
C. C. Ogueke
Department of Food Science and Technology, Federal University of Technology,P.M.B. 1526 Owerri, Nigeria
J.N. Nwosu
Department of Food Science and Technology, Federal University of Technology,P.M.B. 1526 Owerri, Nigeria
C.I. Owuamanam
Department of Food Science and Technology, Federal University of Technology,P.M.B. 1526 Owerri, Nigeria
J.N. Iwouno
Department of Food Science and Technology, Federal University of Technology,P.M.B. 1526 Owerri, Nigeria
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 10 | Page No.: 489-496







ABSTRACT
Ugba is the Ibo name of the fermented African Oilbean seeds (Pentaclethra macrophylla, Benth). It is a traditional food condiment generally produced by natural (local) fermentation in homes as a small family business. It is an important and cheap source of protein for people whose staple foods are deficient in proteins. It is also eaten as a delicacy and used as flavouring for soup. This write up aims to review all published studies on ugba in the direction of the various methods used in the production, the chemical composition of the seeds, the microorganisms involved and the biochemical changes that occur during fermentation and optimization of the fermentation. The nutritional and food values, toxicological properties, health promoting potentials, microbiological safety as well as the storage and preservation have also been highlighted.
PDF Abstract XML References Citation
Received: February 13, 2010;
Accepted: May 04, 2010;
Published: June 23, 2010
How to cite this article
C. C. Ogueke, J.N. Nwosu, C.I. Owuamanam and J.N. Iwouno, 2010. Ugba, the Fermented African Oilbean Seeds; its Production, Chemical Composition, Preservation, Safety and Health Benefits. Pakistan Journal of Biological Sciences, 13: 489-496.
DOI: 10.3923/pjbs.2010.489.496
URL: https://scialert.net/abstract/?doi=pjbs.2010.489.496
DOI: 10.3923/pjbs.2010.489.496
URL: https://scialert.net/abstract/?doi=pjbs.2010.489.496
INTRODUCTION
Ugba is the Igbo name for the fermented African OIlbean seeds (Pentaclethra macrophylla, Benth). It is called Ukana by the Efiks in Southern Nigeria. It is consumed by an estimated 15 million people in Eastern Nigeria, majority of whom are Igbos (Odunfa and Oyeyiola, 1985). It is a traditional food generally prepared in homes as a small family business. The method of production varies from one producer to another resulting in a non-uniform product (Njoku and Okemadu, 1989). The beans that have been fermented for more than three days are taken as a delicacy. Well fermented beans are added to soup as flavouring (Odunfa and Oyeyiola, 1985). It is widely consumed in eastern states of Nigeria with tapioca, stock fish and garden eggs and leaves. It can also be eaten with bitter kola (Garcinia kola) or kola nuts (Cola acuminate and C. nitida) and when prepared with gardenegg leaves is used to eat yam and cocoyam (Okafor et al., 1991; Mbajunwa et al., 1998). It is an important and cheap source of protein for people whose staple foods are deficient in proteins (Obeta, 1983). The quantity of ugba produced annually is not known, since the seeds are collected by individuals and sold in the market to ugba producers.
Nature of the plant and the seeds: Oilbean seeds for ugba production are obtained from a perennial legume tree, Pentaclethra macrophylla, Bentham, commonly called the oilbean tree. The trees are often planted along the sides of roads as shade trees and around communities as cash crops. The fruit is a black, hard and woody pod measuring about 35-36 cm long and 5-10 cm broad. When mature it splits open explosively to release about eight flat, glossy brown seeds measuring about 5-7 cm in diameter and weighing between 15-20 g (Keay et al., 1964; Odunfa, 1986a).
Chemical composition of seeds: The oilbean seeds contain 4-17% carbohydrate, 44-47% oil which has been found to be rich in oleic acid (Nwokedi, 1975; Odoemelam, 2005) and linoleic acid (Onwuliri et al., 2004). Onwuliri et al. (2004) also found out that the saturated fatty acid, lignoceric acid, occurred in high amounts constituting about 10% of the total fatty acid concentration. Some workers said that the oil content could be as low as 38% (Kar and Okechukwu, 1978). They also reported that the oil contains about 75% saturated fatty acids and 25% unsaturated fatty acids. Table 1 (Achinewhu, 1983) shows the fatty acid content of the seeds. Both saturated and unsaturated fatty acids are found in the seeds. For the saturated fatty acids, lignoceric acid appears to be present in the largest amount constituting about 12% while palmitic acid is the least with 3.4%. Behemic acid is also present with 5.2%. The major unsaturated fatty acid in the seeds is linoleic acid constituting 42.8%. Oleic acid is also present in appreciable amounts (29.0%).
| Table 1: | Fatty acid composition of African oilbean seeds*a |
 | |
| *As percentage of total oil. aAchinewhu (1983) | |
Linolenic and gadoleic acids are present in very small amounts (3.2 and 0.28%, respectively). The presence of appreciable amounts of behenic and lignoceric acids is not desirable for edible oils (Odunfa, 1986a).
However, Odoemelam (2005) believes that the high degree of unsaturation makes it suitable for cooking purposes and for use as a drying oil for cosmetics, paints and varnishes.
