
N.H. Mahazar
Department of Food Science, Faculty of Food Science and Technology,
Z. Zakuan
Department of Food Science, Faculty of Food Science and Technology,
H. Norhayati
Department of Food Technology, Faculty of Food Science and Technology,
A.S. MeorHussin
Department of Food Technology, Faculty of Food Science and Technology,
Y. Rukayadi
Department of Food Science, Faculty of Food Science and Technology,
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 3 | Page No.: 154-159







ABSTRACT
Background and Objective: Inoculation of starter culture in cocoa bean fermentation produces consistent, predictable and high quality of fermented cocoa beans. It is important to produce healthy inoculum in cocoa bean fermentation for better fermented products. Inoculum could minimize the length of the lag phase in fermentation. The purpose of this study was to optimize the component of culture medium for the maximum cultivation of Candida sp. and Blastobotrys sp. Materials and Methods: Molasses and yeast extract were chosen as medium composition and Response Surface Methodology (RSM) was then employed to optimize the molasses and yeast extract. Results: Maximum growth of Candida sp. (7.63 log CFU mL–1) and Blastobotrys sp. (8.30 log CFU mL–1) were obtained from the fermentation. Optimum culture media for the growth of Candida sp., consist of 10% (w/v) molasses and 2% (w/v) yeast extract, while for Blastobotrys sp., were 1.94% (w/v) molasses and 2% (w/v) yeast extract. Conclusion: This study shows that culture medium consists of molasses and yeast extract were able to produce maximum growth of Candida sp. and Blastobotrys sp., as a starter culture for cocoa bean fermentation.
PDF Abstract XML References Citation
Received: October 08, 2016;
Accepted: January 13, 2017;
Published: February 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
N.H. Mahazar, Z. Zakuan, H. Norhayati, A.S. MeorHussin and Y. Rukayadi, 2017. Optimization of Culture Medium for the Growth of Candida sp. and Blastobotrys sp. as Starter Culture in Fermentation of Cocoa Beans (Theobroma cacao) Using Response Surface Methodology (RSM). Pakistan Journal of Biological Sciences, 20: 154-159.
DOI: 10.3923/pjbs.2017.154.159
URL: https://scialert.net/abstract/?doi=pjbs.2017.154.159
DOI: 10.3923/pjbs.2017.154.159
URL: https://scialert.net/abstract/?doi=pjbs.2017.154.159
INTRODUCTION
Traditionally, fermentation technologies used molasses for ethanol and animal feed production1. Molasses is a byproduct from sugar refining. It is thick and dark syrup resulting from crystallization and removal of sucrose from original juice2. Molasses contain approximately 50% (w/w) of fermentable sugars including glucose, sucrose and fructose. It has been widely applied in food and non-food industries due to high nitrogen and sugar content. Other than that, molasses was used as a substrate for yeast production as it is highly available, low cost and does not contain substance that can inhibit fermentation3,4.
Molasses as described is a source of carbon essential for the growth of microorgansims. Other than that, nitrogen which is essential for the growth of yeast is one of the main elements playing important role in structure and function that can be found in the cells5. Yeasts obtained nitrogen source from a wide variety of compounds. Yeast extract was made by extracting the cell content by exogenous or endogenous enzyme. Yeast extract are rich in amino acids, vitamins, peptides and growth simulating compounds. It is commonly used as food additives, flavoring and important component in the medium as supplement for the growth of microorganisms3,6,7. Therefore, addition of yeast extract into the culture medium would be able to promote the growth of yeast for inoculum development in cocoa bean fermentation.
Inoculum is important factor in fermentation in order to achieve optimal production of microorganisms. Healthy inoculum would be able to minimize the length of the lag phase in fermentation8. This could be beneficial in fermentation of cocoa beans. Use of starter culture in cocoa bean fermentation was suggested to control the fermentation process producing consistent, predictable and high quality of fermented cocoa9. Thus, the aim of this study was to optimize culture medium for the maximum production of Candida sp. and Blastobotrys sp. The cells obtained were used as starter cultures in fermentation of cocoa beans.
