
Noor Aini Habibah
Faculty of Biology, Universitas Gadjah Mada, 55281 Yogyakarta, Indonesia
LiveDNA: 62.14305
Sukarti Moeljopawiro
Laboratory of Biochemistry, Faculty of Biology, Universitas Gadjah Mada, 55281 Yogyakarta, Indonesia
LiveDNA: 62.14307
Kumala Dewi
Laboratory of Plant Physiology, Faculty of Biology, Universitas Gadjah Mada, 55281 Yogyakarta, Indonesia
LiveDNA: 62.14003
Ari Indrianto
Laboratory of Biotechnology, Faculty of Biology, Universitas Gadjah Mada, 55281 Yogyakarta, Indonesia
LiveDNA: 62.14320
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 4 | Page No.: 197-203







ABSTRACT
Background and Objective: Stelechocarpus burahol is a plant containing flavonoid compounds that have the potential for use as an antihyperuricemic for gout medication. This study was performed to assess flavonoid production, growth and cell differentiation of S. burahol in cell suspension culture. Methodology: Mesocarp was planted in Murashige and Skoog (MS) medium supplemented with 7.5 mg L–1 picloram for the induction of callus. Non-embryonic callus obtained was used in the formation of cell suspension cultures. Growth of cells was determined by fresh and dry weights. During the culturing, the fresh weight, dry weight and flavonoid content were determined as a result of culture status. Results: The growth of the S. burahol cell suspension was slow, the stationary phase occurred at 30 days. The production of flavonoids was not in line with the growth of cells and the maximum production occurred on the 15th day of the log phase. The globular-shaped cells dominated the cell suspension culture at all ages. Fluorescein diacetate (FDA) staining of cells derived from cell cultures aged for 36 days showed that some cells were still viable. Conclusion: The results show that flavonoid production, growth and cell differentiation of a S. burahol cell suspension culture differed according to the culture age.
PDF Abstract XML References Citation
Received: January 12, 2017;
Accepted: February 23, 2017;
Published: March 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Noor Aini Habibah, Sukarti Moeljopawiro, Kumala Dewi and Ari Indrianto, 2017. Flavonoid Production, Growth and Differentiation of Stelechocarpus burahol (Bl.) Hook. F. and Th. Cell Suspension Culture. Pakistan Journal of Biological Sciences, 20: 197-203.
DOI: 10.3923/pjbs.2017.197.203
URL: https://scialert.net/abstract/?doi=pjbs.2017.197.203
DOI: 10.3923/pjbs.2017.197.203
URL: https://scialert.net/abstract/?doi=pjbs.2017.197.203
INTRODUCTION
Stelechocarpus burahol contains flavonoid compounds that have the potential for use as an antihyperuricemic for gout medication. This potential has been investigated by Purwantiningsih et al.1, the results of in vivo tests on both the ethanol extract and hexane extract of S. burahol show a potential for use in lowering blood uric acid levels. The S. burahol also contains antioxidants in its leaves, bark, flowers and fruit2. One of the flavonoid compounds in S. burahol is 3, 7, 3’, 4’ tetrahydroxy 5 methyl flavone, which is the most active antioxidant3.
Biotechnological approaches, particularly plant tissue cultures are found to have significant potential for the production of high-value phytochemicals. Plant tissue methods allow selection independent of season and the rapid and effective production of secondary metabolites4. Synthetic compounds of in vitro cultures have been studies and reported frequently4-6. Moreover, the study regarding particular plants has reported that in vitro cultures contain secondary metabolites at higher levels than the original plant7-9. Three types of new flavonoid have been revealed and identified from the culture of Andrographis paniculata callus10.
