
Yasser A. Ahmed
Faculty of Veterinary Medicine, South Valley University, Qena 83523, Egypt
Soha A. Soliman
Faculty of Veterinary Medicine, South Valley University, Qena 83523, Egypt
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 18 | Page No.: 911-919







ABSTRACT
The current study was undertaken to describe the main histological development stages of long bones (tibia and femur) from Japanese quail (Coturnix coturnix japonica) embryos. Whole Limbs or just tibia and femur of fifty Japanese quail embryos of different ages were fixed and embedded in paraffin or Spurr’s resin. Paraffin and semi-thin, respectively, were undertaken and examined with light microscopy. Limb bud was established at day 5 of incubation. Mesenchymal cells differentiated into chondrocytes forming a cartilage template in the position of the future tibia and femur at day 6 of incubation. At day 7 of incubation, the cartilage template enlarged and had the shape and position of the future tibia and femur. At day 8, central chondrocytes underwent hypertrophy and were surrounded by a periosteal bone collar. Cellular and vascular invasion from the bone collar into the central zone of the cartilage template, cartilage resorption and formation of marrow tunnel and finally peripheral calcification was seen. Vascular cartilage canals penetrating the epiphysis were observed at day 9 and the canals gradually increased in thickness and number toward the hatching day. Articular epiphyseal growth cartilage with resting, proliferative and hypertrophic zones was clearly established by day 10 of incubation. After 17 days of incubation, the zonation of the articular epiphyseal cartilage were much clear, many cartilage canals were present within the epiphyses. In epiphyses of tibia but not femur, foci of chondrocytes hypertrophy were noticed close to the cartilage canals. The current study timed the main histological sequences of development of tibia and femur of embryonic quail.
PDF Abstract XML References Citation
Received: December 11, 2012;
Accepted: January 19, 2013;
Published: April 15, 2013
How to cite this article
Yasser A. Ahmed and Soha A. Soliman, 2013. Long Bone Development in the Japanese Quail (Coturnix coturnix japonica) Embryos. Pakistan Journal of Biological Sciences, 16: 911-919.
DOI: 10.3923/pjbs.2013.911.919
URL: https://scialert.net/abstract/?doi=pjbs.2013.911.919
DOI: 10.3923/pjbs.2013.911.919
URL: https://scialert.net/abstract/?doi=pjbs.2013.911.919
INTRODUCTION
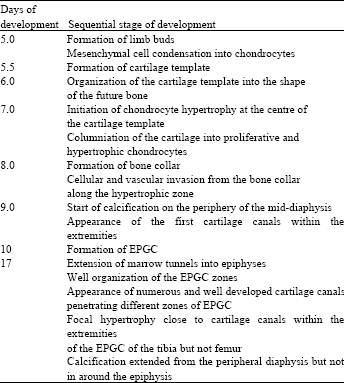
Ossification is the process of development and growth of the bones in both mammals and birds occurs through intaramembranous and (intracartilagenous) endochondral ossification. Intra-membranous ossification is the direct transformation of mesenchymal tissue into bone tissue without formation of cartilage template, while endochondral ossification is the differention of the mesenchymal tissue into cartilage template, which is later transformed into bone tissue. During embryonic limb development, the undifferentiated mesenchymal cells differentiate into cartilage cells or chondrocytes, which secrete a specific collagen type II- and proteoglycan-cartilage rich matrix forming a cartilage template that mimics the outline of the adult bone and develops at its final position reviewed by Mackie et al. (2008). In mammals, the subsequent stages are nearly understood. It has been reported that the chondrocytes at the middle of the cartilage template undergo hypertrophy followed by calcification of its surrounding matrix. Then the hypertrophic chondrocytes physiologically die and the cartilage matrix is resorped. Vascular and cellular invasion of the resorped calcified cartilage initiate deposition of