
C. Sumathi
Environmental Technology Division, Central Leather Research Institute, Council of Scientific and Industrial Research, Chennai-600 020, Tamil Nadu, India
V. Dillibabu
Environmental Technology Division, Central Leather Research Institute, Council of Scientific and Industrial Research, Chennai-600 020, Tamil Nadu, India
Dash-Koney Madhuri
Queens University, Belfast, United Kingdom
D. Mohana Priya
Environmental Technology Division, Central Leather Research Institute, Council of Scientific and Industrial Research, Chennai-600 020, Tamil Nadu, India
C. Nagalakshmi
Environmental Technology Division, Central Leather Research Institute, Council of Scientific and Industrial Research, Chennai-600 020, Tamil Nadu, India
G. Sekaran
Environmental Technology Division, Central Leather Research Institute, Council of Scientific and Industrial Research, Chennai-600 020, Tamil Nadu, India
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 4 | Page No.: 451-461







ABSTRACT
This study stresses the key role which can be played by Tannery Fleshing (TF) hydrolyzing probiotic Pontibacter spp. in aqua feed formulation and identifies the probiotic strains in the fish gut capable of enhancing the overall growth and immune responses. Probiotics included are Pontibacter species (Pb) and Bacillus megaterium (BM) wherein Lactobacillus (LB) served as control. Experimental diets includes tannery fleshing (TF1), TF+LB strain (TF2), TF+BM strain (TF3), TF+Pb strain (TF4), Fishmeal+BM(TF5 ), Fishmeal+Pb and Control fish meal based diet (TF6). Compared with control, total weight gain (TWG), Specific Growth Rate (SGR), Feed Conversion Ratio (FCR) and Protein Efficiency Ratio (PER) in fish fed with diets supplemented with probiotics were significantly increased (p<0.05). NBT, lysozyme activity, total protein and globulin content were highest in TF4 diet. After challenge with Aeromonas hydrophila, TF4 recorded highest survival and TF1 lowest survival in comparison with the control. Growth and related parameters reveals the effective utilization potential of tannery fleshing probiotic as a feed source. Comparative studies with standard fish meal diets reveals that the fish fed with Pontibacter sps and Bacillus megaterium included feeds enhanced both assimilating capacity and immunological responses in Labeo rohita.
PDF Abstract XML References Citation
Received: March 26, 2013;
Accepted: May 08, 2013;
Published: November 26, 2013
How to cite this article
C. Sumathi, V. Dillibabu, Dash-Koney Madhuri, D. Mohana Priya, C. Nagalakshmi and G. Sekaran, 2014. Dietary Inclusion of Protease Producing Novel Pontibacter spp. and Bacillus megaterium as a Probiotic Enhances Immune Responses in Labeo rohita. Pakistan Journal of Biological Sciences, 17: 451-461.
DOI: 10.3923/pjbs.2014.451.461
URL: https://scialert.net/abstract/?doi=pjbs.2014.451.461
DOI: 10.3923/pjbs.2014.451.461
URL: https://scialert.net/abstract/?doi=pjbs.2014.451.461
INTRODUCTION
Recently a diverse microbial community has been reported from the guts of various fish (Bairagi et al., 2002; Clements and Choat, 1995; Luczkovich and Stellwag, 1993). However, reports on probiotic activity of protease producing Pontibacter sps and Bacillus megaterium and their influence on fish growth and immunological parameters remain unexplored. It is often emphasized that probiotic organism to be used should be isolated from the gastrointestinal tract of host species intended to study (Patel et al., 2010; Verschuere et al., 2000).
Probiotics isolated from mature animals are mostly included in feed of immature animals of same species (Gomez-Gil et al., 2000). Nutritional and health aspects of functional foods incorporating probiotic bacteria, especially Lactobacillus, Bifidobacterium, Bacillus, Streptococcus, Pediococcus, Enterococcus genera and yeast of the Saccharomyces, Aspergillus and Torulopsis genera have received considerable attention and eventually led to numerous claims in the literature (Gomes and Malcata, 1999; Huis in’t Veld and Havenaar 1997; Berge and Storebakken, 1996). Probiotic for fermentation combine the positive images of both probiotics and fermentation organisms that catalyze the proteolytic activities and facilitate the conversion of substrate into simpler compounds to aid in easy assimilation (Kunji et al., 1996). The Bacillus species produce proteases which break down proteins (Farzanfar, 2006) and aids in digestion (Sanders et al., 2003). Bacillus genus are preferred in aquaculture industry as they are non pathogenic and produce most of the antibiotics, amino acids and enzymes (Moriarty, 1990; Gullian et al., 2004). Bacillus megaterium was included as a probiotic in feed formulation. Pontibacter sps. PROLR15, a non motile gram-negative coccobacillus which has 92% similarity with existing Pontibacter strains has been recently identified and only a few publications i.e. pond and desert (Wang et al., 2010; Nedashkovskaya et al., 2005) related to their occurrences are cited. This is first report on the protease producing Pontibacter strain PROLR15 used as a probiotic.
It is well documented that a change in diet selects for a different bacterial community (Olsen et al., 2000; Ringo and Olsen, 1999) which may have an effect on the Specific Growth Rate (SGR) and receives the attention of the environmental scientists and biotechnologist (Al-Hafedh, 1999). The replacement of expensive fish meal in aqua feed with agro industrial wastes or by products have been studied earlier (Kaur and Saxena, 2004). Tannery fleshing, generated from pre-tanning operations constitute about 50-60% among tannery solid wastes that are highest in protein (50.9%). Furthermore, TF protein contains all the essential aminoacids required for fish growth (Sumathi et al., 2012). Hence, in the present study Tannery fleshing was utilized as protein source and Tannery fleshing degrading bacteria as the probiotics.
