
A.A. Megahed
Department of Plant pathology, National Research Centre, Dokki, Cairo, Egypt
Kh. A. El-Dougdoug
Department of Agriculture Microbiology, Faculty of Agriculture, Ain Shams University, Shoubra EL-Kheima, Cairo, Egypt
B.A. Othman
Department of Agriculture Microbiology, Faculty of Agriculture, Ain Shams University, Shoubra EL-Kheima, Cairo, Egypt
S.M. Lashin
Department of Plant pathology, National Research Centre, Dokki, Cairo, Egypt
M.A. Ibrahim
Department of Molecular Biology, Genetic Engineering and Biotechnology Division, National Research Centre, Dokki, Cairo, Egypt
A.R. Sofy
Department of Botany and Microbiology, Faculty of Science, Al-Azhar University, 11884 Nasr City, Cairo, Egypt
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 8 | Page No.: 385-390







ABSTRACT
The possibility of making use of the phenome non of Systemic Acquired Resistance (SAR) to control viruses achieved by the soaking treatment of tomato seeds cv. Castl Rock with three growth forms to Bacillus circulans, Pseudomonas fluorescens 2 and Trichoderma harzianum against Tomato mosaic tobamovirus (ToMV) infection. All the application forms of beneficial biotic inducers were reduced the mean number of ToMV local lesions on Datura metel. P. fluorescens 2 was found to be the best treatment in three forms on reduction of local lesion number 42.2, 32.7 and 38.1 of microbial liquid culture, microbial cells or spores and microbial culture filtrate forms, respectively, while the highest mean numbers of local lesions were 51.5, 61.7 and 73.5 of microbial liquid culture, m icrobial cells or spores and microbial culture filtrate, respectively for T. harzianum. The microbial culture filtrate form was more effective than other microbial forms to reduce mean number of ToMV local lesions to B. circulans, P. fluorescens 2 and T. harzianum isolates, 40.7, 32.1 and 51.5, respectively. The individual microbial isolates on all three microbial forms able to vary ToMV local lesions similarity (homologous or heterologous) and morphology (size center and surrounded with halo or without halo) compared with TMV mother strain.
PDF Abstract XML References Citation
Received: December 07, 2012;
Accepted: February 15, 2013;
Published: March 25, 2013
How to cite this article
A.A. Megahed, Kh. A. El-Dougdoug, B.A. Othman, S.M. Lashin, M.A. Ibrahim and A.R. Sofy, 2013. Induction of Resistance in Tomato Plants Against Tomato mosaic tobamovirus Using Beneficial Microbial Isolates. Pakistan Journal of Biological Sciences, 16: 385-390.
DOI: 10.3923/pjbs.2013.385.390
URL: https://scialert.net/abstract/?doi=pjbs.2013.385.390
DOI: 10.3923/pjbs.2013.385.390
URL: https://scialert.net/abstract/?doi=pjbs.2013.385.390
INTRODUCTION
Tomato (Lycopersicon esculantum Mil) is a principal vegetable crop evolving within Egypt. The loss of tomato production was attributed towards infection with different viruses. The erect of growth; decrease of yield and quality of tomato were observed under protective and open field agriculture. Application of culture filtrate from a non pigmented isolate of Erwinia herbicola infecting cucumber and tobacco plants for 12 h before inoculation inhibited the external symptoms due to CMV and TMV inoculation, respectively and resulted in growth increasing compared with untreated plants (Reiss et al., 1988). Aoki et al. (1993) found that, the culture filtrates of fungi including Fomes fomentarius and Schizophyllum commune have a highly active to induce systemic resistance against TMV infection on pepper and tomato plants. In the same time no toxicity was observed on the host plants. Maurhofer et al. (1994) found that the induction of systemic resistance of tobacco to Tobacco Necrosis Virus (TNV) by the root colonizing P. fluorescens strain CHA0 reduced the number of TNV lesions as well as, reduction of lesions diameter compared with non-colonized tobacco plants. Treated tomato plants with plant growth promoting rhizobacteria (PGPR) included Bacillus amyloliquefaciens 937a, B. subtillus 937b and B. pumilus SE34 as industrial formulated seed treatment under field conditions, reduced disease severity and incidence Tomato mottle virus (ToMoV). Further more, some of PGPR treatments increased tomato fruit yield at the first of two or more harvests (Murphy et al., 2000). Kolase and Sawant (2007) mentioned that, the culture filtrate of individual T. harzianum, T. viride and T. longisporum, as well as the combination of culture filtrate T. harzianum and T. viride were induced systemic resistance on tomato plants and reduced the TMV symptoms and TMV local lesions produced on Nicotiana glutinosa. The current study aims to induce systemic acquired resistance in tomato plants using bacterial and fungal liquid culture, cells or spores and cultural filtrates as antiviral against Tomato mosaic tobamovirus.
