform infrared spectroscopy (FTIR) spectra of tested materials also entrapment efficiency and the curcumin release percentage from the nanoparticles were measured as performed by Ali et al.28.
Preparation of virus inocula: Nicotiana (N) tabacum cv. white barley was used as source plants of the virus infections. Virus inocula were prepared by crushing infected leaves with distilled water (1:1 w/v) in a sterilized mortar. Sap extract was squeezed through 2 layers of muslin cloth and the solution was centrifuged at 5000 g for 10 min. (centrifuge2K15,SigmaChemical Co., Osterode, Germany). The clear supernatant was used as virus inoculum.
Antiviral activity: The green house experiments, isolation and detection of the virus were carried out at Virus and Phytoplasma Department, Plant Pathology Research Institute, Agricultural Research Center, Giza from August, 2017 to April, 2018. Potato tubers cultivar Spunta were planted in sterilized soil potting of 30 cm plastic pots (one tuber per pot) under usual and propitious conditions for potato plants. After one month of growth, plants were divided into 14 groups, each group containing 12 plants, 1 of them was healthy (negative control) and others (positive control and treated plants) were mechanically infected with the virus. Three concentrations (500, 1000 and 1500 mg/100 mL) of chitosan, curcumin, native proteins and curcumin loaded milk proteins-chitosan nanoparticles were examined for their antiviral activity. The infected plants were treated by spraying 20 mL of the tested solutions/plant after 7 and 14 days of infections. Two weeks later, leaf samples from each pot were collected and examined by DAS-ELISA and absorption was measured at 405 nm using ELX800 ELISA reader, Bio-TekInstruments, USA. The inhibition (%) was calculated by the formula according to Baranwal and Verma29.
![]()
| C | = | Number of healthy plants |
| T | = | Number of infected plants inoculated by virus mixed with tested solutions |
Preparation of enzyme extract: Leaves of plants inoculated with PVY and sprayed with tested solutions were used to measure the oxidative enzymes activities. About 10 g of fresh plant leaves of each treatment were blended with 20 mL phosphate buffer solution (7.0 pH) and centrifuged at 4000 rpm for 20 min/4°C (centrifuge2K15,SigmaChemical Co., Osterode, Germany). Clear supernatants were used as crude enzymes to determine the activities of peroxidase and polyphenol oxidase according to Anand et al.30.
Determination of peroxidase activity (POX): Peroxidase (POX) activity was assayed spectrophotometrically according to Kar and Mishra31 by measuring the oxidation of pyrogallol to pyrogallin in the presence of H2O2. The reaction mixture consisted of 1.5 mL of 0.05 M pyrogallol, 0.5 mL of enzyme extract and 0.5 mL of 1% H2O2. The reaction mixture was incubated at 25°C for 5min. then the reaction was stopped by adding 0.5 mL of 5% (v/v) H2SO4. The amount of pyrogallin formed was measured at 420 nm.
Determination of polyphenol oxidase activity (PPO): Polyphenol oxidase (PPO) activity was determined following the method of Mayer et al.32. The reaction mixture consisted of 1.5 mL of 0.1 M sodium phosphate buffer (pH 6.5) and 200 μL of the enzyme extract. To start the reaction, 200 μL of 0.01 M catechol was added and the activity was expressed as changes in absorbance at 495 nm.
Serological detection: Enzyme linked immune-sorbent assay (DAS-ELISA) was conducted to test the inhibitory effect of the tested samples according to the protocols described by Clark and Adams33 using Microtitre plate wells coated with the antigen-specific coating-antibody (IgG). About 200 μL/well of (IgG) was diluted in coating buffer at 1:200 and incubated at 37°C for 4 h, according to the supplier’s specifications then remove the reagent from the wells and wash the plates 4 times using the washing buffer (PBST). About 200 μL of the plant sample sap (prepared at a 1:20 dilution in sampling buffer) as well as negative and positive control. The plate was incubated at 4°C overnight. Plates rewashed 4 times as mentioned before. About 200 μL of antibody-AP-conjugate diluted (1:200) from original vial in conjugate buffer were added to each well and incubated at 37°C for 4 h. The plate was washed 4 times. About 200 μL of freshly prepared substrate (1 mg mL–1 4-nitro phenyl phosphate) was dissolved in substrate buffer were added to each well and the plates were incubated at room temperature for 1-2 h. To stop the reaction, 50 μL of 3 M NaOH to each well were added. The alkaline phosphatase (AP) reacts with the substrate 4-nitrophenyl phosphate in an enzymatic reaction, resulting in yellow colored 4-nitrophenol as product. This color formed can be evaluated visually or measured in a spectrophotometer at 405 nm.
