
Hasmiwati .
Subdivision of Parasitology, Faculty of Medicine, Alas University, Padang, Indonesia
Jamsari
Department of Agroecotechnology, Faculty of Agriculture, Alas University, Padang, Indonesia
Yanwirasti
Anatomy Subdivision, Faculty of Medicine, Alas University, Padang, Indonesia
Nuzulia Irawati
Subdivision of Parasitology, Faculty of Medicine, Alas University, Padang, Indonesia
Dahelmi
Department of Biology, Faculty of Mathematics and Natural Sciences, Alas University, Padang, Indonesia
Pakistan Journal of Biological Sciences
Year: 2015 | Volume: 18 | Issue: 6 | Page No.: 273-278







ABSTRACT
Aedes aegypti is the major vector of DHF virus in the tropical and subtropical. The DHF prevention depends on vector control because the vaccine is still in development. Microsatellite has become on effective marker to obtain information about genetic diversity and analyze the structure of genetic population. The aim of this study was to isolate and characterize A. aegypti microsatellite markers, the Dengue Hemorrhagic Fever (DHF) vector in West Sumatra. Sequences containing microsatellites were obtained by enrichment method. Stages of works were as follows: isolation of A. aegypti genomic DNA, restriction with enzymes, ligation with adapters and hybridization by using microsatellite motifs. Furthermore, the candidate fragment contained motifs cloned on plasmid pGEM T easy vector using E. coli DH 5α with blue-white colony screening. The results showed 46 clones were successfully extracted from a total of 152 clones and became microsatellite motifs with repetition: (GA)3, (CTA)3, (GA)3 (TAAG)3, (ACTT)3 (TC)3 (AC)3. Eight pairs of primer were successfully designed from sequences containing microsatellite motifs with feasible flanking regions. The primer evaluation used 32 DNA samples of A. aegypti from 8 cities (population) in West Sumatra. These markers have been successfully amplified 9-17 alleles with amplification products ranging from 129-306 bp, with a high degree of polymorphism. Aedes aegypti microsatellite markers obtained can be used to analyze the structure of genetic population of A. aegypti and the obtained results were the additional microsatellite markers type of A. aegypti than what had previously existed.
PDF Abstract XML References Citation
Received: May 25, 2015;
Accepted: October 02, 2015;
Published: October 14, 2015
How to cite this article
Hasmiwati ., Jamsari, Yanwirasti, Nuzulia Irawati and Dahelmi, 2015. Isolation and Characterization of Aedes aegypti Microsatellite Markers Dengue Hemorrhagic Fever Vector in West Sumatra, Indonesia. Pakistan Journal of Biological Sciences, 18: 273-278.
DOI: 10.3923/pjbs.2015.273.278
URL: https://scialert.net/abstract/?doi=pjbs.2015.273.278
DOI: 10.3923/pjbs.2015.273.278
URL: https://scialert.net/abstract/?doi=pjbs.2015.273.278
INTRODUCTION
Aedes aegypti mosquito, the vector of Dengue Hemorrhagic Fever (DHF) disease is a disease that becomes a major problem in the tropics and subtropics. The DHF is the most important arbovirus disease in humans (Effler et al., 2005; Weaver and Reisen, 2010). This mosquito can also transmit yellow fever virus (Wallis et al., 1983) and Chikungunya virus (Van den Hurk et al., 2010).
Reducing A. aegypti population is the main way to fight against DHF virus because efficient therapy and vaccine for DHF is still in development. According to WHO, prevention of DHF is still simply by controlling the mosquito vector of A. aegypti. Mosquito control strategies that have been developed for decades yet provide maximum results, because it has not been referred to the overall vector data such as vector character itself. Thus, it is necessary for a genetic control strategies that require genetic markers such as microsatellite markers. Genetic control strategies require genetic information in the form of genetic diversity of A. aegypti. Genetic diversity includes genetic structure, gene flow and differentiation among and between the population, which can be studied using several markers such as allozyme, RAPD, RFLP and microsatellite DNA (Lovin et al., 2009). This is a marker that has a high level of confidence (Jarne and Lagoda, 1996).
Microsatellite is also a genetic marker that is often used to study the mating system and population structure (Steffen et al., 1993), linkage, chromosome mapping and population analysis (Silva et al., 1999). According to Robinson et al. (2004), there are five advantages of using microsatellite markers. First, it is distributed in the entire genome. Second, it is codomint. Third it requires a small amount of DNA. Fourth multiple microsatellite alleles can be detected at a single locus using PCR-based filtering. Fifth, the analysis can be carried out semi-automatic.