Cell cultures are composed of homogeneous cells. However, in several studies, it has been reported that some cell suspension cultures consist of cells that undergo differentiation. For example, Cupressus macrocarpa showed oblong cells in the 2nd week of culture; then, in the next 2 weeks, the shape of the cells were transformed into ellipse11. Moreover, a C. arizonica cell suspension culture consistently showed globular-shaped cells, indicating that C. arizonica maintained the globular-shaped cells until the end of the growth period11. Another example is a Rollinia mucosa culture grown in a medium supplemented with picloram. The results of all the concentrations tested showed that culture in picloram was more homogeneous than culture in a medium supplemented with NAA or 2,4-D. However, the culture suspension of Rollinia mucosa still exhibited differentiated cells. At 20-32 days, the differentiated cells were found at their highest amount12. This research studies the flavonoid production, growth and cell differentiation of S. burahol in a cell suspension culture.
MATERIALS AND METHODS
The fruit explant was sterilized using running water and the outer shell was discarded, followed by immersion in liquid detergent (1 mL in 50 mL of distilled water) for 15 min. It was soaked in 5.25% NaClO solution for 30 min and rinsed with sterile water. Sterilization was continued in the laminar cabinet by soaking in 5.25% NaClO solution for 10 min and washing 3 times with sterile distilled water. Mesocarp was separated from the other parts using a scalpel and tweezers. The sterile explants were cut to ±2×1 cm. The explants were planted in a MS medium supplemented with 7.5 mg L–1 picloram. The explant was put on the shelf at an incubation temperature of 24-25°C in dark conditions. Culture bottles were maintained for approximately 3 months. Non-embryonic callus obtained was used in the formation of cell suspension cultures. Suspension cultures were initiated from 3 months old callus by transferring 2 g into a 100 mL Erlenmeyer containing 20 mL of MS medium. The cell culture was shaken on a rotary shaker at 100 rpm for 3-4 weeks13. An Erlenmeyer flask containing 50 mL of medium plus 2.5 g of cells was used for treatments. Cell growth was determined by the fresh and dry weights every 3 days for 36 days. Changes in the shape of the cells to determine the degree of differentiation were determined by distinguishing between globular-shaped (<50 μm), globular (>50 μm) and elongated cells.
The growth curve determination was performed by harvesting the cells every 3 days for 36 days. The cells were placed in the filter and were put in the oven for 48 h at 60°C. The dry cells were weighed and the flavonoid content was analyzed. Cell differentiation was also performed every 3 days for 36 days.
Flavonoid extraction was performed by the procedure detailed by Hao et al.14 with modification. Cells were dried (0.2 g) and ground into a powder using a mortar and pestle. The cells powder was extracted using 5 mL methanol containing 1% (v/v) HCl, followed by the addition 5 mL of 2 N HCl and incubation for 1 h at 90°C. The extract was then dried and re-suspended in methanol and its flavonoid content was analyzed. Total flavonoid content was determined following the procedure specified by Zou et al.15. The 0.5 mL aliquot of the appropriately diluted sample solution was mixed with 2 mL of distilled water and was subsequently added to 0.15 mL of 5% NaNO2 solution. After 6 min, 0.15 mL of 10% AlCl3 solution was added and allowed to stand for 6 min. Two milliliters of 4% NaOH solution was added to the mixture. Immediately, water was added until the final volume of 5 mL was achieved; the mixture was thoroughly mixed and allowed to stand for another 15 min. The absorbance of the mixture was determined at 510 nm. Quercetin was employed as the standard.
RESULTS
The production of secondary metabolites through in vitro culture was reported using cell suspension cultures. Cell culture was obtained by maintaining calluses in a liquid medium on a rotary shaker. The results showed that callus formation from S. burahol mesocarp explants was initiated on 22nd day after planting (Fig. 1).
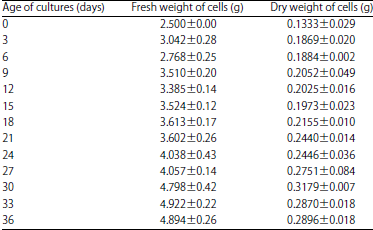
Growth of the cell suspension culture: The growth of the cell suspension culture as indicated by the fresh and dry weights of the cells. In Table 1, the growth of S. burahol was maximized between 30-36 days (the stationary phase of cell suspension culture). Based on the dry weight, the highest growth in the cell culture occurred on the 30th day (0.3179±0.007) of cell culture.