Total chlorophyll content: The SPAD-502 chlorophyll meter was used to determine the total chlorophyll in potato leaves according to Ali et al.34. The chlorophyll content was expressed as SPAD value35.
Statistical analysis: A randomized complete block design with 2 factors (factor A: conc. and factor B: treatments) was employed for analysis of chlorophyll content, peroxidase and polyphenol oxidase. The remind parameters used randomized complete block design with one factor with three replications for each parameter. The treatment means were compared by least significant difference (L.S.D.) test as given by Snedecor and Cochran36 using Assistat program, while the standard deviation (SD) was calculated by using excel program.
RESULTS AND DISCUSSION
Characterizations of nanoparticles: The average particle sizes of milk proteins-chitosan nanocomposites ranged from 275.33-334.90 nm. The average particles sizes of casein, α-La and β-Lg nanoparticles (278.10, 290.83 and 274.80 nm, respectively) decreased as a results of loading curcumin while, it increased in case of WPI and WPC nanoparticles (462.80 and 439.90 nm, respectively). All nanoparticles either without or with curcumin are in nano scale. Zeta potential values ranged from -11.13 to +17.30. In case of curcumin loaded milk proteins-chitosan nanoparticles, the zeta potential ranged from -12.63 to +27.73. The FTIR results depict that the NPs were formed due to the interaction between the carboxyl group (-COO-) of protein and amino groups of chitosan. In addition, there was an interaction between chitosan protein NPs and curcumin, they gave the same spectra for curcumin chitosan protein NPs with slight shifting, which confirms that curcumin was loaded in the nanoparticles (data not shown).
The entrapment efficiency (EE %) of curcumin in the suspended protein nanoparticles ranged from 72.27-77.27% (data not shown).
The curcumin release (%) increased gradually in all treatments as the experiment time increased. It reached 96.33-97.83% after 48 h. which indicate that all the used milk proteins can be successfully used as carrier for curcumin (data not shown).
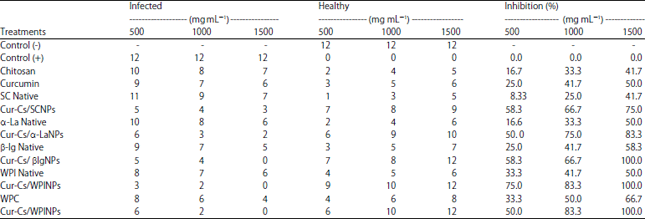
Inhibitory effect (%) of curcumin-loaded milk proteins nanoparticles: Data shown in Fig. 1 depict the external symptoms of PVY of infected plants including leaf crinkle and deformation while in treated plants, the symptoms disappeared to be almost the same as healthy plants. Data in Table 1 show the inhibitory effect (%) of the tested materials estimated by ELISA test. Generally, all the tested materials exhibited antiviral activity against PVY and this effect was concentrations dependent. The antiviral activity of curcumin was more pronounced than chitosan. The native WPC and β-lg showed higher antiviral effect compared to other milk proteins. All of the curcumin loaded nanoparticles form showed higher antiviral activity compared to the native form. It is also observable that the concentration 1500 mg/100 mL led to fully inhibition of the virus especially the nanoparticles of WPC, WPI and β-lg followed by α-la and casein. Also the inhibitory effect of the nanoparticles in most cases is higher than that of curcumin and chitosan alone. The antiviral effect of chitosan can be due to its ability to suppress viral infections by its effect on the plant. Chitosan enhances the plant resistance to infection and induce a broad spectrum of defense responses37. Also it was reported that chitosan inhibited the systemic propagation of viruses and viroids throughout the plant and enhancement of the host’s hypersensitive response to infection38. Moreover, chitosan stimulates the plant immune response39,40. Although, there are many studies on the effect chitosan on plant virus diseases but there are no studies on the effect of curcumin on plant viruses. Many studies dealt with its effect on animal viruses or human viruses. Also it was reported that milk proteins may saturate or interact with the binding of viral DNA or RNA to inhibit viral replication process causing reducing infection of tobacco mosaic virus in pepper, tomato and tobacco14.