MATERIALS AND METHODS
Virus isolate: Tomato mosaic tobamovirus was obtained from Virology Lab. Agric. Microbiol. Dept. Fac. of Agric. Ain Shams Univ., Cairo, Egypt and then maintained in N. tabacum cv. White Burly. The virus isolate was confirmed by using the differential hosts (N. glutinosa, N. tabacum cv. White Burly, N. rustica, Chenopodium amaranticolor, D. metel, D. stramonium and L. esculantum cv. Castl Rock). The virus was propagated in N. tabacum cv. White Burly as a ToMV propagative host.
Source of biotic inducers: Two bacterial and one fungal isolates were obtained from Microbiological Resources Centre (MIRCEN), Fac. of Agric. Ain Shams Univ., Cairo, Egypt. The bacterial cultures were prepared via propagation of B. circulans on nutrient broth (Waksman, 1957), wherease P. fluorescens 2 were propagated in King's B broth (King et al., 1954). The suspensions of bacterial cells were adjusted at about mean density of 5x109 CFU mL-1 according to Raupach et al. (1996). The suspensions of bacterial cells were prepared according to Zehnder et al. (2000). The bacterial cells were collected from the prepared cultures by centrifugation at 6,000 g for 10 min and the collected pellets were resuspended in 0.02 M phosphate buffer, pH 7.0. The bacterial cells were washed twice with the same buffer and then adjusted at about mean density of 5x109 CFU mL-1. The culture filtrate of each bacterial isolate was prepared in specific broth medium according to Raupach et al. (1996) and then the culture was harvested by centrifugation at 1000 g for 10 min at 4°C. The supernatant was then filter sterilized through 0.045 μm Millipore filter to obtain sterilized culture filtrate (Patel and Patel, 1993).
T. harzianum was propagated in potato dextrose broth; the conidial media suspension was adjusted at about mean density of 1010 spores mL-1 according to Chambers and Scott (1995). The fungal spores were prepared by obtained conidial media suspension as described above, followed by centrifugation at 1000 g for 10 min, then the pellets were resuspended in 0.02 M phosphate buffer, pH 7.0 and mean density was adjusted to about 108 spores mL-1. The fungal culture filtrate was prepared according to Chambers and Scott (1995). After obtained the fungal cultures, cultures were harvested by passing them through four layers of muslin cloth and the filtrate centrifuged at 1000 g for 10 min at 4°C. The supernatant was filtered through Whatman No.1 filter paper and then passed through a 0.045 μm Millipore filter.
Greenhouse experiment: The soil used in the experiments was prepared by mixing clay, sand and peat-moss by ratio (3:1:1, v:v:v) and divided into two parts. The first was autoclaved at 121°C for one h, while the second was used as non sterilized soil. Plastic pots (20 cm in diameter) were washed and air dried, then sterilized with chlorox. The sterilized pots were filled with sterilized or non-sterilized soils at rate 1.5 kg pot-1. Three microbial isolates were used to induce the systemic resistance in tomato plants cv. Castl Rock against ToMV. The treatments were as follows: (seeds treated with B. circulans, P. fluorescens 2 and T. harzianum liquid cultures broth), (seeds treated with microbial cells and spores once) and (seeds treated with microbial cultural filtrates). ToMV was challenge-mechanical inoculated with ToMV inoculum onto adexial side of the first pair of tomato leaves, 2 weeks after planting to each treatment. Healthy control plants were inoculated with sterile 0.1 M potassium phosphate buffer (PPB), pH 7.2. The inoculated leaves were rinsed with water and kept under greenhouse conditions and observed until symptoms appeared after 21 days (Murphy et al., 2000).