Research on genetic diversity that is based on microsatellite DNA of A. aegypti has been conducted by Huber et al. (1999), microsatellites as a marker in A. aegypti genetic studies as DHF vector. Ravel et al. (2001, 2002) conducted a preliminary study of the genetics populations of A. aegypti in Mexico. Lovin et al. (2009) did research on the development and validation polymorphic DNA microsatellite and A. aegypti genetics population studies in Haiti. Paupy et al. (2010) combined the morphological data and DNA microsatellites to investigate the morphological and genetic variability of A. aegypti in Niakhar, Senegal. Then Paupy et al. (2012), examined the genetic structure and phylogeography of A. aegypti vector of DHF and yellow fever in Bolivia. Rasheed et al. (2013) examined the structure population of A. aegypti in Pakistan.
West Sumatra is one of endemic areas with high cases of DHF every year. Vector mosquito control strategies undertaken during the course was 3 M (Menutup, Closes; Menguras, Drain and Mengubur, Bury) and breeding place of vector and fogging at the time of the case, result yet significant decrease in DHF cases. Further control needed the data of A. aegypti genetic diversity using microsatellite markers. These markers can be used as a basis for the development of DHF vector mosquito control strategy. The purposes of this study were to isolate and characterize A. aegypti microsatellite markers of DHF vector in West Sumatra.
MATERIALS AND METHODS
Eggs and larvae of A. aegypti were collected from houses in Padang and Solok area in May until October, 2012. Eggs were collected with ovitrap and larvae were taken directly in the bathtub. Both were developed in the laboratory until adult. Genomic DNA was extracted from whole body of Ae. aegypti using cetyltrimethylammonium bromide (CTAB) protocol described by Hoelzel (1994) with modification by Anggraini (1998). A microsatellite-enriched library was obtained using protocols adapted from Edwards et al. (1996) and Zane et al. (2002). Genomic DNA from one individual of A. aegypti (Padang and Solok) was digested with AluI, RsaI and HindII enzyme (Vivantis), the cutting results were ligated then with MluI adaptor (consist of a 21-mer: 5’CTC CTT TTG ACG CGT GGA CTA3’ and a phosphorylated 25-mer at the 5’ end: A 3’ ACA CGA CGA GAA GCA CCT GATp5’ TG). Ligation reaction contained 1 μL DNA, 0.5 μL 21-mer adapter, 1 μL 25-mer adapter, 5 μL 5x Rapid ligation buffer, 1 μL of T4 DNA ligase. PCR reactions were performed under the following conditions: 30 sec initial denaturation at 94°C 1x, followed by 35 cycles of 30 sec at 94oC, 1 min at 60oC, 2 min at 72°C and 10 min at 72°C. DNA was enriched with (AT)n (CT)n (GT)n (AC)n (AG)n (GC)n motifs (Slotman et al., 2007). Microsatellite-enriched DNA fragments were ligated into pGEM-T Easy Vectors (Promega, 1999), which were used to transform DH5α Escherichia coli competent cells (Promega USA Corporation). Positive clones were selected using the β-galactosidase gene and grown overnight with ampicillin. The sequencing process was performed in Macrogen Seoul, Korea, Result was edited by Geneious program 5.5.7 version. Screening of vector sequence was entry into vecscreen in NCBI website, continued with BLASTN to determine the homologues of obtained sequence and compared with GeneBank data result. Microsatellite motifs were found with Microsatellite Finder (Bikandi, 2006).
Primer pairs were designed with Primer 3 from sequences which flank the microsatellite and were tested in DNA of mosquitoes (Padang and Solok). Evaluation was also tested on the DNA derived from six cities/other populations (Painan, Pariaman, Ujung Gading, Lubuk Sikaping, Bukittinggi and Payakumbuh). The PCR was program consisted of 5 min at 94°C followed by 35 cycles of 30 sec at 94°C, 1 min (optimal annealing temperatures; 55-67°C), 2 min at 72°C and 10 min at 72°C. The amplification products were separated under denaturing conditions on a 10% Polyacrylamide gel (PAGE) for 2.5 h at 75 volts using a Mini Protean BIORADs System The microsatellite fragments position was two fragments, it as heterozygous determined by looking at the number of fragments. If there were two fragments were heterozygous and it there was one fragment, it was homozygous. The supporting assumption revealed that all fragments that have the same rate on polyacrylamide gel was the homologous allele (Nei, 1987). Estimation length of alleles was performed using Image J program by converting the distance of marker on each gel.