| Table 1: | Antiviral activity of native milk proteins and curcumin loaded milk proteins-chitosan nanoparticles |
 | |
 | |
| Fig. 1(a-g): | Symptoms of PVY (a) Infected potato plants, (b) Healthy potato plants, (c) Cur-SCNPs, (d) Cur-α-LaNPs, (e) Cur-β-lgNPs, (f) Cur-WPINPs and (g) Cur-WPCNPs |
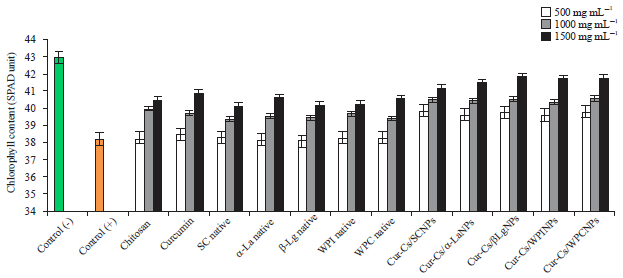 | |
| Fig. 2: | Chlorophyll content (SPAD Unit) in potato plants infected by (PVY) as affected by native milk proteins and its curcumin nanoparticles |
Chlorophyll content: Chlorophyll is a green molecule present in plant cells responsible for greenness of leaves and indicates the nitrogen status in the plant41. Leaf colour gives a good indication of chlorophyll content of leaves42 and used as measure for chloroplast development, photosynthetic capacity, leaf nitrogen content or general plant health43. Chlorophyll content is affected by many factors such as plant genotype, nutrient, leaf thickness or biotic stresses like diseases. The SPAD 502 was designed to detect the chlorophyll content of leaves and used as an indirect indicator of plant N status41.
As shown in Fig. 2 the negative control had high chlorophyll content (42.97), while the chlorophyll content in the positive control was reduced to 38.20. It is observable that both of chitosan and curcumin enhanced the chlorophyll content with non-significant differences and this effect is concentration dependent. Also nanoparticles had higher and significant differences as compared to native form. From these results, it can be concluded that by increasing the concentrations of the tested materials, the content of chlorophyll changed to be almost closed to the negative control readings, which proves the efficacy of tested materials to eliminate the undesirable effects of the virus. The obtained results declare that all curcumin/chitosan/milk proteins nanoparticles enhance the chlorophyll formation in infected plants which is in accordance with Zong et al.44, who reported that chitosan increase the chlorophyll content in Rape in the presence of cadmium.
Oxidative peroxidase (POX) and polyphenol oxidase (PPO): Many defense enzymes are involved in defense reaction against plant pathogens including oxidative peroxidase (POX) and polyphenol oxidase (PPO), which catalyzes the formation of lignin and other oxidative phenols contributing to the formation of defense barriers for reinforcing the cell structure45.
