
Theresia Nur Indah Koni
Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
LiveDNA: 62.16921
Rusman
Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Chusnul Hanim
Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Zuprizal
Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 9 | Page No.: 436-441







ABSTRACT
Background and Objective: Bacillus subtilis is a bacterium that can produce the oxalate decarboxylase (ODC) enzyme. This enzyme decomposes oxalate. ODC enzyme activity is influenced by temperature and pH. This study was conducted to determine the optimum pH and temperature for ODC enzyme activity in B. subtilis FNCC 0059. Materials and Methods: B. subtilis was cultivated in oxalate medium at different incubation temperatures for 24 h. The tested temperatures were 25, 29, 33, 37, 41 and 45°C. In separate experiments, cultures were grown in oxalate medium at varying pH from 4-8.5. The cell growth assay and enzyme activity were performed and all the experiments were done with 3 replicates. Data were analyzed using one-way analysis of variance. Results: The optimum temperature of 37°C could produce maximum ODC enzyme activity (0.041 U mL–1) enzyme activity increased to the point of the optimum pH and then decreased 5.6% when the pH was increased 0.5 from the optimum. Maximum ODC enzyme activity (0.0413 U mL–1) was achieved at pH 5.5. Conclusion: Temperature and the pH of the medium affected ODC enzyme activity. Optimum medium condition of B. subtilis was reached at 37°C and pH 5.5.
PDF Abstract XML References Citation
Received: April 07, 2017;
Accepted: October 02, 2017;
Published: October 11, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Theresia Nur Indah Koni, Rusman, Chusnul Hanim and Zuprizal, 2017. Effect of pH and Temperature on Bacillus subtilis FNCC 0059 Oxalate Decarboxylase Activity. Pakistan Journal of Biological Sciences, 20: 436-441.
DOI: 10.3923/pjbs.2017.436.441
URL: https://scialert.net/abstract/?doi=pjbs.2017.436.441
DOI: 10.3923/pjbs.2017.436.441
URL: https://scialert.net/abstract/?doi=pjbs.2017.436.441
INTRODUCTION
Oxalic acid is an antinutrient in plant foodstuffs1-3. It can bind minerals such as calcium and magnesium to form insoluble oxalate. The high capacity of soluble oxalate to bind calcium in blood serum can lead to hypocalcemia, which can stimulate the secretion of parathyroid hormone that causes the kidneys to reabsorb calcium from bones1,3. High oxalate ion concentration in the blood is filtered by the kidneys and it may combine with Ca2+ to form insoluble oxalate crystals that can block urine flow and cause kidney failure3-5. Fifty percent of oxalate comes from the diet and is secreted by the body through urine. Increasing absorption of oxalate in the form of calcium oxalate occurs in 46% of patients with kidney stones6.
Oxalate can be degraded by some lactic acid bacteria, such as Bifidiobacteria5, lactobacillus bacteria6, Enterococcus faecalis7,8, Paracoccus alkaliphilus9, Oxalobacter formigenes10 and B. subtilis11-13. These bacteria produce enzymes that degrade oxalic acid such as decarboxylase and oxalate oxidase enzyme. Oxalic acid (C2H2O4) can be decomposed to formic acid and carbon dioxide by oxalate decarboxylase (ODC) enzymes14. The ODC enzyme activity is influenced by several changes in medium environmental factors medium such as substrate concentration, pH, temperature and presence of inhibitors15. Microbial growth is influenced by physical factors such as pH and temperature. The acidity or alkalinity of the environment affects the activity and stability of enzymes, which can affect microbial growth and metabolism. Culture pH greatly influences enzymatic processes and transport of components in cell membranes; this was investigated by Waleed et al.16. ODC enzymes can be produced by B. subtilis FNCC 0059 and the maximum production can be achieved based on optimum medium conditions. Information related to decarboxylase enzyme activities produced by B. subtilis has not been reported. Therefore, this study was aimed to determine temperature and pH optimum to produce maximum Bacillus subtilis ODC enzyme.
MATERIALS AND METHODS
Microorganisms: This study was conducted from September-December, 2016. The bacteria B. subtilis FNCC 0059 was purchased from the microbiology laboratory of Pusat Antar Universitas, Universitas Gadjah Mada. The strains were stored freeze dried in a refrigerator. The bacteria were cultured in de man rogosa sharpe (MRS) broth and incubated at 37°C for 24 h this as stock culture and 10% of stock culture bacteria were grown in oxalate medium according to Campieri et al.7.