Previous researchers have reported on the importance of gut flora on immune responses (Galdeano et al., 2007). Nutrition and immunology are inter-related and immunomodulation study based on feed formulation which implies health status of the fish is the impending areas in fisheries research. Effect of immune stimulators in animal and plant products as feed supplements has already been investigated (Iranloye, 2002; Ji et al., 2007). Hence, gut flora and its metabolic activities are the determining factors of nutrition and animal welfare. It is becoming increasingly apparent that the intestinal microflora and its metabolic activities can be an important contributing factor in nutrition, physiology and animal welfare. However, significance of the host intestinal microbial community as a probiotic is limited. Hence, promising prospects are sketched out in accordance with the isolation of tannery fleshing hydrolyzing fish gut bacteria, inclusion of probiotic in the tannery fleshing feed and impact assessment through growth and immune modulating properties.
MATERIALS AND METHODS
Substrate characterization: The Tannery Fleshing (TF) were collected from a commercial tannery near Chennai (Tamil Nadu, India) and were suspended in water. The pH was adjusted to 7.0±0.2 and manually excised into small pieces. The total protein content, fat content and types of amino acids (HPLC) were determined to assess nutritional quality (Sumathi et al., 2012).
Screening and isolation of gut bacteria: The fish gut homogenate was serially diluted in saline and plated on Schaedler HiVegTM Agar (Himedia, India) and incubated for 48 h at 37°C. The major colonies formed on these plates were sub cultured and maintained in slants. The isolates were screened for protease production by inoculating them on skim milk agar plates and gelatin agar plates. The isolates which produced maximum zone were selected and subjected to hydrolysis of TF. These isolates were then inoculated in a liquid media containing MgSO4.7H2O (0.2 g L-1); K2HPO4 (2.0 g L-1); KH2PO4 (2.0 g L-1) and were assayed for higher proteolytic activity. The protease activity was determined after 24 h of incubation. The strains showing higher proteolytic activity were selected, identified and maintained. Among them ANFLR1 strain of Bacillus megaterium and PROLR15 strain of a new Pontibacter spp. exhibited peak protease activity.
Fermentation of TF by ANFLR1 and PROLR15: Fermentation was carried out in a 2 L fermentation chamber at room temperature. The production medium for ANFLR1 contains (g L-1) Sodium chloride, 0.4; Ammonium chloride, 0.015; Dipotassium hydrogen phosphate, 1.25; Potassium dihydrogen phosphate, 0.3 with 5% TF and for PROLR15 was (g L-1) Ferric citrate, 0.1; Sodium chloride, 19.45; Magnesium chloride, 8.8; sodium sulfate, 3.24; Calcium chloride, 1.8; Potassium chloride, 0.55; Sodium bicarbonate, 0.16; Potassium bromide, 0.08; Strontium chloride, 0.034; Boric acid, 0.022; Sodium silicate, 0.004; Sodium fluoride, 0.002; Ammonium nitrate, 0.001; Disodium phosphate, 0.008 with 5% TF. The inoculums were added aseptically into the fermentation chamber inside a laminar air flow. The fermented medium was harvested after 36 h of incubation time along with the sludge and centrifuged at 8000 rpm for 20 min. The supernatant was concentrated by vacuum evaporation, lyophilized and then mixed with the dried sludge.
Experimental feeds and aqua feed formulation: Six experimental feeds were formulated and utilized for the study (TF1, TF2, TF3, T F4, TF5 and TF6) differing in the probiotic inclusion wherein fishmeal based diet served as a control (Table 1). Aqua feed preparation involved separately extruding a mixture of the protein hydrolysates or the raw TF and feed additives into usable form. In experimental feeds raw and fermented TF slurry serves as a sole protein source (38%). Appropriate quantities of dry ingredients were weighed, ground and mixed in a food processor.
Inclusion of probiotic: The sterilized powder of B. megaterium and Pontibacter culture of concentration 108 cells were moistened with other ingredients. The dough was extruded to about 3 mm diameter and then stored in a container -20°C until further use. The proximate compositions of diets were determined according to AOAC protocols (AOAC, 2006).
| Table 1: | Feed composition of probiotic formulated diets |
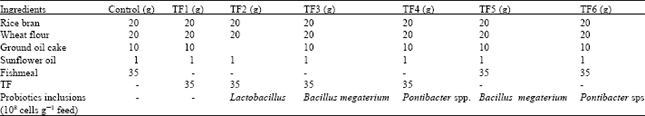 | |
Fish and experimental condition: Labeo rohita fingerlings 10±2 g were obtained from nearest farm in Chennai (India) and acclimatized in 1000 L cylindrical tanks for 15 days with control feed. Three hundred and fifty fingerlings were selected for the study and divided into seven groups. Each group of 50 fingerlings was again divided into two equal duplicate subgroups. Fish were fed twice daily (9:00 a.m. and 17:30 p.m.) to apparent satiation. The fish were fed twice a day and their growth parameters were observed. Blood and serum samples were collected from each group and examined for the following parameters: total protein, albumin, globulin, albumin/globulin ratio, Serum Glutamate Oxaloacetate Transaminase (SGOT), Serum Glutame Pyruvate Transaminase (SGPT), alkaline phosphatase (ALP), serum bactericidal activity, serum lysozyme and Nitroblue tetrazolium (NBT) activities and WBC.
Growth parameters: Growth and nutrient utilization were monitored and analyzed in terms of SGR or percent body weight increase per day and FCR for all the experimental groups were calculated according to Ricker (1979). Recorded mortality data was used for calculating the Relative Percentage Survival (RPS) (Amend, 1981).