bone tissue over the calcified resorped matrix to establish a Primary Ossification Centre (POC). More enzymatic and cellular degradation of the matrix at the distal end of the POC leads to formation of bone marrow reviewed by Goldring et al. (2006). Similarly, a series of changes starting with central chondrocytes hypertrophy, calcification of matrix, physiological death of hypertrophic chondrocytes, matrix resorption, vascular and cellular invasion of the resorped cartilage finally, lead to establishment of Secondary Ossification Centre (SOC) in each extremity of the future bone. Ossification expands from the POC and SOC leaving zones of cartilage between the two ossification centres: the Physeal Growth Cartilage (PGC) between the POC and SOC and Articular-Physeal Growth Cartilage (APGC) above the SOC (Rivas and Shapiro, 2002). In avian, the mechanism of ossification is not much clear as mammals due to the lack of specific histological works done on the avian embryonic lone bones. A histological study of 6-17-day-embryonic-quail long bones shown that the chondrocytes of the cartilage template organized into three zones. Central hypertrophic chondrocytes, which is surrounded on both side with flattened chondrocytes and peripheral epiphyseal chondrocytes. The cartilage matrix resorption occurs without formation of POC. However, ossification occurs around the cartilage outline in the perichondrium; perichondral ossification (Pourlis et al., 2006). In the femur from 14-20-day-old chick embryos, calcification of the matrix does not occur, however the resorped cartilage is directly replaced by bone marrow POC development (Roach and Shearer, 1989). It was reported that in turkey embryos the POC appears at the mid-region of femur and tibia at day 12 and extends into the proximal and distal region, however the authors did not shown a detailed Fig. 1 for the POC (Atalgin and Kurtul, 2009). In both avian and mammalian long bone, the ossification proceeds and the bone increases in length until no cartilage remains except at the articular surface; articular cartilage. Once the animal or bird achieve skeletal maturity, bone stop growing in length, however the bone continue modelling and remodelling to less extend than the in the growing stages reviewed by Mackie et al. (2008). Although, the cartilage is avascular tissue, the embryonic growth cartilage of the long bone epiphyses is invaded by many perichondrial tubular vascular channels, cartilage canals (Blumer et al., 2004). There are many studies on the structure of the growth cartilage of chicken with tibial dyschondroplasia, however, information on the normal process of endochondral ossification of the pre-hatching quail is very limited. The few available studies on embryonic skeletal development of quail are mainly concerning with the timing of chondrification and ossification of different bones but they did not present the histological structure of the quail growth cartilage and its ossification stages (Pourlis et al., 1998; Nakane and Tsudzuki, 1999). The aim of the current study was to describe the sequences of chondrification and ossification in the tibia and femur of the pre-hatching quail during different stages of embryonic development.
MATERIALS AND METHODS
Egg incubation: Fertilized eggs weighting 10-12 g, of the Japanese quail (Coturnix coturnix japonica) were kindly provided by the Quail production unit related to the Department of Histology, Faculty of Veterinary Medicine, South Valley University, Qena, Egypt. The eggs were put in the incubator (c10, from the Poultry Technical Office, Alexandia, Egypt) at 37.5°C and the relative humidity was 65%, where the eggs were rotated every 1 h after the 3rd day of incubation.