RESULTS
The fragments of A. aegypti genomic DNA extraction were good enough, because the concentrations varied from 3.6-60.9 ηg μL‾1 and from 1.82-2.52 purity. Restriction using AluI, RsaI and HindII enzyme resulted in 100-1000 bp DNA fragment (Fig. 1). Candidates of microsatellite motif after enrichment obtained good results (Fig. 2).
 | |
| Fig. 1: | An example of DNA fragment isolation results of Aedes aegypti |
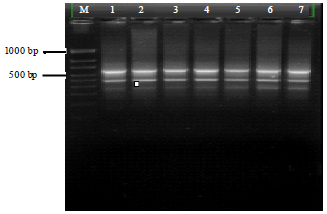 | |
| Fig. 2: | DNA fragments candidate with microsatellite motif, 500 bp of fragment, 400 bp in the samples 1, 2, 3, 4, 5, 6 and 7 |
Forty-six clones were successfully extracted from a total of 152 clones and became candidates of microsatellite motifs (Fig. 3 and 4a), verified again by PCR using 21-mer (Fig. 4b). The results of sequencing using T7SP6 primer contained microsatellite motifs (Table 1).
Characterization of microsatellite motifs was selected based on motif repetition di, tri and tetra-nucleotide (GA, AC, CA, CTA, ACTT and TAAG). The result of primer design can be seen in Table 2. From eight pairs of primer that were designed, only six matched with the sequence of A. aegypti. The results of PCR using six primer pairs in 32 samples from eight populations of A. aegypti are shown in Fig. 5. Nine to seventeen alleles resulted with target sequence, 129-306 bp amplification product, with high polymorphism.
DISCUSSION
Fragment size of 100-1000 bp obtained from the restriction, genomic DNA was selected with high concentration of extracted DNA.The process was done using a three blunt-tipped enzymes simultaneously.
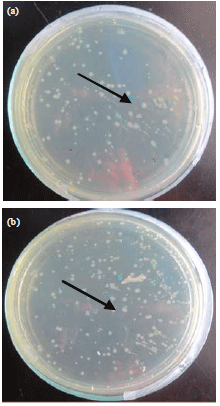 | |
| Fig. 3: | An example a duplicate of colonies resulted from the transformation, (a) Blue colonies, non-recombinant and (b) White colonies, recombinant |
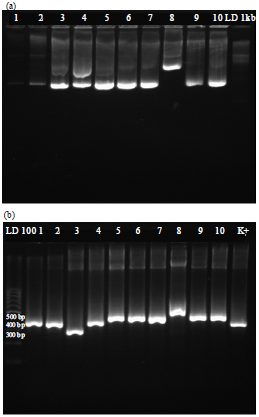 | |
| Fig. 4(a-b): | (a) Electrophoregram of plasmid DNA from recombinant colony and (b) Electrophoregram of recombinant plasmid DNA, PCR with primers 21-mer, LD: Leader, Samples, 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 Plasmid |
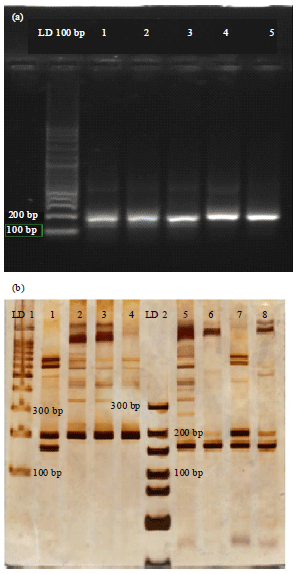 | |
| Fig. 5(a-b): | (a) Electrophoregram of PCR products by agarose gel and (b) Polyacrylamide gel electrophoresis/PAGE, LD: Laeder 1: 100 bp, 2: 10 bp, samples 1, 2, 3, 4, 5, 6, 7 and 8 |
It aimed at improving improve the acquisition of fragment containing the microsatellite motif and facilitate attachment of adapter when ligation (Edwards et al., 1996; Kusumawaty et al., 2005), increasing the chances of getting microsatellite motifs and reducing the same insert fragments (Zane et al., 2002).
The process of enrichment was performed using six types of microsatellite motif; (AT)20, (CT)20, (GT)20, (AC)20, (AG)20 and (GC)20. A total of six microsatellite motifs were grouped into three membrane based on Tm (Edwards et al., 1996; Kusumawaty et al., 2005), continued with hybridization process.