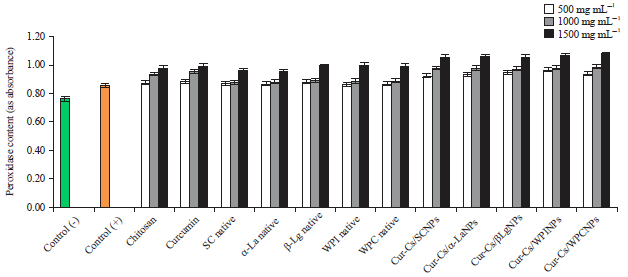 | |
| Fig. 3: | Peroxidase content in potato plants infected by PVY as affected by native milk proteins and its curcumin nanoparticles |
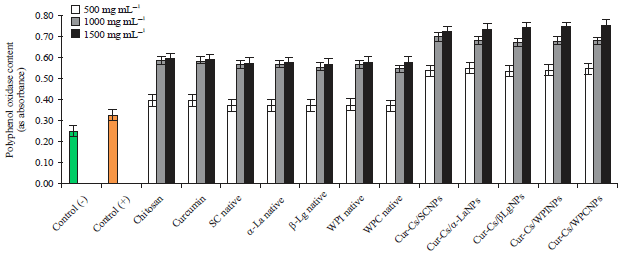 | |
| Fig. 4: | Polyphenol oxidase content in potato plants infected by PVY as affected by native milk proteins and its curcumin nanoparticles |
The antioxidant enzymes PPO and POX are the indication for detoxifying ROS synthesized during stress response46,47. Plant can synthesize and accumulate phenolic compounds in response to stress48. The antioxidant activity of phenolic compounds can play an important role in neutralizing ROS48,49. In agriculture, chitosan is widely used to mimic plant diseases. It has been used as a bio-fungicide, bactericide and virucide, which stimulate the plant defense system against the pathogen and induces the immune system of plants50,51.
Data in Fig. 3 and 4 show POX and PPO activities (as absorbance) in healthy and infected plants. It was 0.760 and 0.250 for healthy plants and 0.850 and 0.320 for infected plants. An increase in enzymatic activity of both enzymes as a result of PVY infection was observed, compared with healthy plants. Non-significant differences were observed between the effect of both of chitosan and curcumin on the oxidative enzymes. It is worth to be mentioned that the values of peroxidase enzyme (POX) and the polyphenol oxidase (PPO) were increased significantly by increasing the concentration of both of native form or the curcumin loaded nanoparticles. It is also observable that all curcumin nanoparticles had significant effect on the determined enzymes as compared to chitosan, curcumin and the native milk proteins.
Chirkov et al.52 reported that treatment of potato plants infected with potato virus X (PVX) with chitosan showed resistance to PVX virus. Also, tomato plants treated with chitosan showed resistance to tomato mosaic virus and improved vegetative growth53. Similarly, chitosan in formulation with plant growth promoting rhizobacteria (PGRP) conferred resistance to leaf curl virus in tomato plant54. Also, chitosan was found effective against squash mosaic virus (SMV)55,56. Furthermore, Sharif et al.51 suggested that this effect might be due to some peculiar properties of the host plant which initiate the antiviral reactions after chitosan treatment. Chandra et al.57 demonstrated that chitosan nanoparticles produce significantly higher defense response in plants compared to natural chitosan and increased activity of the defense enzymes. Although many researches were conducted on the effect of chitosan or chitosan nanoparticle on the oxidative enzymes in plants, no available data were obtained about effect of curcumin or the tested milk proteins on plant viruses.
CONCLUSION
The obtained results revealed that all curcumin-milk proteins nanoparticles at the level of 1500 mg mL–1 especially of WPC, WPI and β-Lg followed by α-La and lastly casein displayed a successful tool to control the potato viral infection (PVY) under greenhouse conditions.
SIGNIFICANCE STATEMENT
This study elucidate the significance of application of nanotechnology in controlling plant virus diseases using promising natural components (milk proteins, chitosan and curcumin). These results will consider as a pioneer data which will help other researchers to continue in this new field.
REFERENCES
- Haase, N.U., 2008. Healthy aspects of potatoes as part of the human diet. Potato Res., 51: 239-258.
CrossRefDirect Link - Scholthof, K.B.G., S. Adkins, H. Czosnek, P. Palukaitis and E. Jacquot et al., 2011. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol., 12: 938-954.
CrossRefDirect Link - Frost, K.E., R.L. Groves and A.O. Charkowski, 2013. Integrated control of potato pathogens through seed potato certification and provision of clean seed potatoes. Plant Dis., 97: 1268-1280.
CrossRefDirect Link - Nolte, P., J.L. Whitworth, M.K. Thornton and C.S. McIntosh, 2004. Effect of seedborne Potato virus Y on performance of Russet Burbank, Russet Norkotah, and Shepody potato. Plant Dis., 88: 248-252.