MATERIALS AND METHODS
Yeast cultures and maintenance: Candida sp. and Blastobotrys sp., used throughout this study were provided by Barry Callebaut Malaysia Sdn. Bhd, Kuala Lipis, Pahang. Stock cultures of this strain were prepared in 1 mL aliquots by adding 0.5 mL of overnight culture with an appropriate media to 0.5 mL of 20% (w/v) glycerol. The cultures were mix thoroughly with glycerol10 by vortex and kept at -80°C. The cultures were maintained on Yeast Peptone Dextrose (YPD) agar containing 1% yeast extract (Oxoid, Basingstoke, UK), 2% peptone (Oxoid, Hampshire, England), 2% glucose (Merck, Darmstadt, Germany) and 2% agar (Oxoid, Hampshire, England).
Inoculum development and growth curve of yeasts: Inoculum for optimization of culture media was prepared by growing the culture in YPD broth. One to two loops of the culture colony from the YPD agar was inoculated into 10 mL of YPD broth by means of an inoculating loop. Incubation was carried out with constant shaking at 180 rpm at 30°C for 24 h in orbital shaker (Jeio-Tech orbital shaking incubator, model SI-600 R, Seoul, Korea). Growth curve of yeasts was determined by inoculation of 1% (v/v) culture into 100 mL of YPD broth. The cultures were shaken using rotary shaker incubator at 30°C with 180 rpm. At 0, 2, 4, 8 and every 4 h until reached 60 h, sampling was done by pipetting out of 10 mL culture from each flask for optical density and cell count analysis. About 2 mL aliquot of culture was transferred into cuvette. The absorbance was read at 600 nm. The cell count was performed by serially diluted 0.1 mL of culture into 0.9 mL Phosphate Buffer Solution (PBS). About 0.1 mL from each dilution was spread onto YPD agar. The plates were then incubated at 30°C for 48 h and the numbers of colonies were counted. The analysis was done three times with three replicates (n = 3×3).
Optimization of Molasses Yeast Extract (MYE) media: Yeast extract (Merck, Darmstadt, Germany) and molasses (Liqueur Agency Sdn. Bhd., Malaysia) were used in fermentation media preparation. The RSM was used to determine optimum yeast growth affected by two factors (molasses and yeast extract). A 22 Central Composite Design (CCD) was employed in this study. A total of 13 runs containing different levels of molasses and yeast extract are shown in Table 1. Fermentation media containing molasses and yeast extract with 100 mL working volume were prepared in 500 mL conical flask.
| Table 1: | Experimental design of Response Surface Methodology (RSM) |
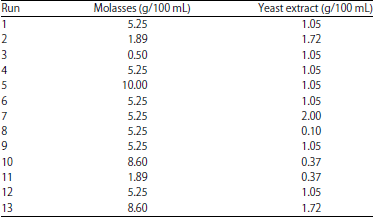 | |
Flasks were plugged with cotton stoppers and sterilized at 121°C for 15 min. Fresh prepared inoculum was added 1% (v/v) aseptically into the fermentation. The runs were conducted for 32 and 36 h in a temperature controlled orbital shaker at 180 rpm, which enabled adequate homogenization for better contact between substrates and the yeast culture11. At the end of fermentation, 1 mL samples were aseptically taken out from the media for determination of cell counts. About 0.1 mL sample was placed into an eppendorf tube containing 0.9 mL PBS solution and mixed thoroughly using vortex. This study suspension was designated as 10–1. About 0.1 mL of diluted samples was transferred into another tube containing 0.9 mL sterile PBS and thoroughly mixed. The dilution was made sequentially from tube to tube to increase diluted cell suspensions. About 0.1 mL from each dilution was pipetted and transferred onto YPD agar plate12. Inverted plates were incubated at 30°C for 48 h and viable colonies grown on agar plates were counted. The analysis was done 3 times with 3 replicates (n = 3×3).