Determination of serum biochemical and hematological parameters: Blood was sampled from the caudal vein of the individual fish after anaesthetization. The whole blood was collected in a syringe, allowed to clot for 1 h in microtubes at room temperature followed by 5 h at 4°C and then serum was harvested by centrifuging at 1500xg for 5 min at 4°C. All serum samples were preserved at -20°C prior to analysis. The different serum samples collected were analyzed for total protein following the method of Lowry method; albumin content by Doumas et al. (1971) globulin content (subtracting albumin from the total protein) and albumin:globulin ratio. Total leucocyte count was determined following the method of Shaw (1930). Alkaline phosphatase was estimated according to the method of Bergmayer (1963) using p-nitrophenyl phosphate as a substrate. SGOT and SGPT were determined by method of Reitman and Frankel (1957).
Challenge test against Aeromonas hydrophila: After 60 days of feeding, fish from each subgroup were challenged intraperitoneally with a lethal dose of 1x108 CFU L-1 of A. hydrophila ATCC 49040 (OD adjusted to 0.5 at 456 nm) and observed for a period of the next 10 days for mortality. Estimation of serum and blood parameters were carried out in the surviving fish after day-10 post infection as per the methods described earlier. From each tank, a minimum of eight and six fish were sampled for serum and blood, respectively. The control and experimental fish were subjected to challenge test by inoculating 10 mL of active pathogenic A. hydrophila strains. The reactions of the fish for next 24 to 48 h were observed and mortality rate was determined. Experiments were conducted in triplicates.
Phagocytosis assay: The assay was performed following Siwicki et al. (1990) and Park and Jeong (1996) with slight modification. The phagocytic cells and phagocytized bacteria were enumerated. Phagocytic Ratio (PR) and Phagocytic Index (PI) were determined by enumerating 100 phagocytes per slide under a microscope. An average of three slides was calculated:
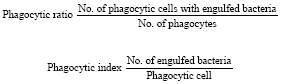 |
Lysozyme activity: Lysozyme activity was analyzed by the turbidometric assay method, originally described by Parry et al. (1965) and modified for microtitre assay by Demers and Bayne (1997). Serum (25 mL per well) was placed in triplicate in a ninety-six-well plate and 175 mL of a suspension of Micrococcus lysodeikticus (75 mg mL-1 in 0•1 M-phosphate buffer with 0•09% (v/v) NaCl, pH 5•8) was added. After the plate had been shaken, the decrease in absorbance at 450 nm was recorded for 5 min. Lysozyme activities were converted to lysozyme concentrations using hen egg white lysozyme as a standard.
Leukocyte metabolic activity assay or respiratory burst activity: The metabolic activity of the phagocytes measured as the ability to produce superoxide anion was assessed by NBT reduction. Superoxide anion production by phagocytic cells was determined by slight modification of the method described by Chung and Scombes (1988) and Misra et al. (2006a).
Serum bactericidal activity: Serum bactericidal activity was estimated following the procedure of Kajita et al. (1990). An equal volume (100 μL) of serum and bacterial suspension (A. hydrophila) was mixed and incubated for 1h at 25°C. A blank control was prepared by replacing serum with sterile PBS. The mixture was then diluted with sterile PBS at a ratio 1:10. The serum-bacterial mixture (100 μL) was pour-plated in nutrient agar and plates were incubated for 24 h at 37 °C. The number of viable bacteria was determined by counting the colonies grown in nutrient agar plates.
Statistical analysis: All data were analyzed by one-way analysis of variance (ANOVA) using the software of the SPSS 11.0 for Windows. When ANOVA identified differences among groups, multiple comparisons among means were made using Duncan's new multiple-range test. The results are presented as Mean±SD and probabilities of p<0.05 were considered significant.
RESULTS
Analysis of growth parameters: Growth parameters data are presented in (Fig. 1). In the present investigation, Bacillus and Pontibacter as probiotic inclusion in Tannery Fleshing feed formulation has positive effect on growth of Labeo rohita depicting significantly higher SGR values and lower FCR values when compared to the control group. Similar results were observed with other immunostimulants (Misra et al., 2006b). This implies that protease producing probiotic facilitated digestion of the protein source (TF) in the fish. The net change in weight of the fish over the control feed was positive (p>0.05) indicating that probiotic incorporated tannery fleshing diet could substantially replace the control feed. Total replacement of fish meal with probiotic containing Tannery fleshing diets had no deteriorating effect and probiotic included in the diet was harmless to fish.
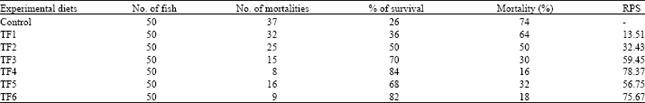
Survival rate: After injection with Aeromonas hydrophila, the first mortality was recorded after 14 h. Mortality was recorded up to 7 days after injection (Table 2). The relative % survival is presented in Table 2. TF4 and TF6 fed groups showed significantly higher survival percentage. Significantly lower survival was recorded for TF1fed groups. After incorporating probiotic with Tannery Fleshing diets, fish species showed enhanced non-specific immune response activities and resistance to experimental challenge with pathogenic bacteria. Mortality percentage was only 8 and 9% in Pontibacter included diets. Relative percentage survival was also high (78 and 76%) in Pontibacter included diets (TF4 and TF6) and coincides with the results of immunostimulant levan included diet (Rairakhwada et al., 2007).