Samples processing for light microscopy: Eggs were collected in the morning and carefully opened at the wide part, and at least five embryos at different stages of pre-hatching development (5, 5.5, 6, 7, 8, 9, 10, 12, 15 and 17th) day of incubation were quickly collected, rinsed in a phosphate-buffered saline. The whole limbs or tibia and femur were dissected, fixed in either 4% neutral-buffered para-formaldehyde (pH 7.4) for 48 h at 4°C. Some specimens were decalcified in 5% Ethylene-Diamine-Tetra-Acetic acid (EDTA) when calcification was established, while others were directly processed without decalcification for Von Kossa staining. Specimens were processed for either paraffin or plastic sectioning. For paraffin sectioning, specimens were dehydrated in ascending grades of ethanol and embedded in paraffin wax blocks. Paraffin sections (3-5 μm thickness) were undertaken and stained with hematoxylin and eosin (H and E), safranin-O, Crossman’s trichrome, Periodic Acid-Schiff’s reagent (PAS) and Von Kossa staining. For plastic sectioning, specimens were fixed in 2.5% glutaraldehyde in 4% paraformaldehyde (pH 7.4) for 58 h at 4°C, post-fixed in 1% osmium tetroxide, dehydrated in ascending grades of acetone and embedded in Spur's resin. Semi-thin sections (0.5 μm thicknesses) were taken, stained with toluidine blue. Paraffin and semi-thin sections were examined with light microscopy. The common international ethical rules were followed as much as possible during sample collection and processing.
RESULTS
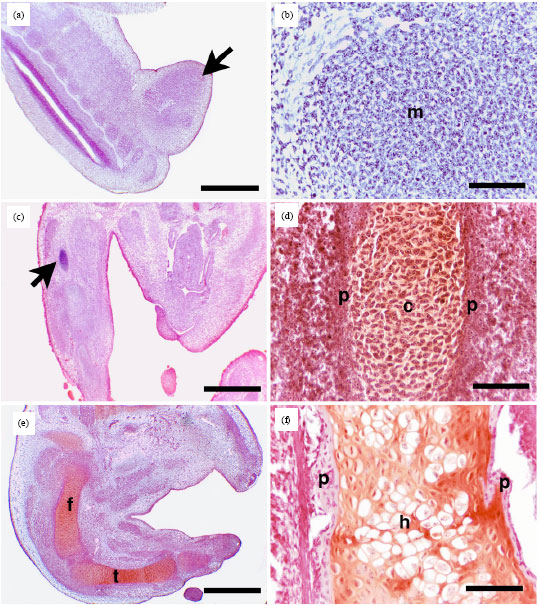
Pattern of chondrification and calcification in the tibia and femur of the pre-hatching Japanese quail: After 5 days of incubation of fertile Japanese quail eggs, the limb bud was established (Fig. 1a). Condensed undifferentiated mesenchymal cells were seen within the limb bud (Fig. 1b). Mesenchymal cells differentiated into chondrocytes forming a cartilage template which was observed for the first time at the position of the future tibia and femur at 5.5th day of incubation (Fig. 1c). The cartilage template showed safranin-O-positive glycosaminoglycan-rich matrix and was surrounded by a perichondrium (Fig. 1d). The chondrogenic differentiation and proliferation of mesenchymal cells progressed and the cartilage template enlarged; where the femur and tibia were formed of hyaline cartilage at the 6th day of incubation (Fig. 1e). Perichondrium enclosed the cartilage template from outsides. At the day 7 of incubation, the chondrocytes of the mid-region of the cartilage template started hypertrophy surrounded on both side with flattened chondrocytes. Many sequences were observed at day 8 of incubation. Most of the chondrocytes in the centre of the cartilage template were truly hypertrophic cells after 8 days of incubation (Fig. 1f).