CrossRefDirect Link - Chatterton, D.E., G. Smithers, P. Roupas and A. Brodkorb, 2006. Bioactivity of β-lactoglobulin and α-lactalbumin-Technological implications for processing. Int. Dairy J., 16: 1229-1240.
CrossRefDirect Link - Kassem, J.M., 2015. Future challenges of whey proteins. Int. J. Dairy Sci., 10: 139-159.
CrossRefDirect Link - Marchetti, M., M.G. Ammendolia and F. Superti, 2009. Glycosaminoglycans are not indispensable for the anti-herpes simplex virus type 2 activity of lactoferrin. Biochimie, 91: 155-159.
CrossRefDirect Link - Taha, S.H., S.A. Mokbel, M. Abdel-Hamid and A.H. Hamed, 2015. Antiviral activity of lactoferrin against potato virus x in vitro and in vivo. Int. J. Dairy Sci., 10: 86-94.
CrossRefDirect Link - Abdelbacki, A.M., S.H. Taha, M.Z. Sitohy, A.I. Abou Dawood, M.M. Abd-El Hamid and A.A. Rezk, 2010. Inhibition of Tomato Yellow Leaf Curl Virus (TYLCV) using whey proteins. Virol. J., Vol. 7.
CrossRefDirect Link - Bykova, V.M. and S.V. Nemtsev, 2002. Syr'evye Istochniki i Sposoby Polucheniya Khitina i Khitozana [Raw Material Sources and Methods of Producing Chitin and Chitosan]. In: Khitin i Khitozan: Poluchenie, Svoistva i Primenenie [Chitin and Chitosan: Production, Properties and Application], Skryabin, K.G., G.A. Vikhoreva and V.P. Varlamov (Eds.). Nauka Publ., Moscow, Russia, ISBN: 5-02-006435-1, pp: 7-23, (In Russian).
- Pospieszny, H. and J.G. Atabekov, 1989. Effect of chitosan on the hypersensitive reaction of bean to alfalfa mosaic virus. Plant Sci., 62: 29-31.
CrossRefDirect Link - Zhao, L., C. Feng, K. Wu, W. Chen, Y. Chen, X. Hao and Y. Wu, 2017. Advances and prospects in biogenic substances against plant virus: A review. Pestic. Biochem. Physiol., 135: 15-26.
CrossRefDirect Link - Jia, Z., D. Shen and W. Xu, 2001. Synthesis and antibacterial activities of quaternary ammonium salt of chitosan. Carbohydr. Res., 333: 1-6.
CrossRefDirect Link - Chirkov, S.N., 2002. The antiviral activity of chitosan (review). Applied Biochem. Microbiol., 38: 1-8.
CrossRefDirect Link - Goel, A., A.B. Kunnumakkara and B.B. Aggarwal, 2008. Curcumin as "Curecumin": From kitchen to clinic. Biochem. Pharmacol., 75: 787-809.
CrossRefPubMedDirect Link - Moghadamtousi, S.Z., H. Abdul Kadir, P. Hassandarvish, H. Tajik, S. Abubakar and K. Zandi, 2014. A review on antibacterial, antiviral and antifungal activity of curcumin. BioMed Res. Int., Vol. 2014.
CrossRefDirect Link - Balbi-Pena, M.I., A. Becker, J.R. Stangarlin, G. Franzener, M.C. Lopes and K.R.F. Schwan-Estrada, 2006. [Control of Alternaria solani in tomato by Curcuma longa extracts and curcumin-I. In vitro evaluation]. Fitopatol. Bras., 31: 310-314, (In Portuguese).
Direct Link - Anand, P., A.B. Kunnumakkara, R.A. Newman and B.B. Aggarwal, 2007. Bioavailability of curcumin: Problems and promises. Mol. Pharm., 4: 807-818.
CrossRefDirect Link - Zhang, Y., Y. Niu, Y. Luo, M. Ge, T. Yang, L.L. Yu and Q. Wang, 2014. Fabrication, characterization and antimicrobial activities of thymol-loaded zein nanoparticles stabilized by sodium caseinate-chitosan hydrochloride double layers. Food Chem., 142: 269-275.