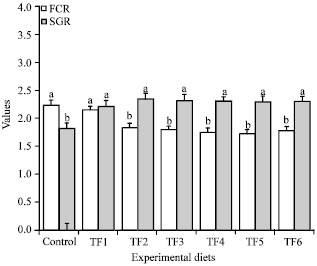 | |
| Fig. 1: | FCR and SGR values of the fish fed with experimental diets after the immunomodulation trial days, Mean values bearing same superscript are not statistically significant, p>0.05, SGR: Ln of final weight-Ln of initial weight)/t (time interval in days)x100, FCR: Feed given (dry weight)/Weight gain (wet weight) |
| Table 2: | Aeromonas challenge test |
 | |
| Table 3: | Serum hematological responses observed before and after the immunostimulation trial period in Labeo rohita fed probiotic included diets |
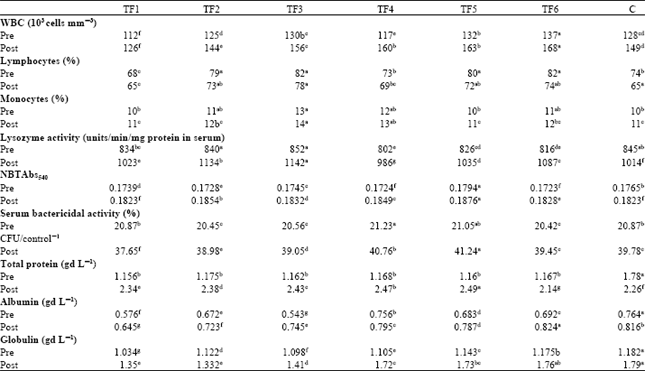 | |
Serum protein, albumin, globulin and A/G ratio: The concentration of total protein in blood serum is used as a basic index for health status of fish. Among the serum protein, albumin and globulin are the major proteins, which play a significant role in the immune responses. There were significant differences in the serum total protein and globulin content among the different experimental groups before and after the challenge study (Table 3). There was marked increase in the WBC count in TF6 fed fish (137-168 103 cells mm-3) when compared to Control (128-149 103 cells mm-3) during pre and post challenge period. A significant decrease (p<0.05) in the serum total protein and globulin content was recorded after challenge with Aeromonas hydrophila either fed with TF1(1.35 g dL-1) diet or control diet (1.89 g dL-1).
In contrast, a significantly higher serum globulin level in TF4-TF6 (1.72, 1.73 and 1.76 g dL-1) groups was noticed during post challenge period. No marked changes in the serum albumin content were recorded either pre-or post-challenge studies in experimental and control diets. The A/G ratio was significantly (p<0.05) increased in the post-challenge period than the pre-challenged period irrespective of the type probiotic inclusion. TF1 group recorded a significantly lower A/G ratio after challenge. During the post-challenge period, a significantly (p<0.05) higher A/G ratio was recorded in the groups fed with Lactobacillus and Pontibacter included Tannery fleshing diets.
Serum lysozyme activity: Changes in lysozyme activity are considerably influenced by the dose and the type of stressors to which fish are exposed. Serum lysozyme activity can be increased using immunostimulants, which may either be due to an increase in the number of phagocytes secreting lysozymes or an increase in the amount of lysozymes synthesized per cell (Edahiro et al., 1990). There was a significant difference (p<0.001) in lysozyme activity among the various treatment groups in the pre and post-challenge period (Table 3). Lysozyme activity was significantly higher (p<0.05) in TF4 groups than the TF3 fed groups during both the pre-and post-challenge period. Similarly, post-challenged fish showed significantly higher lysozyme activity (p<0.05) than the pre-challenged fish fed with probiotic included diets. B. magetarium or Pontibacter Species fermented Tannery Fleshing had higher lysozyme activity in the post-challenge period than in the pre-challenge period. Similar, elevation of lysozyme following dietary incorporation of immunostimulants has been demonstrated in a number of fish species (Paulsen et al., 2003).
Total Leukocyte count: Leukocyte count is considered as an indicator of the health status of fish because of its role in nonspecific or innate immune and inflammatory responses. In the present study, increased WBC count was observed in TF diets. This may be due to metabolic stress mediated by the new protein (Secombes, 1996). The post-challenge increases in leukocyte and monocyte count irrespective of the Tannery fleshing inclusion signify a possible increased infection and inflammatory response mediated by leukocyte against bacteria.
NBT assay or Respiratory burst activity: Respiratory burst activity of phagocytes was measured by reduction of Nitro Blue Tetrazolium (NBT) by intracellular superoxide radicals produced by leukocytes. The production of superoxide radicals as examined by NBT reduction was significantly influenced by the probiotic diets. Maximum increase in the NBT reduction value was observed in Pontibacter incorporated diets and was similar in response to Lactobacillus and Bacillus formulated diets. This further confirms the immunomodulatory properties of Tannery fleshing hydrolyzing probiotics. Similar observations are reported with inclusion of turmeric enriched diet and levan supplemented diets in Cyprinus carpio (Rairakhwada et al., 2007). This result further reveals the positive influence of Probiotic on the survival of Labeo rohita following A. hydrophila infection.
Serum bactericidal activity: Many investigators have reported enhanced bactericidal activity by the phagocytic cells in different fish species (Jorgensen et al., 1993; Sharp and Secombes, 1993). Serum bactericidal activity was lowest in the control group and highest in the experimental groups. The highest activity was observed in probiotics included diet. This shows a positive correlation with the results of NBT assay in which the highest activity was observed in the probiotic incorporated diets. In the remaining test groups (TF1) and control the serum bactericidal activity was very low. The bacterial counts were only slightly lower than the control group and there was no significant difference. Serum bactericidal activity in TF3 to TF4 diets fed fish was significantly (p>0.05) higher than control fish on all the assay days (Table 3). The serum bactericidal activity was higher in the group of fish fed with Pontibacter incorporated diets. In our study, survival was marked to be negatively correlated with total plate count of bacteria, thus indicating a positive correlation of bactericidal power of serum (lysing more number of bacteria and reducing total plate count) with high survivability. The increased serum bactericidal activity in probiotics included diets confirms that various humoral factors involved in innate and/or adaptive immunities are elevated in the serum to protect the host effectively from infection.