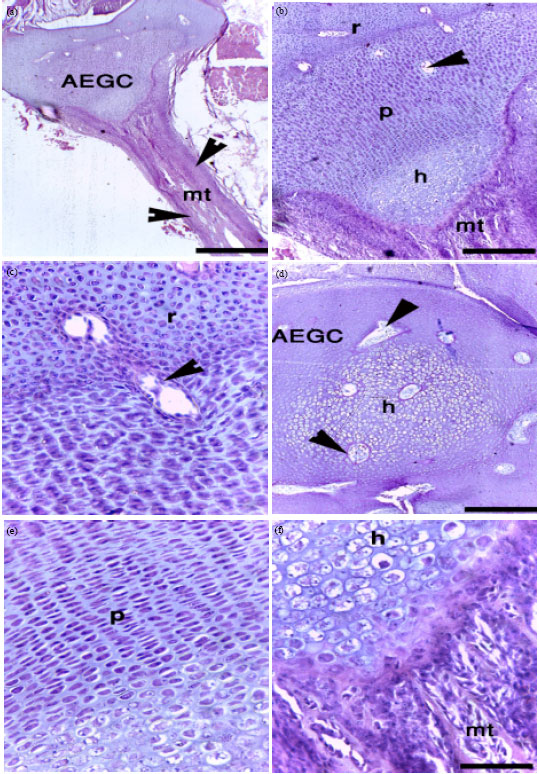 | |
| Fig. 1(a-f): | Formation of the cartilage template in the tibia and femur of the pre-hatching quail. Paraffin (a-f) sections of the limb from 5- (a, b), 5.5- (c, d), 7- (e) and 8- (f) day-quail embryos stained with H and E (a-c) and safranin-O (d-e). A: Limb bud (arrow). b: Mesenchymal cell condensation in the limb bud (m). c: Cartilage template in the tibia (arrow). d: Safranin-O positive cartilage template (c) Surrounded by perichondrium (p). e: Cartilaginous femur (f) Tibia (t), f: hypertrophic chondrocytes (h) At the middle of cartilage template surrounded with perichondrium. Bars = 200 and parts b, c, e and 20 μm in b, d, f |
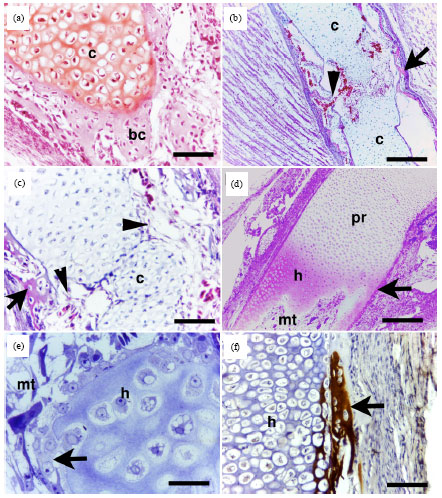
Periosteal bone collar was observed during this sage of development (Fig. 2a). At day 9 of incubation, the hypertrophic chondrocytes in the centre of the cartilage template underwent physiological cell death and the cartilage matrix was invaded by vascular and cellular elements (Fig. 2b, c). The cartilage matrix was gradually resorped (Fig. 2b, e) and in advanced stages a marrow tunnels was formed and extended deeper into the epiphysis (Fig. 2d) where some of chondrocytes were in directed contact with marrow cells (Fig. 2e). Calcification of the embryonic quail growth cartilage could only be observed after 9 days of incubation. Calcification was confined to the periphery of the mid-diaphysis and become more prominent later on but has never been seen within the matrix of the hypertrophic cartilage before day 18 days at the distal ends of hypertrophic cartilage closed to the marrow tunnels extended into the epiphysis. The epiphyses were formed completely from AEGC with chondrocytes organized into resting, proliferative and hypertrophic zones at day 10 of incubation and by progression in age, the zonation was much clear.
Development of the cartilage canals in the femur and tibia of the pre-hatching Japanese quail: Cartilage canals were observed for the first time at the day 9 of incubation, where unbranched small cartilage canals extend from the perichondrium into the extremities of the EPGC between the future epiphysis and PGC (Fig. 3a). The wall of the canals was first not well defined from the surrounding cartilage tissue. One day later, the canals became better defined and contained blood capillaries of different sizes and mesenchymal cells (Fig. 3b). At the day 12 of incubation, the canals became thicker and numerous (Fig. 3c) and contained large blood capillaries and abundant mesenchymal cells (Fig. 3d). Later on, the cartilage canals penetrated down into the proliferative zone (Fig. 3e) and the cartilage surrounding the canals in this area was less cellular with small chondrocytes concentrically arranged around the canals (Fig. 3f). Some canals appeared to be originated from the articular surface and pass down into the matrix of the EPGC. The cartilage canals became thicker and well developed by the advancing days of incubation.