Determination of oxalate: Oxalate was determined using the kinetic spectrophotometric method based on the catalytic effect of the oxalate on rhodamine B oxidation by potassium dichromate in dilute sulfuric acid medium, according to Jiang et al.17. The tube was placed in the water at 90°C, after 15 min the reaction was quenched by cooling it down with the tap water and then the absorbance at 555 nm. The calibration curve was constructed by plotting the difference in absorbance versus oxalate concentration to prepare a calibration graph.
Oxalate decarboxylase enzyme activity assay: Bacillus subtilis has been inoculated in oxalate medium, Campieri et al.7 were harvested and then centrifuged at 10000 rpm for 10 min, the supernatant was used to determine oxalate decarboxylase activity. Oxalate decarboxylase activity was determined by monitoring the change in oxalate using the Jin et al.18 method.
Effect of incubation temperature on enzyme production: The effect of temperature on the production of oxalate decarboxylase was determined by incubating the Bacillus subtilis into the oxalate medium at different temperatures for 24 h according to Campieri et al.7. Temperatures incubated were 25, 29, 33, 37, 41 and 45°C.
Effect of pH on enzyme production: In separate experiments, the effect of pH on ODC enzyme activity was determined by growing B. subtilis in oxalate medium according to Campieri et al.7. pH was adjusted with 2 mol L–1 HCl added to adjust the pH of the medium to 4-5 or with 2 mol L–1 NaOH to adjust the pH of the medium to 6-8, as shown by Tanner and Bonermann19. The culture was incubated for 24 h at 37°C.
Statistical analysis: Data were recorded in triplicate and results are expressed as Mean±SD of 3 replicates. Data were analyzed using one-way analysis of variance (ANOVA). Duncan’s multiple range test was used to separate the means at the 5% level of probability20.
RESULTS
Effect of incubation time on ODC enzyme activity and growth of Bacillus subtilis: Bacterial growth can be seen from the value of optical density (OD). The results show that the activity of oxalate decarboxylase enzyme is maximized when growth is at stationary phase. The lag phase of B. subtilis occurred at the 6th h and then, they entered the log phase at the 12th h and the stationery phase at the 24th h (Fig. 1).
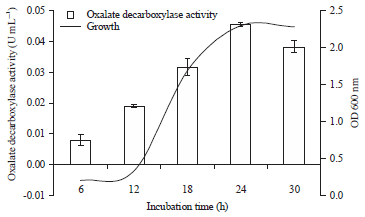 | |
| Fig. 1: | Oxalate decarboxylase activity and growth of Bacillus subtilis at different incubation times |
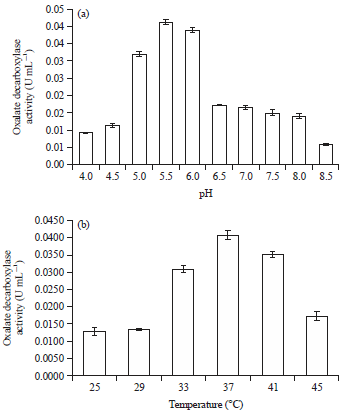 | |
| Fig. 2(a-b): | (a) Effect of pH on the activity of Bacillus subtilis oxalate decarboxylase and (b) Effect of temperature on the activity of Bacillus subtilis oxalate decarboxylase. Mean±SD |
Increasing bacterial growth occurs up to the 24th h and starts to decrease at the 24th h. Maximum absorbance (2.301) is achieved at the 24th h. The maximum growth value was reached at the 24th h incubation, then it is decreased at the 30th h incubation with an absorbance value of 2.283. The best growth conditions for B. subtilis bacteria after 24 h of incubation was used for further experiments.
Effect of pH on ODC enzyme activity: The pH levels of the medium ranged from 4-8.5. The effect of pH on the ODC enzyme activity in B. subtilis is presented in Fig. 2a. ODC activity was increased 4.4 times when the pH increased from 4-5.5. When pH increased from 5.5-6, ODC activity was reduced 0.02 times. The results showed that pH has a significant effect on ODC enzyme activity in B. subtilis. According to Duncan's multiple range tests, it appeared that the pH of 5.5 treatment was higher than other treatments (p<0.05). The highest ODC enzyme activity (0.0413 U mL–1) was detected with pH 5.5 and the lowest (0.0059 U mL–1) was with pH 8.5.