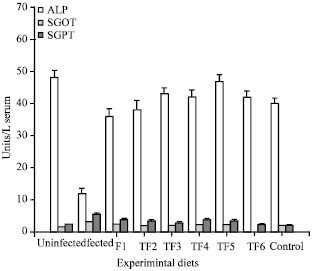 | |
| Fig. 2: | Effect of probiotic included diets on Serum enzymes, The values represent the Mean±SE of triplicates |
Serum ALP, SGOT and SGPT levels: Increased activity of ALP was marked in the group of fish fed with probiotics on different days. Alkaline phosphatase is associated with the absorption of glucose, lipid, calcium and inorganic phosphate (Eguchi, 1995). Increased phosphatase activity indicates higher breakdown of the energy reserve, which is utilized for the growth and survival of fish (Ghosh et al., 2008). It is known that the liver is rich in GOT and GPT and damage to the liver may result in high serum GOT and GPT activities (Fig. 2). Low serum GOT and GPT activities of probiotic formulated diets suggest healthy liver with negligible damage, supporting suppressed hepatic amino acid utilization. Generally, decreased AST and ALT activities in the serum indicates that oxaloacetate and glutamate were not available to the Krebs cycle through this route of transmission (Shakoori et al., 1996).
Phagocytic responses: Most of the immunostimulants used in aquaculture are believed to enhance innate immunity in fish by stimulating leucocyte activity. Leucocytes derive their bactericidal power from a vast range of cellular functions, including phagocytosis, pinocytosis and production of superoxide anions and hydrolytic enzymes (Ellis, 1999). Phagocytic ratios (Fig. 3) and phagocytic indices (Fig. 4) in the fish fed with different probiotic included Fleshing diets were significantly (p>0.05) higher than control fish during the assay period. Diets with Bacillus megaterium and Pontibacter inclusion (TF3 and TF4) possessed highest phagocytic ratio and phagocytic index.
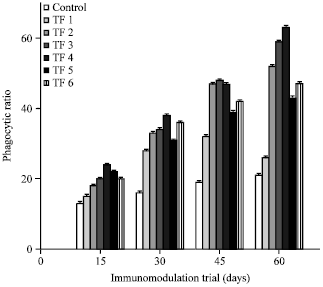 | |
| Fig. 3: | Phagocytic ratio observed on different immunomodulation trial days, The values represent the Mean±SE of triplicates |
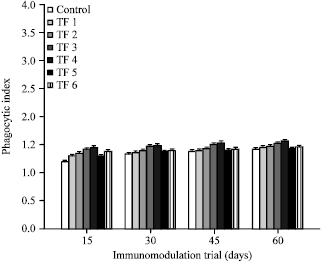 | |
| Fig. 4: | Phagocytic index observed on different immunomodulation trial days, The values represent the Mean±SE of triplicates |
DISCUSSION
Recent data on probiotic applications indicate that non-viable microbial components act in a beneficial manner which aid in development of the immune system, digestive processes and provide nutritional benefits (Ramirez and Dixon, 2003; Rowland et al., 1998). In the present study, the proteolytic strains Bacilllus megaterium and Pontibacter species that facilitates the breakdown of proteins were isolated from Labeo rohita gut (Nibedita and Ghosh, 2008).
Previous researchers have reported the diets with a probiotic supplement exhibited greater growth than those fed with the control diet (Lara-Flores et al., 2003). Various authors have shown that lactic acid bacteria are part of the normal intestinal flora of fish (Ringo and Gatesoupe, 1998; Havenaar et al., 1992) that are commonly associated with nutritious environments (Walstra et al., 1999; Ringo and Olsen, 1999). The administration of L. acidophilus as a probiotic induces a growth promotion effect in, Arctic charr (Salvelinus alpinus), Atlantic salmon (Salmo salar) and rainbow trout (Oncorhynchus mykiss) (Gonzalez et al., 2000). Similar results in the present work suggest that the strain of B. megaterium and Pontibacter species can also be used effectively as probiotic in aquafeed.
Although there are few reports on the effects of exogenous enzyme supplementation in diets for fish, a number of studies on the use of proteases to improve the digestibility of feed ingredients in poultry, pigs and cattle have been published. Exogenous enzymes extracted from bacteria like Bacillus circulans or fungi incorporated into larval feeds were found to aid in digestion and promote better growth and survival in broiler chickens (Ghazi et al., 2002) diets resulted in improved digestibility of protein and increased production of milk and eggs. Correspondingly, protease activity of the probiotic strains enhanced growth and immune responses in Labeo rohita.
Immunostimulants can increase the host's nonspecific immunity by either increasing the number of phagocytes or activating phagocytosis and respiratory burst (Shoemaker et al., 1997) to fight against the microbial pathogen. Innate immunity due to lysozyme is caused by lysis of bacterial cell wall and these stimulate the phagocytosis of bacteria. Many researchers have reported the enhancement of the lysozyme activity, complement activity and bactericidal activity (Selvaraj et al., 2005) due to administration of immunostimulants. Respiratory burst activity of phagocytes measured by reduction of NBT by intracellular superoxide radicals produced by leucocytes showed enhanced activity with diet incorporated with probiotic and the highest activity was recorded in the diet supplemented with Pontibacter included diets. In the present study, increased respiratory burst activity can be correlated with increased bacterial pathogen killing activity of phagocytes.