Histology of the tibia and femur of the Japanese quail at the last day of incubation: After 17 days of incubation and just before hatching the tibia and femur of the Japanese quail were similar in shape to and in the same position of the adult bones. Each bone was formed of a bone diaphysis and two large epiphyses. The epiphyses were completely made of AEGC (Fig. 4a, b), however, the diaphysis was surrounded by bone collar and contained irregular central marrow tunnels that extended into the epiphyses (Fig. 4a, b). The chondrocytes in the EPGC were well organized into resting, proliferative and hypertrophic zones (Fig. 2b, c). No POC were observed, so no PGC or cartilage plate was found. Chondrocytes underwent focal hypertrophy in the extremities of the tibia, however this focal hypertrophy was not observed in the femur during the same stage of development (Fig. 4d).
Summary of the sequences of the long bone development of the pre-hatching Japanese quail: The main development stages of tibia and femur in the embryonic quail are summarized in Table 1.
DISCUSSION
The current study was undertaken with the aim of describing and timing the most dramatic events of the endochondral ossification in the pre-hatching quail. To achieve this aim, paraffin and semithin sections of the tibia and femur from 5-17-day-quail embryos were examined with the light microscopy and the most important sequences were summarized in Table 1.
| Table 1: | Sequential embryonic developmental stages in tibia and femur of Japanese quail |
 | |
 | |
| Fig. 2(a-f): | Cartilage resorption and calcification of the tibia and femur of the pre-hatching quail. Paraffin (a-c, e and semithin d, e) sections of the femur (a-e) from 8- (a), 9- (b, c), 10 (d) and 12- (e, f) day-quail embryos stained with safranin-O (a), Crossman’s trichrome (b, c), toluidine blue (d, e) and Von Kossa staining (e). a: Cartilage matrix (c) surrounded by periosteal bone collar (bc). b: Cellular and vascular invasion (arrowheads) of cartilage (c) from bone collar (arrow). c: Higher magnification from b. Note cellular and vascular invasion (arrowheads) of cartilage (c) from bone collar (arrow). d: Part of AEGC with hypertrophic (h) and proliferative (pr) zones close to marrow tunnels (mt). e: cartilage-marrow tunnel erosion surface (arrow), note hypertrophic chondrocytes (h) and marrow a tunnel cavity (mt).f: Calcification (arrow) of the periphery of hypertrophic zone (h). Bars = 20 μm in a, c, f, 80 μm in b, d and 8 μm in e |
The sequences associated with mesenchymal cells condensation within the limb bud, formation and elongation of a cartilage template to take the shape and position of the future long bone, hypertrophy and death of the chondrocytes in the middle of the cartilage template, formation of periosteal bone collar, followed by cellular invasion from the perichondrium into the middle cartilage and formation, chondrocytes death, cartilage resorption and formation of irregular marrow tunnels with no signs of calcification of the cartilage matrix and no formation of POC. The remnant cartilage was termed as EPGC, where the chondrocytes were organized into resting, proliferative and hypertrophic chondrocytes. This observation is supported by a similar observation in chicken (Roach and Shearer, 1989). In mammals, the cartilage matrix is markedly calcified before to resorption and formation of POC and PGC are well described (Mackie et al., 2008).