Statistical analysis: Minitab software (Version 14.0, Minitab Co., PA, USA) was used for statistical analysis. Analysis of variance (ANOVA) was carried out for its statistical significance. The p<0.05 was considered as significant.
RESULTS
Inoculum development and growth curve of yeasts: Determination of growth curve requires inoculation of viable cells into broth medium and incubated at optimum conditions. Under these conditions, the cells produce rapidly and dynamic of growth charted by plotting number of cells versus incubation time. In this study, yeasts (Candida sp. and Blastobotrys sp.) were inoculated separately into enrich medium, YPD broth to determine the growth curve of these yeasts. Time of incubation period which gave the maximum number of yeast cells was used for harvesting the cell in optimization of molasses-yeast extract media for the cultivation of Blastobotrys sp. and Candida sp. Result shows Blastobotrys sp., lag phase was approximately 6 h before enter the log phase (Fig. 1). The log phase of Blastobotrys sp., was from 12 h until 32 h. Blastobotrys cell number increased to maximum number from 105 CFU mL–1 until 108 CFU mL–1 after incubated for 32 h. From this experiment, Blastobotrys sp., achieved maximum number of cells after 32 h of incubation after no further increased was observed in yeast cells over incubation time. Therefore, the time of incubation was used for harvesting the yeast cell in next experiment. Meanwhile, Candida sp., showed lag phase occurred less than 10 h (Fig. 2). Growth of Candida sp., started to increase from 104-108 CFU mL–1. Candida sp., achieved maximum number of cells after incubation of 36 h. From this experiment, Blastobotrys sp. and Candida sp., achieved 8.28 and 7.55 log CFU mL–1 after 32 and 36 h of incubation, respectively. Thus, these times of incubation were used for next experiment.
Optimization of Molasses Yeast Extract (MYE) media: In this study, high production of Candida sp. and Blastobotrys sp., were evaluated in 13 different concentration of medium component using RSM.
| Table 2: | Analysis of variance (ANOVA) for the fitted regression model of Candida sp., production |
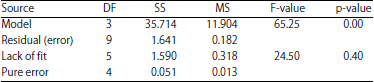 | |
| DF: Degree of freedom, SS: Sum of square, MS: Mean of square | |
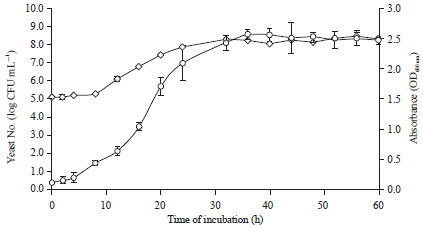 | |
| Fig. 1: | Blastobotrys sp., growth in YPD medium. Right axis: ◯: Absorbance at 600 nm. Left axis: ◇: Yeast number (log CFU mL–1) |
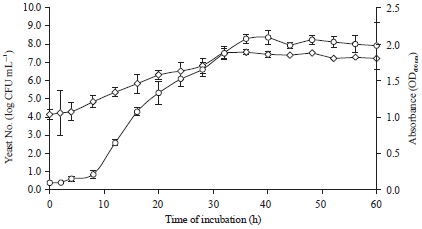 | |
| Fig. 2: | Candida sp. growth in YPD medium. Right axis: ◯: Absorbance at 600 nm. Left axis: ◇: Yeast number (log CFU mL–1) |
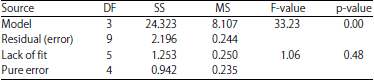
| Table 3: | Analysis of variance (ANOVA) for the fitted regression model of Blastobotrys sp., production |
 | |
| DF: Degree of freedom, SS: Sum of square, MS: Mean of square | |
| Table 4: | Validated yeast concentration of optimized molasses-yeast extract |
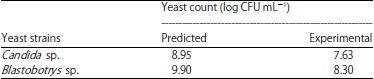 | |
Table 2 and 3 show the results on analysis of variance (ANOVA) to indicate the adequacy of fitted model. The p<0.05 indicates that the equation obtained was appropriate and suitable after model reduction. The experimental results for both of Candida sp. and Blastobotrys sp., were fitted in the second order polynomial model as follow:
| (1) |
| (2) |
where, Y is the response (i.e., Candida sp. and Blastobotrys sp., yield), x1 is molasses and x2 is yeast extract.