 | |
| Fig. 1: | Callus from mesocarp of S. burahol on MS medium supplemented with 7.5 mg L–1 picloram after of 2 months period for culturing |
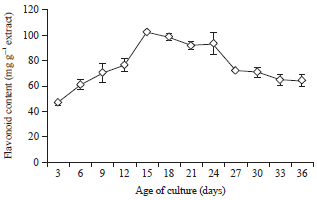 | |
| Fig. 2: | Flavonoid content of S. burahol cell culture in MS medium supplemented with 7.5 mg L–1 picloram in all growth phases |
The lag phase of the cell suspension cultures occurred from 0-6 days. It was followed by the log phase at 6-30 days and the cells then reached the stationary phase.
Flavonoid content: The flavonoid content of the cell suspension culture is presented in Fig. 2. The highest flavonoid content was obtained on the 15th day of growth (Fig. 2). The maximum production of flavonoids in the S. burahol cell suspension culture was produced during the cell growth in the log phase. At the stationary phase (30 days), the production of flavonoids was decreased.
Cell differentiation: The shape of cells changed in the cell suspension cultures, as shown in the cell differentiation process. Therefore, the cell culture contains various forms of cells with a different degree of differentiation. The FDA staining of cells derived from cell cultures aged 36 days showed that some cells were still viable (Fig. 3). The observation also revealed that the viable cells were globular and formed cell aggregates, whereas, the cells that were undergoing the differentiation were found to be elongated and indicated the non-viable conditions.
The level of cell differentiation in the cell suspension culture was differentiated into globular (<50 μm), globular (>50 μm) and elongated (Fig. 4). The observation of S. burahol cell culture showed that the cell composition was different based on the level of cell differentiation (Fig. 5). The globular-shaped cells with size <50 μm dominated the cell suspension cultures at all ages (Fig. 5), while the elongated-shaped cells were the least frequent. The highest number of elongated-shaped cells was found to be concentrated at 33 days of culture and the lowest amount was found at the age of 36 days.
| Table 1: | Growth of cell cultures of S. burahol mesocarp explants in MS medium with 7.5 mg L–1 picloram |
 | |
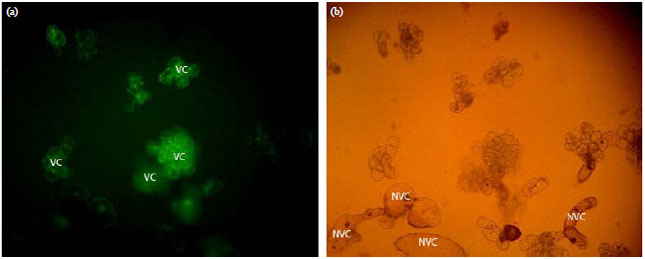 | |
| Fig. 3(a-b): | Stelechocarpus burahol cell viability indicated by fluorescein diacetate (FDA) staining in cell cultures aged 36 days and maintained in liquid MS medium with 7.5 mg L–1 picloram (a) VC: Viable cells and (b) NVC: Non-viable cells |
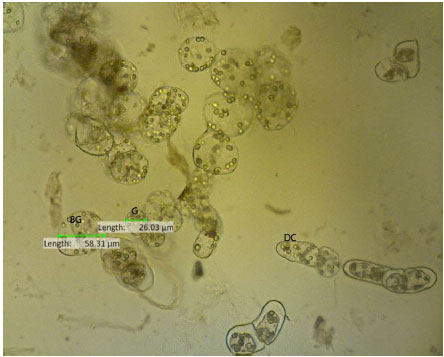 | |
| Fig. 4: | Level of cell differentiation of S. burahol cell culture in MS medium supplemented with 7.5 mg L–1 picloram. G: Globular (<50 μm), BG: Globular (>50 μm), DC: Differentiated cells |
DISCUSSION
The callus that formed from S. burahol mesocarp explants in 7.5 mg L–1 picloram in MS medium was a friable callus. These results are in agreement with those obtained by Stella and Braga16 for Rudgea jasminoides, friable white calluses have been selected on 8.28 μM picloram in modified MS medium. Fitch and Moore17 reported that a high concentration of picloram in the media resulted in a proportionally less differentiated and more translucent white callus in sugarcane culture. Friable callus is a good material for cell suspension culture18.