CrossRefDirect Link - Kim, T.H., H.H. Jiang, Y.S. Youn, C.W. Park and K.K. Tak et al., 2011. Preparation and characterization of water-soluble albumin-bound curcumin nanoparticles with improved antitumor activity. Int. J. Pharmaceut., 403: 285-291.
CrossRefDirect Link - Esmaili, M., S.M. Ghaffari, Z. Moosavi-Movahedi, M.S. Atri and A. Sharifizadeh et al., 2011. Beta casein-micelle as a nano vehicle for solubility enhancement of curcumin; food industry application. LWT-Food Sci. Technol., 44: 2166-2172.
CrossRefDirect Link - Calvo, P., C. Remunan-Lopez, J.L. Vila-Jato and M.J. Alonso, 1997. Novel hydrophilic chitosan-polyethylene oxide nanoparticles as protein carriers. J. Applied Polym. Sci., 63: 125-132.
CrossRefDirect Link - Ali, I.H., I.A. Khalil and I.M. El-Sherbiny, 2016. Single-dose electrospun nanoparticles-in-nanofibers wound dressings with enhanced epithelialization, collagen deposition and granulation properties. ACS Applied Mater. Interfaces, 8: 14453-14469.
CrossRefDirect Link - Baranwal, V.K. and H.N. Verma, 1997. Characteristics of a virus inhibitor from the leaf extract of Celosia cristata. Plant Pathol., 46: 523-529.
CrossRefDirect Link - Anand, T., A. Chandrasekaran, S. Kuttalam, T. Raguchander, V. Prakasam and R. Samiyappan, 2007. Association of some plant defense enzyme activities with systemic resistance to early leaf blight and leaf spot induced in tomato plants by azoxystrobin and Pseudomonas fluorescens. J. Plant Interact., 2: 233-244.
CrossRefDirect Link - Kar, M. and D. Mishra, 1976. Catalase, peroxidase and polyphenoloxidase activities during rice leaf senescence. Plant Physiol., 57: 315-319.
CrossRefDirect Link - Mayer, A.M., F. Harel and R. Ben-Shaul, 1966. Assay of catechol oxidase-A critical comparison of methods. Phytochemistry, 5: 783-789.
CrossRefDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Ali, M.M., A. Al-Ani, D. Eamus and D.K.Y. Tan, 2012. A new image processing based technique to determine chlorophyll in plants. Am.-Eurasian J. Agric. Environ. Sci., 12: 1323-1328.
Direct Link - Hassan, O. and T. Chang, 2017. Chitosan for eco-friendly control of plant disease. Asian J. Plant Pathol., 11: 53-70.
CrossRefDirect Link - Badawy, M.E.I. and E.I. Rabea, 2011. A biopolymer chitosan and its derivatives as promising antimicrobial agents against plant pathogens and their applications in crop protection. Int. J. Carbohydr. Chem., Vol. 2011.
CrossRefDirect Link - Iriti, M. and F. Faoro, 2008. Abscisic acid is involved in chitosan-induced resistance to Tobacco Necrosis Virus (TNV). Plant Physiol. Biochem., 46: 1106-1111.
CrossRefDirect Link - Iriti, M. and E.M. Varoni, 2015. Chitosan-induced antiviral activity and innate immunity in plants. Environ. Sci. Pollut. Res. Int., 22: 2935-2944.
CrossRefDirect Link - Borhan, M.S., S. Panigrahi, M.A. Satter and H. Gu, 2017. Evaluation of computer imaging technique for predicting the SPAD readings in potato leaves. Inform. Process. Agric., 4: 275-282.
CrossRefDirect Link - Gaddanakeri, S.A., D.P. Biradar, N.S. Kambar and V.B. Nyamgouda, 2007. Productivity and economics of sugarcane as influenced by leaf colour chart based nitrogen management. Karnataka J. Agric. Sci., 20: 466-468.