Variations in lysozyme activity appear to be related to the ability of probiotic strains to adhere to the intestinal mucus. Lysozyme is a humoral non-specific defence protein widely distributed in nature, including fish. Although its exact physiological role is not yet understood, there is a general acceptance that lysozyme is involved in the defence against micro-organisms. Lysozyme hydrolyses N-acetylmuramic acid and N-acetylglucosamine, which are constituents of the peptidoglycan layer of bacterial cell walls. Lysozyme activity in fish serum has been reported to increase after injecting a bacterial product, in response to bacterial infection and after probiotic supplementation (Moyner et al., 1993). Panigrahi et al. (2004) showed significantly higher serum lysozyme activity in rainbow trout fed with Lb. rhamnosus JCM 1136 at 109 CFU g-1 feed for 30 days. The results coincide with the above researchers indicating the potential application of Pontibacter spp. as imuunostimulating probiotic.
These results suggest that probiotics included Tannery Fleshing diet when fed at a medium dose for 60 days could enhance immunity, growth and survival of L. rohita fingerlings which can be attributed to the availability of good quality protein and essential amino acids. Total protein content, globulin content and lysozyme level increased in the fish after feeding with probiotic diets. Increase in albumin/globulin percentage after Tannery fleshing administration in this study corroborates with Mondal et al. (2008). Such enhancement in the lysozyme activity could also be correlated with enhanced phagocytic activity. The in-vivo activation of phagocytic cells by Tannery fleshing probiotics might have also induced other antimicrobial mechanisms, which include release of lysosomal enzymes, cationic peptides and production of reactive oxygen species (Kwak et al., 2003) thereby preventing the adherence and colonization of microorganisms (Alexander and Ingram, 1992) leading to prevention of infection and disease. In the present study, an increased growth rate was observed in Labeo rohita fed feed containing B. megaterium and Pontibacter compared with control and the high rates of establishment of bacterium in the gastro-intestinal tract of fish treated with probiotics may have suppressed the A. hydrophila infection, which ultimately resulted in the higher survival (Kumar et al., 2006; Ziaei-Nejad et al., 2006).
The alkaline phosphatase levels were drastically decreased during the infection in control group fish, but probiotic restored the level to normal as in the uninfected fish. As a result of infection with A. hydrophila, SGOT and SGPT were increased significantly in the infected control fish. However, these elevated levels were normalized by probiotic diets and were similar to the uninfected controls. All these results indicate that probiotic included Tannery fleshing diet increases the resistance of L. rohita so that it can withstand the adverse conditions of a challenge.
Mortality following challenge with A. hydrophila was decreased in the group of fish fed with probiotic incorporated diets. There was an inverse relationship between the mortality rate and the probiotic inclusion in the diet. Probiotics have an important role in disease control strategies for aquaculture and may provide an alternative to the use of antimicrobial compounds. The result suggests that the probiotics used in this study can be used effectively as a commercial product for use in aquaculture. Ongoing study is to find metabolic pathways used by these microorganisms in the alimentary tracts of fish, to explain the lifelong host-microbe homeostasis and evaluate the potential therapeutic applications of the bacterial metabolites.
CONCLUSION
In the present study, an increased growth rate was observed in Labeo rohita fed feed containing B. megaterium and Pontibacter compared with control diet. Pontibacter sps and Bacillus megaterium as probiotic included in Tannery Fleshing diet when fed at a medium dose for 60 days could enhance immunity, growth and survival of L. rohita fingerlings indicates that Tannery fleshing utilizing Pontibacter spp. and Bacillus megaterium isolated from Labeo rohita enhanced overall survival and boosted up the immunity. The observed improvement in fish immune parameters as well as growth and survival using novel Pontibacter spp. and Bacillus megaterium as probiotic bacteria may open a new chapter for screening new strains of probiotic bacteria from fresh water fish Labeo rohita for extensive use in aquaculture.
REFERENCES
- AOAC., 2005. Official Methods of Analysis. 18th Edn., Association of Official Analytical Chemists Inc., Arlington, TX., USA, ISBN: 9780935584752.
Direct Link - Farzanfar, A., 2006. The use of probiotics in shrimp aquaculture. FEMS Immunol. Med. Microbiol., 48: 149-158.
CrossRefDirect Link - Bairagi, A., K.S. Ghosh, S.K. Sen and A.K. Ray, 2002. Enzyme producing bacterial flora isolated from fish digestive tracts. Aquacult. Int., 10: 109-121.
CrossRefDirect Link - Berge, G.M. and T. Storebakken, 1996. Fish protein hydrolyzate in starter diets for Atlantic salmon (Salmo salar) fry. Aquaculture, 145: 205-212.
CrossRef - Chung, C. and C.J. Scombes, 1988. Analysis of events occurring within teleost macrophages during the respiratory burst. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol., 89: 539-544.
CrossRef - Clements, K.D. and J.H. Choat, 1995. Fermentation in tropical marine herbivorous fishes. Physiol. Zool., 68: 355-378.
Direct Link - Demers, N.E. and C.J. Bayne, 1997. The immediate effects of stress on hormones and plasma lysozyme in rainbow trout. Dev. Comp. Immunol., 21: 363-373.
CrossRefPubMedDirect Link - Doumas, B.T., W.A. Watson and H.G. Biggs, 1971. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta, 31: 87-96.
CrossRefPubMedDirect Link - Edahiro, T., M. Hamaguchi and R. Kusuda, 1990. Effect of glycyrrhizine against Streptococcal infection of young yellowtail, Seriola quinqueradiata. Suisanzoshoku, 38: 239-243.
Direct Link - Eguchi, M., 1995. Alkaline phosphatase isozymes in insects and comparison with mammalian enzyme. Comp. Biochem. Physiol. B: Biochem. Mol. Biol., 111: 151-162.
CrossRef - Galdeano, C.M., A. de Moreno de LeBlanc, G. Vinderola, M.E.B. Bonet and G. Perdigon, 2007. Proposed model: Mechanisms of immunomodulation induced by probiotic bacteria. Clin. Vaccine Immunol., 14: 485-492.