The adequacy for 2 regression models (Candida sp. and Blastobotrys sp.) production were determined by (R2 = 0.956 and 0.917), which means 95.6 and 91.7% are the response variability of the model. The adjusted R2 (0.941 and 0.889) further supported the model accuracy. Based on the regression model a maximum production of Candida sp., was predicted to produce log 8 CFU mL–1 using 10 g/100 mL molasses and 2 g/100 mL yeast extract. Blastobotrys sp., production was predicted to give log 9 CFU mL–1 using 1.92 g/100 mL molasses and 2 g/100 mL yeast extract. Validation of the predicted models was done experimentally based on optimum concentration of independent variables. Under optimized culture medium, Candida sp. and Blastobotrys sp., produced 7.63 and 8.30 log CFU mL–1 (Table 4). The good correlation between predicted and experimental values verifies the validity of the response models. Molasses seem to affect the production of yeasts.
DISCUSSION
Microbial growth curve has sigmoid curve consist of lag, log and stationary phase. Lag phase referred as adaptation time with growth rate is close to zero. The cells are biochemically active but not increase in cell number. During this period, metabolic pathway of the cells is initiated prior to start the cell division. Length of lag phase varies might be because of initial inoculum size and environmental factor such as nutrient, incubation temperature and pH of culture medium13. When cells are adapted, they started to divide. This phase called as exponential or log phase14. Exponential or log phase characterized by high energy consumption with constant growth rate15. The cells continue to grow until the total number of viable cell constant in stationary phase12,13. During exponential growth phase, yeast cells used sugar via glycolysis produced ethanol and acetic acid into the medium. As sugar depleted, yeast cells start to switch from consumption of sugar to ethanol until completely utilized16. Kusch et al.15 reported Candida albican made energy from glucose in medium through substrate phosphorylation or oxidative phosphorylation. In a medium with rich glucose medium based, the mitochondrial structure were disrupted but it back to normal when aerobic metabolism replaces alcoholic fermentation17. When the cells lack of essential nutrient, they enter resting stage known as stationary phase18. Stationary phase occurred when the cells stop to divide19. In a study by Abadias et al.8, maximum growth of Candida sake was observed after 30 h grown in molasses medium. Maximum number achieved in short period was due to agitation-aeration combination in fermenter. The best agitation speed allows the oxygen distributed well in the culture medium to be used by yeast cells for respiration.
The MYE medium consist of molasses as carbon source. Molasses contain fermentable sugar compose of fructose, glucose and sucrose. The membrane of yeast cell was not freely permeable for all sugar20. The sugars were transported into the cell through protein transport embedded in the cell membrane either in their intact form (glucose and fructose) or they were firstly hydrolysed to monosaccharide components21. Hydrolysis of disaccharide sugar depends on the yeast strain and the nature of sugar may occur inside or outside the cell membrane. In most cases, hydrolysis of sucrose occurred outside the cell membrane by the action of enzyme invertase into glucose and fructose22,23. Molasses was found to be the most preferred carbon source compared to glucose, sucrose and rice water by marine yeasts (Debaryomyces hansenii, Debaryomyces hansenii, Candida sake and Candida tropicalis) in a study by Sarlin and Philip24. Molasses are rich in nutrients mostly required by the microorganism, such as nitrogen, sodium, phosphorus and potassium25. Optimal sugar concentration for the yeast growth was found to be 17.5% (w/v) of commercial sucrose and average 8.75% (w/v) of cane molasses26. Different in concentrations between these 2 carbon sources probably because of the impurities in cane molasses such as SO2, clorets and Al2O3. High impurities concentration may inhibit the yeast growth26. Sarlin and Philip24 stated that molasses served as a good carbon source with addition of growth promoting components which one of it found to be amino acid27.