| Table 1: | Microsatellite motif resulted from the analysis of recombinant plasmid |
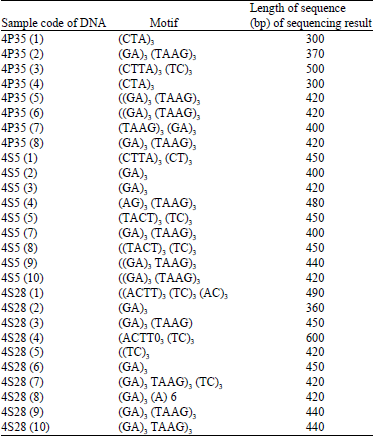 | |
| Table 2: | Microsatellite Primer resulted from DNA genome characterization of Aedes aegypti |
 | |
Hybridization used HYBON nylon membrane (Amersham, USA) which already contained non-labeled radioactive microsatellite motif. In order to get strong binding, oligonucleotide (motif) at the membrane was exposed to ultraviolet light (UV). Fixation process was made in the oven, useful for the formation of covalent bonds between DNA and membranes. This is in line the argument proposed with Reed and Mann (1985), that the strong bonding between membrane and DNA could be performed by UV radiation.
The success of getting a candidate fragment containing A. aegypti microsatellite motif was a very useful result because A. aegypti does not have abundant microsatellites such in plants and animals eukaryotes. Meglecz et al. (2007) revealed that A. aegypti was characterized by a low abundance of microsatellites in the genome.
Microsatellite motifs found did not show much variation of motifs, with short repetition (three times), because of the frequency and type of microsatellite may occur at a rate which varied. Some researchers reported that motif repetition (GT)n in A. maculatus occurred approximately once in every 28 kb (Rongnoparut et al., 1996), once in every 26 kb in A. gambiae and once in every 10,000 kb in A. aegypti (Knudson et al., 1996; Chambers et al., 2007). Three primers of motif in a nucleotide, one primer of motif in tri-nucleotides and two primers of motif in tetra-nucleotides ((CA)3, (GA)3, (AC)3, (CTA)3, (ACTT)3 and (TAAG)3) designed, obtained from 28 samples which had flanking region. Primer designing performed on closest microsatellite motifs or on short flanking regions caused no product in primer that has been designed. Testing in designed primers of 32 samples DNA from eight populations (Padang, Painan, Pariaman, Ujung Gading, Lubuk Sikaping, Bukittinggi, Solok and Payakumbuh) resulted in PCR products. It concluded this result that the pairs of primer were successfully designed and could be used to analyze the structure of A. aegypti population in West Sumatra. Separation of product size for detecting alleles produced homozygous and heterozygous alleles, indicated the presence of polymorphism.
CONCLUSION
There are several conclusions of this study; First, the results of extraction and characterization of the genome of A. aegypti by enrichment method through the process of restriction, hybridization and cloning produced some of microsatellite motifs; di-nucleotides, tri-nucleotides and tetra-nucleotides (AC, AG, CTA, ACTT and TAAG). Second, we were able to design successfully eight pairs of primer, six pairs produced good products of amplification, resulted nine to 17 alleles, with 129 -306 bp amplification product. Third, the alleles were polymorphic and could distinguish whether homozygous and heterozygous individuals. Finally, we obtained A. aegypti microsatellite markers and could be used to analyze the genetic structure of A. aegypti populations and could increase the number of preexist microsatellite markers of A. aegypti.
ACKNOWLEDGMENTS
The author would like to thank and appreciation profusely to HPEQ Project/Improvement of Quality Education Doctor Program (PHKPKPD), Faculty of Medicine, Andalas University by DIKTI Fiscal Year 2012 and 2013 with contract number: 02/PHKPKPD/FKUNAND/ 2013, for financial support to conduct research. Thanks were also extended to all staffs and Biomedical Laboratory analysts in Faculty of Medicine, Alas University.
REFERENCES
- Chambers, E.W., J.K. Meece, J.A. McGowan, D.D. Lovin and R.R. Hemme et al., 2007. Microsatellite isolation and linkage group identification in the yellow fever mosquito Aedes aegypti. J. Hered., 98: 202-210.
CrossRefDirect Link - Edwards, K.J., J.H. Barker, A. Daly, C. Jones and A. Karp, 1996. Microsatellite libraries enriched for several microsatellite sequences in plants. Biotechniques, 20: 758-760.
PubMedDirect Link - Effler, P.V., L. Pang, P. Kitsutani, V. Vorndam and M. Nakata et al., 2005. Dengue fever, Hawaii, 2001-2002. Emerg. Infect. Dis., 11: 742-749.
CrossRefPubMedDirect Link - Huber, K., L. Mousson, F. Rodhain and A.B. Failloux, 1999. Short report: Microsatellite sequences as markers for population genetic studies of the mosquito Aedes aegypti, the vector of dengue viruses. Am. J. Trop. Med. Hyg., 61: 1001-1003.
PubMedDirect Link - Jarne, P. and P.J.L. Lagoda, 1996. Microsatellites, from molecules to populations and back. Trens. Ecol. Evol., 11: 424-429.