 | |
| Fig. 3(a-f): | Development of the cartilage canal of the tibia and femur of the pre-hatching quail. Paraffin (a, c-f) and semithin (b) sections of the tibia (a-d) and femur (e, f) from 9- (a), 10- (b), 12- (c, d), 17- (e, f) day quail embryos stained with safranin-O (a), toluidine blue (b), H and E (c, d) and Crossman trichrome (e, f). Arrows indicate cartilage canals (a-f). Note proximal tibia (ti) and distal femur (f). Bars = in a, f, μm (b), μm (c, e, g) and μm (h). Bars = 200 μm in a, c, 8 μm in b, 20 μm in d, f and 80 μm in e |
Many molecular signals have been reported to control the embryonic bone development. Mesenchymal cell condensation, which is essential for cell-cell signalling (Hall and Miyake, 1995), needs TGF-β to initiate the cellular condensation (Leonard et al., 1991). The expressional factor, Sox 9 plays an important role in the differentiation of mesenchymal cells into chondrocytes (Bi et al., 1999) to form a safranin-O positive cartilage template surrounded by a perichondrium as shown in the current study. The cartilage template contains flattened proliferative chondrocytes that undergo further hypertrophic differentiation at the middle of the cartilage template. The chondrocyte in transition between proliferative and hypertrophic chondrocytes (pre-hypertrophic chondrocytes) express Indian hedgehog (Ihh), inducing the parathyroid hormone-related protein (PTHrP) expression in the perichondrial cells (Bitgood and McMahon, 1995). Parathyroid hormone-related protein has receptors on the pr-hypertrophic chondrocytes, PTH/PTHrP-receptors (Juppner et al., 1991). Activation of PTH/PTHrP by the PTHrP delays the hypertrophy of the chondrocytes allowing elongation of the cartilage template (Vortkamp et al., 1996). While 2 and Runx 3 are expressed by the pr-hypertrophic and hypertrophic chondrocytes induce more hypertrophy (Yoshida et al., 2004). Transcriptional factors, Ihh, PTHrP and Runx 2 and 3 act together to regulate the chondrocytes proliferation/ hypertrophy cycles of the cartilage template. The hypertrophic chondrocytes are marked by the expression of collagen type X and alkaline phosphatise (Simsa and Ornan, 2007) which are needed for matrix calcification, and subsequently formation of the POC.
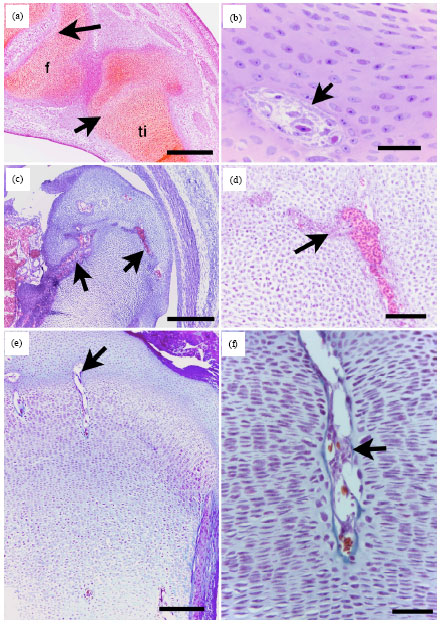 | |
| Fig. 4(a-f): | Organization of the chondrocytes in the EPGC of the tibia and femur of the pre-hatching quail. Paraffin section stained with H and E (a-d) of the femur (a-c) and tibia (d) of 17-day quail embryos. a: the EPGC in epiphysis and the diaphysis with marrow tunnels (mt) and surrounded with bone collar (arrows). B: Cartilaginous head of the femur; note resting (r), proliferative (p) and hypertrophic (h) zones close to the marrow tunnels (mt). c: Higher magnification of b; note resting (r), proliferative (p) and hypertrophic (h) close to the POC. D: Focal hypertrophy of the chondrocytes (h). Arrowheads in b-c indicate cartilage canals. Bars = 200 μm in A, 80 μm in b, c and 20 μm in d |
The current study shown that the perichondrium close to the hypertrophied central chondrocytes of the cartilage template underwent intamembranous ossification and rings of bone or bone collar were established in this part. A similar observation has been reported in a previous study in quail (Pourlis et al., 2006). Matrix resorption occurs in both mammalian and avian growth cartilage. The difference is that resorption in mammalian growth cartilage occurs after calcification of the cartilage matrix, while in avian growth cartilage, resorption occurs even without calcification. Calcification of cartilage matrix in mammals may is important to strength the bone and delay the rate of resorption to give