CONCLUSION
This study demonstrated that cultivation of Candida sp. and Blastobotrys sp., could be produced in liquid medium. Maximum growth of these 2 yeasts was produced in medium consist of molasses and yeast extract. Under optimized culture media, yeasts were able to produce about 7.63 log CFU mL–1 of Candida sp. and 8.30 log CFU mL–1 of Blastobotrys sp. The optimum concentration of medium compositions for Candida sp., was 10% (w/v) molasses and 2% (w/v) yeast extract, while for Blastobotrys sp., was 1.94% (w/v) molasses and 2% (w/v) yeast extract, respectively.
ACKNOWLEDGMENTS
This study was supported by Grant Universiti Putra Malaysia Yaya Rukayadi under the project No. GP-IPS/2014/9446800 and Barry Callebaut Services, Asia Pacific, Sdn. Bhd. Malaysia. Thank you very much to all those who have helped the completion of the study and writing of this manuscript.
REFERENCES
- Sarka, E., Z. Bubnik, A. Hinkova, J. Gebler and P. Kadlec, 2012. Molasses as a by-product of sugar crystallization and a perspective raw material. Procedia Eng., 42: 1219-1228.
CrossRefDirect Link - Xu, W., L. Liang and M. Zhu, 2015. Determination of sugars in molasses by HPLC following solid-phase extraction. Int. J. Food Propert., 18: 547-557.
CrossRefDirect Link - Bekatorou, A., C. Psarianos and A.A. Koutinas, 2006. Production of food grade yeasts. Food Technol. Biotechnol., 44: 407-415.
Direct Link - Skountzou, P., M. Soupioni, A. Bekatorou, M. Kanellaki, A.A. Koutinas, R. Marchant and I.M. Banat, 2003. Lead(II) uptake during baker's yeast production by aerobic fermentation of molasses. Process Biochem., 38: 1479-1482.
CrossRefDirect Link - Da Cruz, S.H., E.M. Cilli and J.R. Ernandes, 2002. Structural complexity of the nitrogen source and influence on yeast growth and fermentation. J. Inst. Brewing, 108: 54-61.
CrossRefDirect Link - Hakobyan, L., L. Gabrielyan and A. Trchounian, 2012. Yeast extract as an effective nitrogen source stimulating cell growth and enhancing hydrogen photoproduction by Rhodobacter sphaeroides strains from mineral springs. Int. J. Hydrogen Energy, 37: 6519-6526.
CrossRefDirect Link - Sayed, E.T., N.A.M. Barakat, M.A. Abdelkareem, H. Fouad and N. Nakagawa, 2015. Yeast extract as an effective and safe mediator for the Baker's-yeast-based microbial fuel cell. Ind. Eng. Chem. Res., 54: 3116-3122.
CrossRefDirect Link - Abadias, M., N. Teixido, J. Usall and I. Vinas, 2003. Optimization of growth conditions of the postharvest biocontrol agent Candida sake CPA-1 in a lab-scale fermenter. J. Applied Microbiol., 95: 301-309.
CrossRefDirect Link - Lefeber, T., Z. Papalexandratou, W. Gobert, N. Camu and L. De Vuyst, 2012. On-farm implementation of a starter culture for improved cocoa bean fermentation and its influence on the flavour of chocolates produced thereof. Food Microbiol., 30: 379-392.
CrossRefDirect Link - Anane, E., E. van Rensburg and J.F. Gorgens, 2013. Optimisation and scale-up of α-glucuronidase production by recombinant Saccharomyces cerevisiae in aerobic fed-batch culture with constant growth rate. Biochem. Eng. J., 81: 1-7.