The influence of temperature on ODC enzyme activity: Bacillus subtilis ODC enzyme activities were increased approximately 3.3 times with increasing the temperature up to the optimum temperature on 37°C. Enzyme activity decreased at higher temperatures. These data are presented in Fig. 2b. Oxalate decarboxylase enzyme activity was increased with an incubation temperature of 25-37°C, but it was decreased at temperatures above the optimum temperature (37°C). This occurs because temperature affects the activity of the enzyme in the metabolism of cells and that affects the rate of microbial growth. Bacillus subtilis incubated at 37°C produced oxalate decarboxylase enzyme activity that was significantly higher (p<0.05) compared with the other incubation temperatures. The effect on oxalate decarboxylase enzyme activity of six incubation temperature treatments were significantly different for each treatment (p<0.05).
DISCUSSION
The results of this study found that the growth of B. subtilis FNCC 0059 initially slowed at 6th-12th h incubation, then increased exponentially at 18th incubation, until it peaked at 24th h incubation then remained and decreased at 30th h incubation. The growth of B. subtilis FNCC 0059 follows the general microbial growth as reported by Mahazar et al.21 that three growth phase of microbial consists of lag, log and stationary phase. Lag phase occurs when bacterial inoculum is first introduced into the medium, during the lag phase, the growth rate is close to zero, the cells are active but they are not increasing in cell number22. Bacterial cells start to divide when they acclimate to their environment and this phase is called the log phase. In the log phase, they start regular division by binary fission15. During the log phase, environmental factors such as pH and temperature were important to support bacterial growth19,22. The cell continues to grow until the total number of cells remains constant. This third phase is called the stationary phase.
The highest growth of B. subtilis FNCC 0059 is achieved at 24 h, this is in line with the maximum ODC enzyme activity. This finding is in line with the study of Sasikumar et al.6 which reports the maximum activity of ODC L. plantarum 3050 WCFS and 0373 WCFS1 achieved when growth peaks.
The ODC enzyme activity increased when pH increased up to 5.5, but it decreased by 5.6% when pH increased to 0.5 from the optimum pH (5.5). Low ODC activity at pH 6 might be caused by increasing pH from the optimum, as reported by Kusumadjaja and Dewi 23. They showed that papaia enzyme activity is increased with increasing pH up to 6, but it is decreased at pH 7. Intracellular pH is recognized as one of the most important factors that affect enzyme activities24. The activities of enzymes will increase up to optimum pH and decreased after pH optimum, activities most enzymes have a bell-shaped curve25. Drastic changes in pH can destroy the plasma membrane of bacteria and inhibit enzyme activity and membrane transport proteins22,25. Oxalate decarboxylase (ODC) an optimal at pH of 4.313.
This study was different with Lee et al.12 study that reported oxalate-degrading activity of recombinant B. subtilis PBY was highest at pH 5; B. subtilis YvrK has optimum pH 4.312,19, Anand et al.26, who showed that ODC activity in B. subtilis CU1065 started at pH 3, reached its maximum at pH 5 and disappeared at pH 7.5. Different microbes have different optimum pH as stated by Dutton et al.27 that some of the microbes that produce oxalate decarboxylase have different pH optimum: pH 3 in Collybia velutipes, pH 5.2 in Aspergillus niger and pH 4 in Myrothecium verrucaria. He also showed that Coriolus versicolor at pH 2 had a specific enzyme activity of 0.005 U mg–1. Graz et al.28 reported that oxalate degrading enzymes have optimum activity at low pH values.
In this study, the optimum temperature for ODC enzyme from B. subtilis FNCC 0059 was 37°C. On optimum temperature, the rate of the enzyme activity is high because substrates collide with active sites on the enzyme more frequently as the molecules move rapidly29. Biswanger24 state that enzymes display their highest activity at their respective optimum conditions, deviations from the optimum condition cause a reduction of enzyme activity.
This result matches with Lee et al.12 increased ODC activity at 37°C and Molina et al.13 study that reported the rate of oxidase activity of wild-type B. subtilis is increased in line with increasing temperature, reaching highest activity at 35°C. The optimum temperature in this study was higher than the study by Zhu and Hong30, which reported that the fungus Trametes versicolor was incubated at 25°C with pH of 2-3 to produce the enzyme oxalate decarboxylase activity of 1.2-3.2 U mg–1 protein after 8 days of incubation. While Anbazhagan et al.31 reported L. plantarum produced maximum ODC activity when the temperature at 30°C. This difference is caused by the different microorganisms used in each experiment.
The way enzymes respond to temperature is fundamental to many areas of biology15,32. Enhancement of temperature caused increasing of enzyme activity and at the same time cause enzyme activity to be lost by denaturation32. Prescott et al.15 suggested that the effect of the temperature on microbial growth is due to the sensitivity of enzyme catalyzed reactions. At low temperatures, the growth would accelerate doubly, by increased temperature22. The reaction rate would increase every increasing of temperature 10°C,15,22.