Virus variability: Random sample 5 discs (5 cm) of inoculated tomato plants of each treatment were used for inoculation 4 indicator host (D. metel) at a same physiologically age. Each leaf of the indicator plants were mechanically inoculated with 50 μL of virus inoculum per leaf by a spatula, (Noordam, 1973). Diversity of single local lesion and number were used to determine the virus variability. Virus concentration was detected as mean number of L.L. by latin square method on D. metel as indicator host for ToMV.
RESULTS
Confirmation of Tomato mosaic tobamovirus isolate: Tomato mosaic tobamovirus was obtained from Virology Lab. Agric. Microbiol. Dept., Fac. of Agric. Ain Shams Univ., Cairo, Egypt and confirmed biologically by differential hosts which showed different symptom types. Local lesions; D. metel, D. stramonium, N. glutinosa, C. murale and C. amaranticolor produced necrotic local lesion while N. rustica and N. tabaccum cv. White Burly produced local lesion firstly and followed by systemic symptoms on inoculated leaves after 3-5 days post inoculation, (Table 1, Fig. 1). Systemic symptoms, L. esculantum cv. Castl Rock appeared mild mosaic.
 | |
| Fig. 1(a-g): | Leaves of differential hosts mechanically inoculated with ToMV isolate showing different symptoms, (a) N. tabaccum cv. White Burly, (b) C. amaranticolor, (c) N. rustica, (d) D. stramonium, (e) D. metel (f) C. murale and (g) N. glutinosa |
| Table 1: | Reaction of differential hosts with ToMV |
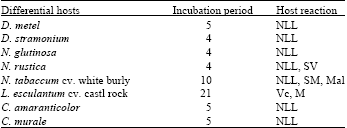 | |
| NLL: Necrotic local lesion, SM: Severe mosaic, Mal: Malformation, Vc: Vein clearing | |
Indication of host resistance: Three microbial isolates were used to induce the systemic resistance in tomato plants against Tomato mosaic virus (ToMV). These isolates were two bacterial isolates (B. circulans and P. fluorescens 2) and one fungal isolate (T. harzianum), they used as seed soaking by three forms, microbial liquid culture, microbial cells or spores and microbial culture filtrate under sterilized soil. The induced systemic resistance was detected biologically only by virus concentration and variability, mean number of ToMV local lesions on D. metel as ToMV indicator host. It was for determination of virus concentration, as well as ToMV variability was detected by L.L. diversity (Table 2, 3).
| Table 2: | Effect of biotic inducers on ToMV infectivity in tomato plants |
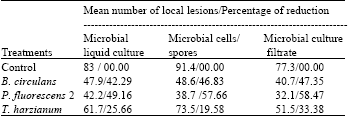 | |
All forms of biotic inducer treatments were reduced the mean number of L.L. compared with control. P. fluorescens 2 was found to be the best treatment among the used three forms on reduction of L.L. numbers 49.16, 57.66 and 58.47% of microbial liquid culture, microbial cells or spores and microbial culture filtrate, respectively. The microbial culture filtrate from the three microbial isolate forms has the highest percentage reduction of mean L.L number compared with the other forms of treatments. The lower percentage reduction of mean L.L numbers was 25.66, 19.58 and 33.38% of microbial liquid culture, microbial cells or spores and microbial culture filtrate, respectively for T. harzianum:
| Table 3: | Single local lesion diversity of ToMV inoculated tomato plants treated with individual biotic inducers on D. metel leaves |
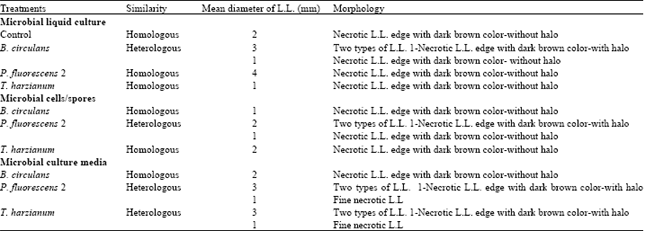 | |
ToMV isolate variability via local lesion diversity was estimated as frequency to use a parent strain of C ToMV in D. metel that gave conspicuous local lesions as homologous necrotic L.L. edge with dark brown color-without halo-2 mm in diameter. One then looks for effect of biotic inducers produced ToMV isolate concentration per single local lesion, similarity and characteristic morphology (center, surrounded halo, chlorotic, necrotic, size, fine and large) local lesion in D. metel. This method index of ISR in tomato according to severe of local lesions diversity (Fig. 2). The virus inoculated plants treated with individual biotic inducers (Table 2), (B. circulans, P. fluorescens 2 and T. harzianum) revealed different local lesions (Fig. 2).