The lag phase of cell suspension cultures of S. burahol was from 0-6 days, followed by the log phase at 6-30 days of culture and the cells reached the stationary phase between 30-36 days. The highest biomass was obtained at 30 days in the stationary phase. In the suspension culture of B. cordata, the highest biomass was obtained at 14 days in the exponential phase19. In the culture of Orthosiphon stamineus, the lag phase for a period of 6 days, followed by the log phase.
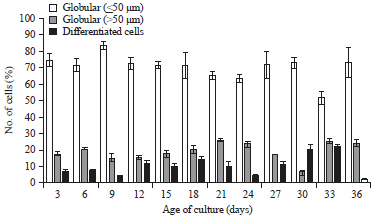 | |
| Fig. 5: | Cell composition based on different levels of differentiation of S. burahol cell culture in MS medium supplemented with 7.5 mg L–1 picloram |
The fresh weight cell culture of O. stamineus reached the maximum biomass after 18 days of culture, followed by a stationary phase20. In Pinus pinaster suspension cultures, growth was kept nearly exponential for a period of 15 days, reaching a maximum of biomass production at day 1821. The data showed that the growth of cell suspension cultures of S. burahol was slower than the growth of other plant cell suspension cultures.
Cell suspension cultures of S. burahol were able to produce flavonoid. This is in agreement with Fu et al.22 that cell suspension culture could produce flavonoids. In the former studies, it has been revealed that the callus culture of S. burahol could also produce flavonoids23. Flavonoid production in the cell suspension culture or callus culture of S. burahol was lower than flavonoid produced by mesocarp. Kumar et al.24 showed that lower concentrations of flavonoid were detected from the callus culture of Heliotropium indicum than in vivo tissue culture (leaf, stem and flower). Tissue differentiation is important for secondary metabolite production. Different levels of differentiation of cultured tissue have differing sensitivities to the environment resulting in different responses25,26.
The production of flavonoids in the cell suspension cultures of S. burahol was not in line with its growth and the maximum production is reached within 15 days at log phase. Verbascoside and linarin production in the culture of B. cordata was in line with the growth and reached the production peak during the stationary phase. However, in the culture of B. cordata, hydroxycinnamic acid content was almost the same for all cultures regardless of age19.
Cell suspension cultures of S. burahol were composed of viable and non-viable cells. The FDA staining distinguished cell viability. The use of FDA staining is based on the nature of the cell membrane, which is permeable to the non-polar substrate and less permeable to fluorescein polar products. The non-polar compound is a substrate that can be digested enzymatically and fluorescent products obtained remain within the cell to produce bright green fluorescent images26. Enzymatic breakdown of the cells can only occur if the cells are live (viable). Therefore, the staining has been proven as a viable indicator of living cells.
These data showed that S. burahol cell suspension cultures at all ages contain slightly differentiated cells. The division process continually produces small cells with the globular-shapes. Some of these cells will grow into globular cells with a larger size. Some cells will differentiate further to form elongated cells. Since only some of the cells undergo further growth and differentiation while continuing the process of cell division, the number of small-sized globular cell numbers remains dominant. The percentage of differentiated cells in cultures of R. mucosa can reach 95% within 20-32 days12, whereas, it can reach 22% within 36 days in S. burahol. This result was most likely due to cell division occurring continuously until the end of culture. This result is reinforced by the data of cell growth, which continued to increase until 36 days. Cell division occurs consistently, producing tiny globular-shape cells. The dominance of globular cells until the end of the growth period also occurred in C. arizonica11. Cupressus macrocarpa cells showed the oblong shape in the 2nd week and during the next 2 weeks, the cells become elliptical11. In Rollinia mucosa, the culture that was grown in medium supplemented with picloram showed that at all concentrations tested had a homogeneous suspension. Additionally, growth ratios and cellular viability were higher (80-90%) than medium supplemented with NAA or 2,4-D12. Although cell suspension cultures for Rollinia mucosa were homogenous, they still had differentiated cells. The highest number of differentiated cells was found at 20-32 days of culture at 70-95%. The medium supplemented with NAA and 2,4-D also showed a homogenous culture, but differentiated cells were still present (elongated cells), particularly during the final phase of the growth cycle12.