CrossRefDirect Link - Lovin, D.D., K.O. Washington, B. deBruyn, R.R. Hemme and A. Mori et al., 2009. Genome-based polymorphic microsatellite development and validation in the mosquito Aedes aegypti and application to population genetics in Haiti. BMC Genom.
CrossRefDirect Link - Meglecz, E., S.J. Anderson, D. Borguet, R. Butcher and A. Caldas et al., 2007. Microsatellite flanking region similarities among different loci within insect species. Insect Mol. Biol., 19: 175-185.
CrossRefDirect Link - Paupy, C., C. Brengues, O. Ndiath, C. Toty, J.P. Herve and F. Simard, 2010. Morphological and genetic variability within Aedes aegypti in Niakhar, Senegal. Infect. Genet. Evol., 10: 473-480.
CrossRefDirect Link - Paupy, C., G. Le Goff, C. Brengues, M. Guerra and J. Revollo et al., 2012. Genetic structure and phylogeography of Aedes aegypti, the dengue and yellow-fever mosquito vector in Bolivia. Infect. Genet. Evol., 12: 1260-1269.
CrossRefDirect Link - Rasheed, S.B., M. Boots, A.C. Frantz and R.K. Butlin, 2013. Population structure of the mosquito Aedes aegypti (Stegomyia aegypti) in Pakistan. Med. Vet. Entomol., 27: 430-440.
CrossRefDirect Link - Ravel, S., N. Monteny, D.V. Olmos, J.E. Verdugo and G. Cuny, 2001. A preliminary study of the population genetics of Aedes aegypti (Diptera: Culicidae) from Mexico using microsatellite and AFLP markers. Acta Trop., 78: 241-250.
CrossRefDirect Link - Ravel, S., J.P. Herve, S. Diarrassouba, A. Kone and G. Cuny, 2002. Microsatellite markers for population genetic studies in Aedes aegypti (Diptera: Culicidae) from Cote d'Ivoire: Evidence for a microgeographic genetic differentiation of mosquitoes from bouake. Acta Trop., 82: 39-49.
CrossRefDirect Link - Reed, K.C. and D.A. Mann, 1985. Rapid transfer of DNA from agarose gels to nylon membranes. Nucl. Acids Res., 13: 7207-7221.
CrossRefDirect Link - Robinson, A.J., C.G. Love, J. Batley, G. Barker and D. Edwards, 2004. Simple sequence repeat marker loci discovery using SSR primer. Bioinformatics, 20: 1475-1476.
CrossRefDirect Link - Rongnoparut, P., S. Yaicharoen, N. Sirichotpakorn, R. Rattanarithikul, G.C. Lanzaro and K.J. Linthicum, 1996. Microsatellite polymorphism in Anopheles maculatus, a malaria vector in Thailand. Am. J. Trop. Med. Hyg., 55: 589-594.
PubMedDirect Link - Silva, F., L. Gusmao and A. Amorim, 1999. Segregation analysis of tetra- and pentanucleotide short tandem repeat polymorphisms: Deviation from mendelian expectations. Electrophoresis, 20: 1697-1701.
CrossRefDirect Link - Slotman, M.A., N.B. Kelly, L.C. Harrington, S. Kitthawe and J.W. Jones et al., 2007. Polymorphic microsatellite markers for studies of Aedes aegypti (Diptera: Culicidae), the vector of dengue and yellow fever. Mol. Ecol. Notes, 7: 168-171.
CrossRefDirect Link - Steffen, P., A. Eggen, G. Stranzinger, R. Fries, A.B. Dietz and J.E. Womack, 1993. Isolation and mapping of polymorphic microsatellites in cattle. Anim. Genet., 24: 121-124.
CrossRefDirect Link - Van den Hurk, A.F., S. Hall-Mendelin, A.T. Pyke, G. Smith and J.S. Mackenzie, 2010. Vector competence of australian mosquitoes for chikungunya virus. Vector-Borne Zoonotic Dis., 10: 489-495.
CrossRefDirect Link - Wallis, G.P., W.J. Tabachnick and J.R. Powell, 1983. Macrogeographic genetic variation in a human commensal: Aedes aegypti, the yellow fever mosquito. Genet. Res., 41: 241-258.
CrossRefDirect Link - Weaver, S.C. and W.K. Reisen, 2010. Present and future arboviral threats. Antiviral Res., 85: 328-345.
CrossRefDirect Link - Zane, L., L. Bargelloni and T. Patarnello, 2002. Strategies for microsatellite isolation: A review. Mol. Ecol., 11: 1-16.
CrossRefDirect Link