In the microbial liquid culture form, B. circulans induced two types of heterologous local lesions, necrotic edge with dark brown color, surrounded with halo, 3 mm in diameter. The second were 2 mm in diameter of necrotic local lesions with dark brown edge, without halo, (Fig. 2-a1). On the other hand, B. circulans induced homologous necrotic local lesions with dark brown edge, without halo 1 mm in diameter, in microbial cells or spores form, (Fig. 2-b1). In microbial culture filtrate B. circulans induce 2 mm in diameter of necrotic local lesions wit h dark brown edge, without halo, (Fig. 2-a1). P. fluorescens 2 was induced homologous necrotic local, 4 mm in diameter, surrounded without halo, the edge with dark brown color in the microbial liquid culture form (Fig. 2-a2). In microbial cells or spores form, P. fluorescens 2 induced heterologous local lesions. Type one were necrotic local lesions, edge with dark brown color, without halo and it has 3 mm in diameter. The second were 2 mm in diameter of necrotic local lesions without halo, surrounded with dark brown edge, (Fig. 2b2). P. fluorescens 2 produced heterologous local lesion, the first type 1 mm in diameter necrotic local lesions and its edge with dark brown color, when have not halo. The second one were necrotic local lesions, 3 mm in diameter, with dark brown edge, without halo, in microbial culture filtrate (Fig. 2-b3).
In the microbial liquid culture form T. harzianum induced homologous necrotic local lesions, 1 mm in diameter, surrounded without halo, the edge with dark brown color, (Fig. 2-c1), while it was induced 2 mm in diameter necrotic local lesions with dark brown edge, without halo, in microbial cells or spores form, (Fig. 2-c2). On the other microbial culture filtrate, two types of heterologous local lesions, necrotic edge with dark brown color, without halo, 1 mm in diameter were induced by T. harzianum. The second were 3 mm in diameter of necrotic local lesions with dark brown edge, surrounded with halo, (Fig. 2-c3).
DISCUSSION
The obtained results of ToMV concentration revealed that, three forms of biotic inducer treatments were reduced the mean number of L.L. compared with control. P. fluorescens 2 was found to be the best treatment in three forms on reduction of L.L. numbers 42.2, 32.7 and 38.1 of microbial liquid culture, microbial cells or spores and microbial culture filtrate, respectively. The microbial culture filtrate from the three microbial isolate forms has the lowest mean number of L.L compared with the other forms treatments. The highest mean numbers of L.L were 51.5, 61.7 and 73.5 of microbial liquid culture, microbial cells or spores and microbial culture filtrate, respectively for T. harzianum. The same results were obtained by Zhang et al. (2002) and Zhao and Wu (2007).
 | |
| Fig. 2: | Single local lesion diversity of ToMV inoculated tomato plants treated with individual biotic inducers on D. metel leaves, (c) Control, (a1) B. circulans liquid culture, (a2) B. circulans cells, (a3) B. circulans culture filtrate, (b1) P. fluorescens 2 liquid culture, (b2) P. fluorescens 2 cells, (b3) P. fluorescens 2 culture filtrate, (c1) T. harzianum liquid culture, (c2) T. harzianum spores, (c3) T. harzianum culture filtrate |
Also, the lower percentage reduction of mean L.L numbers were due to the T. harzianum treatment as 25.66, 19.58 and 33.38% of microbial liquid culture, microbial cells or spores and microbial culture filtrate, respectively for. The same results were obtained by Zhang et al. (2002) and Zhao and Wu (2007). ToMV isolate variability via local lesion diversity was estimated as frequency to use a parent strain of ToMV in D. metel by ToMV concentration per single local lesion, similarity and characteristic morphology (center, surrounded halo, chlorotic, necrotic, size, fine and large) of local lesion. Induced mutants are not a random selection of all possible mutants for several reasons: (1) the initial recognition of mutants is based on a difference in symptoms produced on a few selected hosts. (2) The mutants selected are generally those that produced intact virus particles in reasonable yield. (3) Only mutations in the coat protein cistron can be examined by chemical methods at the present time. (4) Silent mutations can occur in which there is a change in a base in the RNA, (Matthews, 1970).