The FDA staining results showed that the cells were further differentiated into non-viable cells that finally died. The non-continuing process of cell differentiation was possible because of the absence of conditions that encourage the growth of cells towards further differentiation. The differentiated cells that died would be destroyed; therefore, the number of these cells decreased sharply within 36 days.
CONCLUSION
The cell suspension cultures of S. burahol growth are slow. The production of flavonoids is not in line with the growth of cells and the maximum production is reached within 15 days. The globular-shaped cells dominate the cell suspension culture at all ages.
SIGNIFICANCE STATEMENTS
There are only a few reports in the literature on the in vitro culture of Stelechocarpus burahol. In fact, S. burahol has a significant potential to be developed as medication since it contains many bioactive compounds. The information of its growth and flavonoid production could contribute to the knowledge of flavonoid production using an in vitro culture method.
ACKNOWLEDGMENTS
The authors thank the Directorate General of Higher Education Ministry of Research Technology and Higher Education, Republic of Indonesia, for financially supporting this study as part of the Doctoral Programme at the Universitas Gadjah Mada.
REFERENCES
- Purwantiningsih, A.R. Hakim and I. Purwantini, 2010. Antihyperuricemic activity of the kepel [Stelechocarpus burahol (Bl.) Hook. F. & Th.] leaves extract and xanthine oxidase inhibitory study. Int. J. Pharmacy Pharma. Sci., 2: 123-127.
Direct Link - Tisnadjaja, D., E. Saliman, Silvia and P. Simanjuntak, 2006. Study of burahol (Stelechocarpus burahol (Blume) Hook & Thomson) as an antioxidative compounds containing fruit. Biodiversitas, 7: 199-209.
Direct Link - Sunarni, T., S. Pramono and R. Asmah, 2007. Antioxidant-free radical scavenging of flavonoid from the leaves of Stelechocarpus burahol (Bl.) Hook f. and Th. Indonesian J. Pharm., 18: 111-116.
Direct Link - Costa, P., S. Goncalves, P. Valentao, P.B. Andrade and A. Romano, 2013. Accumulation of phenolic compounds in in vitro cultures and wild plants of Lavandula viridis L'Her and their antioxidant and anti-cholinesterase potential. Food Chem. Toxicol., 57: 69-74.
CrossRefDirect Link - Gadzovska, S., S. Maury, A. Delaunay, M. Spasenoski, D. Hagege, D. Courtois and C. Joseph, 2013. The influence of salicylic acid elicitation of shoots, callus, and cell suspension cultures on production of naphtodianthrones and phenylpropanoids in Hypericum perforatum L. Plant Cell Tissue Organ Culture, 113: 25-39.
CrossRefDirect Link - Bonfill, M., S. Mangas, E. Moyano, R.M. Cusido and J. Palazon, 2010. Production of centellosides and phytosterols in cell suspension cultures of Centella asiatica. Plant Cell Tissue Organ Culture, 104: 61-67.
CrossRefDirect Link - Boonsnongcheep, P., S. Korsangruang, N. Soonthornchareonnon, Y. Chintapakorn, P. Saralamp and S. Prathanturarug, 2010. Growth and isoflavonoid accumulation of Pueraria candollei var. candollei and P. Candollei var. mirifica cell suspension cultures. Plant Cell Tissue Organ Culture, 101: 119-126.
CrossRefDirect Link - Kumar, V., D. Desai and V. Shriram, 2014. Hairy root induction in Helicteres isora L. and production of diosgenin in hairy roots. Nat. Prod. Bioprospecting, 4: 107-112.