PubMedDirect Link - Ghazi, S., J.A. Rooke, H. Galbraith and M.R. Bedford, 2002. The potential for the improvement of the nutritive value of soya-bean meal by different proteases in broiler chicks and broiler cockerels. Br. Poult. Sci., 43: 70-77.
CrossRefDirect Link - Ghosh, S., A. Sinha and C. Sahu, 2008. Dietary probiotic supplementation in growth and health of live-bearing ornamental fishes. Aquacult. Nutr., 14: 298-299.
CrossRef - Gomes, A.M.P. and X.F. Malcata, 1999. Bifidobacterium ssp. and Lactobacillus acidophilus: Biological, biochemical, technological and therapeutically properties relevant for use as probiotics. Trends Food Sci. Technol., 10: 139-157.
CrossRef - Gomez-Gil, B., A. Roque and J.F. Turnbull, 2000. The use and selection of probiotic bacteria for use in the culture of larval aquatic organisms. Aquaculture, 191: 259-270.
CrossRefDirect Link - Gonzalez, C.L., J.P. Enicinas, M.L. Garcia Lopez and A. Otero, 2000. Characterization and identification of lactic acid bacteria from freshwater fishes. Food Microbiol., 17: 383-391.
Direct Link - Gullian, M., F. Thompson and J. Rodriguez, 2004. Selection of probiotic bacteria and study of their immunostimulatory effect in Penaeus vannamei. Aquaculture, 233: 1-14.
CrossRefDirect Link - Iranloye, B.O., 2002. Effect of chronic garlic feeding on some haematological parameters. Afr. J. Biomed. Res., 5: 81-82.
Direct Link - Jorgensen, J.B., G.J.E. Sharp, C.J. Secombes and B. Robertsen, 1993. Effect of a yeast-cell-wall glucan on the bactericidal activity of rainbow trout macrophages. Fish Shellfish Immunol., 3: 267-277.
CrossRefDirect Link - Kajita, Y., M. Sakai, S. Atsuta and M. Kobayash, 1990. The immunostimulatory effects of levamisole on rainbow trout, Oncorhynchus mykiss. Fish Pathol., 25: 93-98.
Direct Link - Nibedita, K. and K. Ghosh, 2008. Enzyme producing bacteria in the gastrointestinal tracts of Labeo rohita (Hamilton) and Channa punctatus (Bloch). Turk. J. Fish. Aquatic Sci., 8: 115-120.
Direct Link - Kaur, V.I. and P.K. Saxena, 2004. Incorporation of brewery waste in supplementary feed and its impact on growth in some carps. Bioresour. Technol., 91: 101-104.
CrossRef - Kunji, E.R.S., I. Mierau, A. Hagting, B. Poolman and W.N. Konings, 1996. The proteotytic systems of lactic acid bacteria. Antonie van Leeuwenhoek, 70: 187-221.
CrossRefPubMedDirect Link - Kwak, J.K., S.W. Park, J.G. Koo, M.G. Cho, R. Buchholz and P. Goetz, 2003. Enhancement of the non-specific defence activities in carp (Cyprinus carpio) and flounder (Paralichthys olivcaces) by oral administration of Schizophyllan. Acta Biotechnologica, 23: 359-371.
CrossRefDirect Link - Lara-Flores, M., M.A. Olvera-Novoa, B.E. Guzman-Mendez and W. Lopez-Madrid, 2003. Use of the bacteria Streptococcus faecium and Lactobacillus acidophilus and the yeast Saccharomyces cerevisiae as growth promoters in Nile tilapia (Oreochromis niloticus). Aquaculture, 216: 193-201.
CrossRefDirect Link - Luczkovich, J.J. and E.J. Stellwag, 1993. Isolation of cellulolytic microbes from the intestinal tract of pinfish, Lagodon rhomboids: Size-related changes in diet and microbial abundance. Mar. Biol., 116: 381-388.
Direct Link - Misra, C.K., B.K. Das, S.C. Mukherjee and P. Pattnaik, 2006. Effect of long term administration of dietary β-glucan on immunity, growth and survival of Labeo rohita fingerlings. Aquaculture, 255: 82-94.
CrossRefDirect Link - Misra, K.C., K.B. Das, C.S. Mukherjee and P. Pattnaik, 2006. Effect of multiple injections of β-glucan on non-specific immune response and disease resistance in Labeo rohita fingerlings. Fish Shellfish Immunol., 20: 305-319.
Direct Link - Mondal, S., T. Roy, S.K. Sen and A.K. Ray, 2008. Distribution of enzyme-producing bacteria in the digestive tracts of some freshwater fish. Acta Ichthyol. Et Piscatorial, 38: 1-8.
Direct Link - Moyner, K., K.H. Roed, S. Sevatdal and M. Heum, 1993. Changes in non-specific immune parameters in Atlantic salmon, Salmo salar L., induced by Aeromonas salmonicida infection. Fish Shellfish Immunol., 3: 253-265.
CrossRef - Nedashkovskaya, O.I., S.B. Kim, M. Suzuki, L.S. Shevchenko and M.S. Lee et al., 2005. Pontibacter actiniarum gen. nov., sp. nov., a novel member of the phylum `Bacteroidetes`, and proposal of Reichenbachiella gen. nov. as a replacement for the illegitimate prokaryotic generic name Reichenbachia Nedashkovskaya et al. 2003. Int. J. Syst. Evol. Microbiol., 55: 2583-2588.
CrossRef - Panigrahi, A., V. Kiron, T. Kobayashi, J. Puangkaew, S. Satoh and H. Sugita, 2004. Immune responses in rainbow trout Oncorhynchus mykiss induced by a potential probiotic bacteria Lactobacillus rhamnosus JCM 1136. Vet. Immunol. Immunopathol., 102: 379-388.