Because of the fast reactions, the metabolism of microbes and microbial growth will increase more rapidly until it reaches the maximum level while exceeding from maximum temperature will cause the death of microbes. This occurs due to denaturation of enzymes, transport carriers and other proteins32. High temperatures also destroy the microbial cell membrane by damaging and decomposing the lipid bilayer coating22.
The present study revealed that the pH 5.5 and temperature 37°C could produce maximum ODC enzyme activity. The results also showed that 24 h incubation would give maximum ODC enzyme activity. The findings of this study are beneficial to researchers and industry who need to produce ODC enzyme from B. subtilis FNCC 0059. Authors believe that the ODC enzyme produced from B. subtilis has great potential for use in the process of fermentation of oxalate-rich foods so that they are safe for consumption by humans or animals.
CONCLUSION
Bacillus subtilis FNCC 0059 was able to produce ODC enzyme required for the degradation of oxalate. The optimum pH was 5.5 and the optimum temperature was 37°C. Bacillus subtilis may be used in the conversion of feed ingredients containing oxalate.
SIGNIFICANCE STATEMENTS
This study discovers optimum medium condition to B. subtilis produce enzyme oxalate decarboxylase. It can be beneficial as a reference in producing enzyme to degradate oxalate in food or feed. This study will help the researcher to uncover the critical areas of the medium condition compatable for B. subtilis produce maximum of enzyme oxalate decarboxylase that many researchers were not able to explore. Thus, a new theory on these pH and temperature of medium compatable for B. subtilis to produce maximum oxalate decarboxylase enzyme may be arrived at in this study.
ACKNOWLEDGMENTS
The authors gratefully appreciate to the Directorate of Higher Education, Ministry of Education The Republic of Indonesia for Doctoral program Scholarship (BPP-DN). Authors also thank to staff of the Biochemical laboratory, Faculty of Animal Science, Universitas Gadjah Mada, for their support in this experiment, Waldi Parera and Tri A.Y. Foenay for their comments on a previous draft of this manuscript.
REFERENCES
- Svedruzic, D., S. Jonsson, C.G. Toyota, L.A. Reinhardt, S. Ricagno, Y. Lindqvist and N.G. Richards, 2005. The enzymes of oxalate metabolism: Unexpected structures and mechanisms. Arch. Biochem. Biophys., 433: 176-192.
CrossRefDirect Link - Rahman, M.M., R.B. Abdullah and W.E.W. Khadijah, 2013. A review of oxalate poisoning in domestic animals: Tolerance and performance aspects. J. Anim. Physiol. Anim. Nutr., 97: 605-614.
CrossRefDirect Link - Giardina, S., C. Scilironi, A. Michelotti, A. Samuele, F. Borella, M. Daglia and F. Marzatico, 2014. In vitro anti-Inflammatory activity of selected oxalate-degrading probiotic bacteria: Potential applications in the prevention and treatment of hyperoxaluria. J. Food Sci., 79: 384-390.
CrossRefDirect Link - Sasikumar, P., S. Gomathi, K. Anbazhagan, A. Abhishek and E. Paul et al., 2014. Recombinant Lactobacillus plantarum expressing and secreting heterologous oxalate decarboxylase prevents renal calcium oxalate stone deposition in experimental rats. J. Biomed. Sci., Vol. 21.
CrossRef - Campieri, C., M. Campieri, V. Bertuzzi, E. Swennen and D. Matteuzzi et al., 2001. Reduction of oxaluria after an oral course of lactic acid bacteria at high concentration. Kidney Int., 60: 1097-1105.
CrossRefDirect Link - Gomathi, S., P. Sasikumar, K. Anbazhagan, S. Sasikumar, M. Kavitha, M.S. Selvi and G.S. Selvam, 2014. Screening of indigenous oxalate degrading lactic acid bacteria from human faeces and South Indian fermented foods: Assessment of probiotic potential. Sci. World J., Vol. 2014.
CrossRef - Anbazhagan, K., C.E. Raja and G.S. Selvam, 2007. Oxalotrophic Paracoccus alcaliphilus isolated from Amorphophallus sp. rhizoplane. World J. Microbiol. Biotechnol., 23: 1529-1535.
CrossRefDirect Link - Turroni, S., B. Vitali, C. Bendazzoli, M. Candela and R. Gotti et al., 2007. Oxalate consumption by lactobacilli: Evaluation of oxalyl-CoA decarboxylase and formyl-CoA transferase activity in Lactobacillus acidophilus. J. Applied Microbiol., 103: 1600-1609.