CrossRefDirect Link - Jalal, M.A.F., K.H. Overton and D.S. Rycroft, 1979. Formation of three new flavones by differentiating callus cultures of Andrographis paniculata. Phytochemistry, 18: 149-151.
CrossRefDirect Link - Sparapano, L. and G. Bruno, 2004. Cupressus callus and cell suspension cultures: Effect of seiridins on their growth and sensitivity. In vitro Cell. Dev. Biol. Plant, 40: 617-625.
CrossRefDirect Link - Figueiredo, S.F.L., C. Simoes, N. Albarello and V.R.C. Viana, 2000. Rollinia mucosa cell suspension cultures: Establishment and growth conditions. Plant Cell Tissue Organ Culture, 63: 85-92.
CrossRefDirect Link - Hao, G., X. Du, F. Zhao, R. Shi and J. Wang, 2009. Role of nitric oxide in UV-B-induced activation of PAL and stimulation of flavonoid biosynthesis in Ginkgo biloba callus. Plant Cell Tissue Organ Culture, 97: 175-185.
CrossRefDirect Link - Zou, Y., Y. Lu and D. Wei, 2004. Antioxidant activity of a flavonoid-rich extract of Hypericum perforatum L. in vitro. J. Agric. Food Chem., 52: 5032-5039.
CrossRefDirect Link - Stella, A. and M.R. Braga, 2002. Callus and cell suspension cultures of Rudgea jasminoides, a tropical woody Rubiaceae. Plant Cell Tissue Organ Culture, 68: 271-276.
CrossRefDirect Link - Fitch, M.M.M. and P.H. Moore, 1990. Comparison of 2,4-D and picloram for selection of long-term totipotent green callus cultures of sugarcane. Plant Cell Tissue Organ Culture, 20: 157-163.
Direct Link - Estrada-Zuniga, M.E., F. Cruz-Sosa, M. Rodriguez-Monroy, J.R. Verde-Calvo and E.J. Vernon-Carter, 2009. Phenylpropanoid production in callus and cell suspension cultures of Buddleja cordata Kunth. Plant Cell Tissue Organ Culture, Vol. 97.
CrossRefDirect Link - Wai-Leng, L. and C. Lai-Keng, 2004. Establishment of Orthosiphon stamineus cell suspension culture for cell growth. Plant Cell Tissue Organ Culture, 78: 101-106.
CrossRefDirect Link - Azevedo, H., A. Dias and R.M. Tavares, 2008. Establishment and characterization of Pinus pinaster suspension cell cultures. Plant Cell Tissue Organ Culture, 93: 115-121.
CrossRefDirect Link - Fu, C.X., D.X. Zhao, Y. Huang and F.S. Ma, 2005. Cellular aggregate size as the critical factor for flavonoid production by suspension cultures of Saussurea medusa. Biotechnol. Lett., 27: 91-95.
CrossRefDirect Link - Habibah, N.A., S. Moeljopawiro, K. Dewi and A. Indrianto, 2016. Flavonoid production in callus cultures from mesocarp Stelechocarpus burahol. Biosaintifika, 8: 214-221.
CrossRefDirect Link - Kumar, M.S., S. Chaudhury and S. Balachandran, 2014. In vitro callus culture of Heliotropium indicum Linn. for assessment of total phenolic and flavonoid content and antioxidant activity. Applied Biochem. Biotechnol., 174: 2897-2909.
CrossRefDirect Link - Ptak, A., A.E. Tahchy, G. Wyzgolik, M. Henry and D. Laurain-Mattar, 2010. Effects of ethylene on somatic embryogenesis and galanthamine content in Leucojum aestivum L. cultures. Plant Cell Tissue Organ Culture, 102: 61-67.
CrossRefDirect Link - Maurmann, N., C.M.B. de Carvalho, A.L. Silva, A.G. Fett-Neto, G.L. von Poser and S.B. Rech, 2006. Valepotriates accumulation in callus, suspended cells and untransformed root cultures of Valeriana glechomifolia. In Vitro Cell. Dev. Biol. Plant, 42: 50-53.
CrossRefDirect Link