a time for the formation of POC. In avian, the matter is different, calcification doesn't occur so the bone is less strength and less pressure made on the embryo within the small 10 g weight egg of quail and rapid resorption to accelerate the bone formation. We noticed that at the last day of pre-hatching and in post-hatching development, calcification occurs and increases (unpublished work). The bone collar is likely to limit the expansion of the cartilage template beyond the size and shape of the future long bone. The invading cells from the periosteal bone collar are likely to be osteoclasts, which are known as a major source of metalloproteinases (MMPs). Metalloproteinases are essential for the breakdown of the cartilage matrix after death of the hypertrophic chondrocytes, initiating resorption of the cartilage matrix (Sasano et al., 2002). The EPGC was established in both sides of the POC as presented in the current study. Previous studies presented similar results in turkey (Simsa and Ornan, 2007) and in quail (Pourlis et al., 2006). However, unlike the current study, they did not follow the changes of the EPGC. The chondrocytes of the EPGC was organized into resting, proliferative and hypertrophic chondrocytes, similar to growth cartilage of adult bone (Rivas and Shapiro, 2002). The EPGC was invaded by vascular cartilage canals from the perichondrium into the epiphysis and the number and thickness of the canals increased by advancing in age. This in agreements with other species such as rabbits (Rivas and Shapiro, 2002) and chickens (Blumer et al., 2004). Cartilage canals provide not only the vasculature and nutrition to the chondrocytes but the undifferentiated mesenchymal cells that may contribute to formation of the SOC (Burkus et al., 1993; Blumer et al., 2004; Blumer et al., 2005). The last dramatic event in the embryonic long bone development in the quail was the focal hypertrophy of the chondrocytes close to the cartilage canals in the epiphysis of tibia but not femur. The hypertrophy of chondrocytes will facilitate the formation of the SOC in the tibia in the post-hatching growing quail. The femur may show this hypertrophic foci and subsequent SOC formation later than the tibia or may not have a SOC, which will be interesting to follow up in the post-hatching birds. In our study (submitted for publication), we found that only the femur but not the tibia will have SOC.
CONCLUSION
In conclusion, the current study described the histological developmental stages of tibia and femur of the pre-hatching quail from 5 to 17 days of incubation. The study will continue to look at the changes of the endochondral ossification of the tibia and femur of post-hatching growing quail.
REFERENCES
- Atalgin, S.H. and I. Kurtul, 2009. A morphological study of skeletal development in turkeys during the pre-hatching stage. Anat. Histol. Embryol., 38: 23-30.
CrossRefPubMedDirect Link - Bi, W., J.M. Deng, Z. Zhang, R.R. Behringer and B. de Crombrugghe, 1999. Sox9 is required for cartilage formation. Nat. Genet., 22: 85-89.
PubMedDirect Link - Bitgood, M.J. and A.P. McMahon, 1995. Hedgehog and Bmp genes are coexpressed at many diverse sites of cell-cell interaction in the mouse embryo. Dev. Biol., 172: 126-138.
CrossRefPubMedDirect Link - Blumer, M.J.F., H. Fritsch, K. Pfaller and E. Brenner, 2004. Cartilage canals in the chicken embryo: Ultrastructure and function. Anat. Embryol., 207: 453-462.
CrossRefDirect Link - Blumer, M.J.F., S. Longato, E. Richter, M.T. Perez, K.Z. Konakci and H. Fritsch, 2005. The role of cartilage canals in endochondral and perichondral bone formation: Are there similarities between these two processes? J. Anat., 206: 359-372.