CrossRefDirect Link - Burrell, M.R., V.J. Just, L. Bowater, S.A. Fairhurst, L. Requena, D.M. Lawson and S. Bornemann, 2007. Oxalate decarboxylase and oxalate oxidase activities can be interchanged with a specificity switch of up to 282 000 by mutating an active site lid. Biochemistry, 46: 12327-12336.
CrossRefDirect Link - Lee, E., B.C. Jeong, Y.H. Park and H.H. Kim, 2014. Expression of the gene encoding oxalate decarboxylase from Bacillus subtilis and characterization of the recombinant enzyme. BMC Res. Notes, Vol. 7.
CrossRef - Molina, L., T. Goodall, U. Twahir, E. Moomaw and A. Angerhofer, 2015. Real-time kinetic studies of Bacillus subtilis oxalate decarboxylase and Ceriporiopsis subvermispora oxalate oxidase using a luminescent oxygen sensor. J. Biochem. Technol., 5: 826-831.
Direct Link - Makela, M., S. Galkin, A. Hatakka and T. Lundell, 2002. Production of organic acids and oxalate decarboxylase in lignin-degrading white rot fungi. Enzyme Microbial Technol., 30: 542-549.
CrossRefDirect Link - Waleed, M., A.A. Faiza and I. Nizar, 2015. Production optimization of pullulanase enzyme produced by Bacillus cereus isolated from Syrian sources. Int. Food Res. J., 22: 1824-1830.
Direct Link - Jiang, Z.L., M.X. Zhao and L.X. Liao, 1996. Catalytic spectrophotometric methods for the determination of oxalic acid. Anal. Chim. Acta, 320: 139-143.
CrossRefDirect Link - Jin, Z.X., C. Wang, W. Chen, X. Chen and X. Li, 2007. Induction of oxalate decarboxylase by oxalate in a newly isolated Pandoraea sp. OXJ-11 and its ability to protect against Sclerotinia sclerotiorum infection. Can. J. Microbiol., 53: 1316-1322.
CrossRefDirect Link - Tanner, A. and S. Bornemann, 2000. Bacillus subtilis YvrK is an acid-induced oxalate decarboxylase. J. Bacteriol., 182: 5271-5273.
Direct Link - Mahazar, N.H., Z. Zakuan, Y. Rukayadi, H. Norhayati and A. Meorhussin, 2017. Optimization of culture medium for the growth of Candida sp. and Blastobotrys sp. as starter culture in fermentation of cocoa beans (Theobroma cacao) using Response Surface Methodology (RSM). Pak. J. Biol. Sci., 20: 154-159.
CrossRefDirect Link - Van Eunen, K. and B.M. Bakker, 2014. The importance and challenges of in vivo-like enzyme kinetics. Perspectives Sci., 1: 126-130.
CrossRefDirect Link - Anand, R., P.C. Dorrestein, C. Kinsland, T.P. Begley and S.E. Ealick, 2002. Structure of oxalate decarboxylase from Bacillus subtilis at 1.75 A resolution. Biochemistry, 41: 7659-7669.
CrossRefDirect Link - Dutton, M.V., M. Kathiara, I.M. Gallagher and C.S. Evans, 1994. Purification and characterization of oxalate decarboxylase from Coriolus versicolor. FEMS Microbiol. Lett., 116: 321-325.
Direct Link - Graz, M., K. Rachwal, R. Zan and A. Jarosz-Wilkolazka, 2016. Oxalic acid degradation by a novel fungal oxalate oxidase from Abortiporus biennis. Acta Biochim. Polonica, 63: 595-600.
Direct Link - Zhu, C.X. and F. Hong, 2010. Induction of an oxalate decarboxylase in the filamentous fungus Trametes versicolor by addition of inorganic acids. Applied Biochem. Biotechnol., 160: 655-664.
CrossRefDirect Link - Anbazhagan, K., P. Sasikumar, S. Gomathi, H.P. Priya and G.S. Selvam, 2013. In vitro degradation of oxalate by recombinant Lactobacillus plantarum expressing heterologous oxalate decarboxylase. J. Applied Microbiol., 115: 880-887.
CrossRefDirect Link - Daniel, R.M., M.E. Peterson, M.J. Danson, N.C. Price and S.M. Kelly et al., 2010. The molecular basis of the effect of temperature on enzyme activity. Biochem. J., 425: 353-360.
CrossRefDirect Link