CrossRefDirect Link - Yonten, V. and N. Aktas, 2014. Exploring the optimum conditions for maximizing the microbial growth of Candida intermedia by response surface methodology. Preparat. Biochem. Biotechnol., 44: 26-39.
CrossRefDirect Link - Ibarz, A. and P.E.D. Augusto, 2015. An autocatalytic kinetic model for describing microbial growth during fermentation. Bioprocess Biosyst. Eng., 38: 199-205.
CrossRefDirect Link - Kusch, H., S. Engelmann, R. Bode, D. Albrecht, J. Morschhauser and M. Hecker, 2008. A proteomic view of Candida albicans yeast cell metabolism in exponential and stationary growth phases. Int. J. Med. Microbiol., 298: 291-318.
CrossRefDirect Link - Rezaei, M.N., E. Dornez, P. Jacobs, A. Parsi, K.J. Verstrepen and C.M. Courtin, 2014. Harvesting yeast (Saccharomyces cerevisiae) at different physiological phases significantly affects its functionality in bread dough fermentation. Food Microbiol., 39: 108-115.
CrossRefDirect Link - Zakrajsek, T., P. Raspor and P. Jamnik, 2011. Saccharomyces cerevisiae in the stationary phase as a model organism-characterization at cellular and proteome level. J. Proteomics, 74: 2837-2845.
CrossRefDirect Link - Herman, P.K., 2002. Stationary phase in yeast. Curr. Opin. Microbiol., 5: 602-607.
CrossRefDirect Link - Lester, R.L., B.R. Withers, M.A. Schultz and R.C. Dickson, 2013. Iron, glucose and intrinsic factors alter sphingolipid composition as yeast cells enter stationary phase. Biochimica Biophysica Acta (BBA)-Mol. Cell Biol. Lipids, 1831: 726-736.
CrossRefDirect Link - Horak, J., 2013. Regulations of sugar transporters: insights from yeast. Curr. Genet., 59: 1-31.
CrossRefDirect Link - Flores, C.L., C. Rodriguez, T. Petit and C. Gancedo, 2000. Carbohydrate and energy-yielding metabolism in non-conventional yeasts. FEMS Microbiol. Rev., 24: 507-529.
CrossRefPubMedDirect Link - D'Amore, T., I. Russell and G.G. Stewart, 1989. Sugar utilization by yeast during fermentation. J. Ind. Microbiol. Biotechnol., 4: 315-323.
CrossRefDirect Link - Garjonyte, R., V. Melvydas and A. Malinauskas, 2016. Mediated amperometry reveals different modes of yeast responses to sugars. Bioelectrochemistry, 107: 45-49.
CrossRefDirect Link - Sarlin, P.J. and R. Philip, 2013. A molasses based fermentation medium for marine yeast biomass production. Int. J. Res. Mar. Sci., 2: 39-44.
Direct Link - El-Gendy, N.S., H.R. Madian and S.S.A. Amr, 2013. Design and optimization of a process for sugarcane molasses fermentation by Saccharomyces cerevisiae using response surface methodology. Int. J. Microbiol., Vol. 2013.
CrossRefDirect Link - Dorta, C., R. Cruz, P. de Oliva-Neto and D.J.C. Moura, 2006. Sugarcane molasses and yeast powder used in the fructooligosaccharides production by Aspergillus japonicus-FCL 119T and Aspergillus niger ATCC 20611. J. Ind. Microbiol. Biotechnol., 33: 1003-1009.
CrossRefDirect Link - Nakata, H., M. Tamura, T. Shintani and K. Gomi, 2014. Evaluation of baker's yeast strains exhibiting significant growth on Japanese beet molasses and compound analysis of the molasses types. J. Biosci. Bioeng., 117: 715-719.
CrossRefDirect Link