The obtained results showed that, all individual biotic treatments were able to vary local lesion similarity (homologous or heterologous) morphology (size center, surrounded with halo or without halo) produced to ToMV infection. These results are in accordance to Koganezawa et al. (1998) and Zhang et al. (2002). The size of TMV local lesions on Xanthi-nc tobacco plants reduced by approximately 50% compared with control plants as a result to induce systemic resistance, (Vernooij et al., 1995), these results in accordance to obtained results.
The SAR was indicated by virus variability (local lesion diversity) produced by ToMV for individual of microbial isolates. The obtained results showed that, all individual biotic treatments were able to vary local lesion similarity (homologous or heterologous) morphology (size center, surrounded with halo or without halo) produced to ToMV infection. These results revealed that biotic inducers under study have an effect on chemical and physiological changes in plant which duo to virus variability based on L.L. diversity.
ACKNOWLEDGMENT
Our special thanks, for all staff member of Virology Lab. Agric. Microbiol. Dept. and Microbiological Resources Centre (MIRCEN), Fac. of Agric. Ain Shams Univ., Cairo, Egypt, for their kindly helping and supporting by Tomato mosaic tobamovirus isolate and microbial isolates, respectively. Also many thanks to National Research Centre (NRC), Dokki, Cairo, Egypt for providing the fund for my research.
REFERENCES
- Aoki, M., M. Tan, A. Fukushima, T. Hieda and S. Kubo et al., 1993. Antiviral substances with systemic effects produced by basidiomycetes such as Fomes fomentarius. Biosci. Biotechnol. Biochem., 57: 278-282.
Direct Link - King, E.O., M.K. Ward and D.E. Raney, 1954. Two simple media for the demonstration of pyocyanin and fluorescin. Transl. Res., 44: 301-307.
PubMedDirect Link - Koganezawa, H., T. Sato and T. Sasaya, 1998. Effects of probenazole and saccharin on symptom appearance of tobacco mosaic virus in tobacco. Ann. Phytopathol. Soc. Japan, 64: 80-84.
Direct Link - Maurhofer, M., C. Hase, P. Meuwly, J.P. Metaraux and G. Defago, 1994. Induction of systemic resistance of Tobacco necrosis virus by the root-colonizing Pseudomonas fluorescens CHAO: Influence of the gacA gene of pyoverdine production. Phytopathology, 84: 139-146.
Direct Link - Murphy, J.F., G.W. Zehnder, D.J. Schuster, E.J. Sikora, J.E. Polston and J.W. Kloepper, 2000. Plant growth-promoting rhizobacterial mediated protection in tomato against Tomato mottle virus. Plant Dis., 84: 779-784.
CrossRefDirect Link - Patel, H.R. and B.N. Patel, 1993. Inhibition of chlorotic mottle virus infection by plant extracts and other substances. Indian J. Mycol. Plant Pathol., 23: 191-193.
Direct Link - Raupach, G.S., L. Liu, J.F. Murphy, S. Tuzun and J.W. Kloepper, 1996. Induced systemic resistance in cucumber and tomato against Cucumber mosaic cucmovirus using plant growth-promoting rhizobacteria (PGPR). Plant Dis., 80: 891-894.
Direct Link - Reiss, E., I. Weber and W.D. Ostermann, 1988. A bacterial culture filtrate with antiphytoviral activity on cucumber against cucumber mosaic virus and tobacco mosaic virus. Archiv Phytopathol. Pflanzenschutz, 24: 353-355.
Direct Link - Vernooij, B., L. Friedrich, P.A. Goy, T. Staub, H. Kessmann and J. Ryals, 1995. 2,6-Dichloroisonicotinic acid-induced resistance to pathogens without the accumulation of salicylic acid. Mol. Plant-Microbe Interact., 8: 228-234.
Direct Link - Zehnder, G.W., C. Yao, J.F. Murphy, E.R. Sikora and J.W. Kloepper, 2000. Induction of resistance in tomato against Cucumber mosaic cucumovirus by plant growth-promoting rhizobacteria. BioControl, 45: 127-137.
Direct Link