CrossRefDirect Link - Burkus, J.K., T.M. Ganey and J.A. Ogden, 1993. Development of the cartilage canals and the secondary center of ossification in the distal chondroepiphysis of the prenatal human femur. Yale J. Biol. Med., 66: 193-202.
Direct Link - Goldring, M.B., K. Tsuchimochi and K. Ijiri, 2006. The control of chondrogenesis. J. Cell Biochem., 97: 33-44.
PubMedDirect Link - Hall, B.K. and T. Miyake, 1995. Divide, accumulate, differentiate: Cell condensation in skeletal development revisited. Int. J. Dev. Biol., 39: 881-893.
PubMedDirect Link - Juppner, H., A.B. Abou-Samra, M. Freeman, X.F. Kong and E. Schipani et al., 1991. A G protein-linked receptor for parathyroid hormone and parathyroid hormone-related peptide. Science, 254: 1024-1026.
CrossRefDirect Link - Leonard, C.M., H.M. Fuld, D.A. Frenz, S.A. Downie, J. Massague and S.A. Newman, 1991. Role of transforming growth factor-beta in chondrogenic pattern formation in the embryonic limb: Stimulation of mesenchymal condensation and fibronectin gene expression by exogenenous TGF-beta and evidence for endogenous TGF-beta-like activity. Dev. Biol., 145: 99-109.
CrossRefPubMedDirect Link - Mackie, E.J., Y.A. Ahmed, L. Tatarczuch, K.S. Chen and M. Mirams, 2008. Endochondral ossification: How cartilage is converted into bone in the developing skeleton. Int. J. Biochem. Cell. Biol., 40: 46-62.
CrossRefPubMedDirect Link - Nakane, Y. and M. Tsudzuki, 1999. Development of the skeleton in Japanese quail embryos. Dev. Growth Differ., 41: 523-534.
CrossRefDirect Link - Pourlis, A.F., J. Antonopoulos and I.N. Magras, 2006. A light and electron microscopic study of the limb long bones perichondral ossification in the quail embryo (Coturnix coturnix Japonica). Ital. J. Anat. Embryol., 111: 159-170.
PubMed - Pourlis, A.F., I.N. Magras and D. Petridis, 1998. Ossification and growth rates of the limb long bones during the pre-hatching period in the quail (Coturnix coturnix Japonica). Anat. Histol. Embryol., 27: 61-63.
PubMed - Rivas, R. and F. Shapiro, 2002. Structural stages in the development of the long bones and epiphyses: A study in the New Zealand white rabbit. J. Bone Joint Surg. Am., 84-A: 85-100.
PubMedDirect Link - Roach, H.I. and J.R. Shearer, 1989. Cartilage resorption and endochondral bone formation during the development of long bones in chick embryos. Bone Miner., 6: 289-309.
CrossRefDirect Link - Sasano, Y., J.X. Zhu, M. Tsubota, I. Takahashi, K. Onodera, I. Mizoguchi and M. Kagayama, 2002. Gene expression of MMP8 and MMP13 during embryonic development of bone and cartilage in the rat mandible and hind limb. J. Histochem. Cytochem., 50: 325-332.
PubMedDirect Link - Simsa, S. and E.M. Ornan, 2007. Endochondral ossification process of the turkey (Meleagris gallopavo) during embryonic and juvenile development. Poult. Sci., 86: 565-571.
CrossRefDirect Link - Vortkamp, A., K. Lee, B. Lanske, G.V. Segre, H.M. Kronenberg and C.J. Tabin, 1996. Regulation of rate of cartilage differentiation by Indian hedgehog and PTH-related protein. Science, 273: 613-622.
CrossRefPubMedDirect Link - Yoshida, C.A., H. Yamamoto, T. Fujita, T. Furuichi and K. Ito et al., 2004. Runx2 and Runx3 are essential for chondrocyte maturation, and Runx2 regulates limb growth through induction of Indian hedgehog. Genes. Dev., 18: 952-963.
CrossRefDirect Link