CrossRefDirect Link - Park, K.H. and H.D. Jeong, 1996. Enhanced resistance against Edwardsiella tarda infection in tilapia (Oreochromis niloticus) by administration of protein-bound polysaccharide. Aquaculture, 141: 135-143.
CrossRef - Patel, A.K., J.J. Ahire, S.P. Pawar, B.L. Chaudhari, Y.S. Shouche and S.B. Chincholkar, 2010. Evaluation of probiotic characteristics of siderophoregenic Bacillus spp. isolated from dairy waste. Applied Biochem. Biotechnol., 160: 140-155.
CrossRefDirect Link - Paulsen, S.M., H. Lunde, R.E. Engstad and B. Robertsen, 2003. In vivo effects of β-glucan and LPS on regulation of lysozyme activity and mRNA expression in Atlantic salmon (Salmo salar L.). Fish Shellfish Immunol., 14: 39-54.
CrossRef - Rairakhwada, D., A.K. Pal, Z.P. Bhathena, N.P. Sahu, A. Jha and S.C. Mukherjee, 2007. Dietary microbial levan enhances cellular non-specific immunity and survival of common carp (Cyprinus carpio) juveniles. Fish Shellfish Immunol., 22: 477-486.
CrossRefDirect Link - Kumar, R., S.C. Mukherjee, K.P. Prasad and A.K. Pal, 2006. Evaluation of Bacillus subtilis as a probiotic to Indian major carp Labeo rohita (Ham.). Aquacult. Res., 37: 1215-1221.
CrossRef - Ramirez, R.F. and B.A. Dixon, 2003. Enzyme production by obligate intestinal anaerobic bacteria isolated from oscars (Astronotus ocellatus), angelfish (Pterophyllum scalare) and southern flounder (Paralichthys lethostigma). Aquaculture, 227: 417-426.
CrossRef - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Ringo, E. and F.J. Gatesoupe, 1998. Lactic acid bacteria in fish: A review. Aquaculture, 160: 177-203.
CrossRefDirect Link - Ringo, E. and R.E. Olsen, 1999. The effect of diet on aerobic bacterial flora associated with intestine of Arctic charr (Salvelinus alpinus L.). J. Applied Microbiol., 86: 22-28.
CrossRef - Rowland, I.R., C.J. Rumney, J.T. Coutts and L.C. Lievense, 1998. Effect of Bifidobacterium longum and inulin on gut bacterial metabolism and carcinogen-induced aberrant crypt foci in rats. Carcinogenesis, 19: 281-285.
CrossRefDirect Link - Sanders, M.E., L. Morelli and T.A. Tompkins, 2003. Spore formers as human probiotics: Bacillus, Sporolactobacillus and Brevibacillus. Compr. Rev. Food Sci. Food Saf., 2: 101-110.
Direct Link - Selvaraj, V., K. Sampath and V. Sekar, 2005. Administration of yeast glucan enhances survival and some non-specific and specific immune parameters in carp (Cyprinus carpio) infected with Aeromonas hydrophila. Fish Shellfish Immunol., 19: 293-306.
Direct Link - Ji, S.C., O. Takaoka, G.S. Jeong, S.W. Lee, K. Ishimaru, M. Seoka and K. Takii, 2007. Dietary medicinal herbs improve growth and some non-specific immunity of red sea bream Pagrus major. Fish. Sci., 73: 63-69.
CrossRef - Shakoori, A.R., A.L. Mughal and M.J. Iqbal, 1996. Effects of sublethal doses of fenvalerate (a synthetic pyrethroid) administered continuously for four weeks on the blood, liver and muscles of a freshwater fish, Ctenopharyngodon idella. Bull. Environ. Contam. Toxicol., 57: 487-494.
CrossRefDirect Link - Sharp, G.J.E. and C.J. Secombes, 1993. The role of reactive oxygen species in the killing of the bacterial fish pathogen Aeromonas salmonicida by rainbow trout macrophages. Fish Shellfish Immunol., 3: 119-129.
CrossRef - Siwicki, A.K., D.P. Anderson and O.W. Dixon, 1990. In vitro immunostimulation of rainbow trout (Onchorynchus mykiss) spleen cells with levamisole. Dev. Comp. Immunol., 14: 231-237.
CrossRefPubMedDirect Link - Shoemaker, C.A., P.H. Klesius and J.A. Plumb, 1997. Killing of Edwardsiella ictaluri by macrophages from channel catfish immune and susceptible to enteric septicemia of catfish. Vet. Immunol. Immunopathol., 58: 181-190.
CrossRef - Sumathi, C., D. Mohanapriya, A.B. Mandal and S. Ganesan, 2012. Production of different proteases from fish gut microflora utilizing tannery fleshing. Eng. Life Sci., 12: 223-237.
CrossRef - Verschuere, L., G. Rombaut, P. Sorgeloos and W. Verstraete, 2000. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev., 64: 655-671.
CrossRefPubMedDirect Link - Wang, Y., K. Zhang, F. Cai, L. Zhang, Y. Tang, J. Dai and C. Fang, 2010. Pontibacter xinjiangensis sp. nov., in the phylum `Bacteroidetes` and reclassification of [Effluviibacter] roseus as Pontibacter roseus comb. nov. Int. J. Syst. Evol. Microbiol., 60: 99-103.
CrossRef - Ziaei-Nejad, S., M.H. Rezaei, G.A. Takami, D.L. Lovett, A.R. Mirvaghefi and M. Shakouri, 2006. The effect of Bacillus spp. bacteria used as probiotics on digestive enzyme activity, survival and growth in the Indian white shrimp Fenneropenaeus indicus. Aquaculture, 252: 516-524.
CrossRefDirect Link