Direct Link - Varinderpal-Singh, Bijay-Singh, Yadvinder-Singh, H.S. Thind and R.K. Gupta, 2010. Need based nitrogen management using the chlorophyll meter and leaf colour chart in rice and wheat in South Asia: A review. Nutr. Cycl. Agroecosyst., 88: 361-380.
CrossRefDirect Link - Zong, H., S. Liu, R. Xing, X. Chen and P. Li, 2016. Protective effect of chitosan on photosynthesis and antioxidative defense system in edible rape (Brassica rapa L.) in the presence of cadmium. Ecotoxicol. Environm. Saf., 138: 271-278.
CrossRefDirect Link - Kumar, M., V. Yadav, N. Tuteja and A.K. Johri, 2009. Antioxidant enzyme activities in maize plants colonized with Piriformospora indica. Microbiology, 155: 780-790.
CrossRefDirect Link - Joseph, B. and D. Jini, 2010. Insight into the role of antioxidant enzymes for salt tolerance in plants. Int. J. Bot., 6: 456-464.
CrossRefDirect Link - Naikoo, M.I., M.I. Dar, F. Raghib, H. Jaleel and B. Ahmad et al., 2019. Role and Regulation of Plants Phenolics in Abiotic Stress Tolerance: An Overview. In: Plant Signaling Molecules: Role and Regulation Under Stressful Environments, Khan, M.I.R., P.S. Reddy, A. Ferrante and N.A. Khan (Eds.). Chapter 9, Woodhead Publishing, USA., ISBN: 978-0-12-816451-8, pp: 157-168.
- Kumar, A., P.C. Mali and V.K. Manga, 2010. Changes of some phenolic compounds and enzyme activities on infected pearl millet caused by Sclerospora graminicola. Int. J. Plant Physiol. Biochem., 2: 6-10.
Direct Link - Li, B., S. Shi, C. Shan, Q. Zhou and M. Ibrahim et al., 2013. Effect of chitosan solution on the inhibition of Acidovorax citrulli causing bacterial fruit blotch of watermelon. J. Sci. Food Agric., 93: 1010-1015.
CrossRefPubMedDirect Link - Sharif, R., M. Mujtaba, M.U. Rahman, A. Shalmani and H. Ahmad et al., 2018. The multifunctional role of chitosan in horticultural crops; a review. Molecules, Vol. 23, No. 4.
CrossRefDirect Link - Chirkov, S.N., A.V. Il'ina, N.A. Surgucheva, E.V. Letunova, Y.A. Varitsev, N.Y. Tatarinova and V.P. Varlamov, 2001. Effect of chitosan on systemic viral infection and some defense responses in potato plants. Russian J. Plant Physiol., 48: 774-779.
CrossRefDirect Link - Abd El-Gawad, H.G. and A.M. Bondok, 2015. Response of tomato plants to salicylic acid and chitosan under infection with tomato mosaic virus. Am.-Eurasian J. Agric. Environ. Sci., 15: 1520-1529.
Direct Link - Mishra, S., K.S. Jagadeesh, P.U. Krishnaraj and S. Prem, 2014. Biocontrol of tomato leaf curl virus (ToLCV) in tomato with chitosan supplemented formulations of Pseudomonas sp. under field conditions. Aust. J. Crop Sci., 8: 347-355.
Direct Link - Nagorskaya, V., A. Reunov, L. Lapshina, V. Davydova and I. Yermak, 2014. Effect of chitosan on Tobacco Mosaic Virus (TMV) accumulation, hydrolase activity and morphological abnormalities of the viral particles in leaves of N. tabacum L. cv. Samsun. Virologica Sinica, 29: 250-256.
CrossRefDirect Link - Firmansyah, D., Widodo and S.H. Hidayat, 2017. Chitosan and plant growth promoting rhizobacteria application to control Squash mosaic virus on cucumber plants. Asian J. Plant Pathol., 11: 148-155.
CrossRefDirect Link - Chandra, S., N. Chakraborty, A. Dasgupta, J. Sarkar, K. Panda and K. Acharya, 2015. Chitosan nanoparticles: A positive modulator of innate immune responses in plants. Scient. Rep., Vol. 5.
CrossRefDirect Link