Also they have been found to contain 36.2-43.89% crude protein which contains the 20 essential amino acids. However, the sulphur containing amino acid content is much lower than those found in other plant proteins (Mbadiwe, 1978; Mba et al., 1974; Odoemelam, 2005). The high content of other essential amino acids makes the seeds a potential source of protein (Achinewhu, 1982). Table 2 shows the amino acid profile of the seeds. Glutamic acid appears to be the largest amino acid contained in the seeds. This may be responsible for its use as a flavouring for soups in south eastern Nigeria. Aspartic acid, lysine and phenylalanine are also present in appreciable amounts in the seeds.
Preparation of ugba: Methods for ugba preparation vary from one community to the other. In this method described by Obeta (1983), the seeds are boiled in water for 16-18 h to remove the tough testa. The cotyledons are then sliced, boiled again for 30 min and left overnight in water at room temperature. The sliced cotyledons are then washed in water and packaged in leaves of banana.
Another method described by Odunfa and Oyeyiola (1985) and Odunfa (1986a) shows that the seeds are boiled in water over an open fire for 4-5 h or even up to 12 h. The cotyledons are then removed from the seed coats and washed. The cotyledons are again boiled overnight over a low flame, allowed to cool, drained and washed several times to remove bitter components in the cotyledons and soaked for a period of 6 h. The cotyledons are then cut into long thin slices which are mixed with salt, put in a clean pot, covered and fermented for up to 5 days at room temperature. Usually after 2-3 days of fermentation the sliced cotyledons are wrapped in banana leaves and tied tightly.
| Table 2: | Amino acid content (g/100 g protein) of African oilbean seedsb,c |
 | |
| bMba et al. (1974), cAchinewhu (1982) | |
Njoku and Okemadu (1989) also described another production method. The seeds are boiled for 5-8 h, after which the hard shells are removed. The cotyledons are cooled, washed and sliced into 4-5x0.1-0.2 cm slices. These are washed again and boiled for another 1-2 h, cooled and soaked in water for about 10-12 h. They are washed and allowed to drain for ½-1 h. in a basket lined with banana leaves (Musa sapientum linn). They are then wrapped about 40-50 g of slices using another leaf (Mallotus oppositifolius) and incubated for 72 h at room temperature.
Another method has been described by Sokari and Wachukwu (1997). These workers said toasting the bean seeds in hot (100°C) sand and holding for a further 30 min at 100°C significantly improved dehulling. They also said that slicing to 1 mm, boiling for 30 min and soaking for 2 h removed the bitter taste associated with the seeds. They claimed that the technique reduced the general production time by 2 days and the quality of ugba produced from this process was the same as that produced from the rather more cumbersome and time-consuming traditional technique.
The differences in the various processing methods are responsible for the variations in the products from one community to the other. The wrapped ugba at different stages of fermentation are sold to consumers and they are often told the length of fermentation at the time of purchase.
Microorganisms involved in the fermentation: Several workers have investigated the microorganisms involved in the fermentation (Table 3).
Only bacteria are involved in the fermentation (Obeta, 1983; Odunfa and Oyeyiola, 1985; Ejiofor et al., 1987; Ogueke and Aririatu, 2004). The main fermenting microorganisms have been identified to be proteolytic Bacillus sp. (Obeta, 1983) which include B. subtilis (most predominant), B. licheniformis, B. megaterium, B. macerans and B. circulans.
| Table 3: | Succession of the major microorganisms encountered in fermentation of sliced oilbean seeds (per gram)e |
 | |
| eObeta (1983) | |
Their numbers increased tremendously from 103 at the start of fermentation to 108 at the end of the fermentation (72 h). Other bacteria identified in the fermenting slices include coagulase negative Staphylococcus sp., Micococcus sp. (their numbers decreased after 72 h of fermentation), Leuconostoc mesenteroides; Lactobacillus plantarum, Streptococcus lactis, Proteus sp., Enterobacter sp. and E. coli. Some workers isolated the yeasts Candida tropicalis and Geotrichum candidum during fermentation (Ejiofor et al., 1987).
Since protein hydrolysis is the major biochemical change in ugba fermentation (Oyeyiola, 1981), it can be assumed that the Bacillus sp. are the main fermenting organisms. They were found to persist until the end of the fermentation and their numbers increased throughout the period of fermentation while the numbers of others decreased after 24 h of fermentation (Obeta, 1983). Also Bacillus sp. are important sources of proteases (Fogarty and Griffin, 1973). The other bacteria only managed to grow on the little carbohydrate present in the seeds, majority of which may have been lost due to leaching during the preparatory stages (Ruiz-Teran and Owens, 1999). The disappearance of these bacteria could also be due to the activities of B. subtilis, the predominant bacterium in the fermentation, which is known to produce the antibiotic bacitracin. The antibiotic may have inhibited the growth of these bacteria and the disappearance of Micrococcus sp. especially at 96 h of fermentation is believed to be due to this (Ogueke and Aririatu, 2004). Micrococcus sp. are very sensitive to bacitracin (British Pharmacopoeia Commission, 1993).
Since the bean seeds were boiled for hours before fermentation the microorganisms involved in the fermentation could not have originated from the beans. The bacteria involved in the fermentation probably were introduced through the air, water, utensils, leaves used in wrapping or by handling during the preparatory stages (Obeta, 1983; Odunfa and Oyeyiola, 1985). Example Staphylococcus sp. are more commonly associated with the skin and hence are easily disseminated through handling. Also addition of salt would selectively favour the growth of Staphylococcus and Micrococcus sp. which are known to be salt tolerant (Adam and Moss, 1999).
Changes that occur during fermentation: Various biochemical changes occur during the fermentation. Obeta (1983) found that pH increased from 6.5 at 0 h to 9.0 at 48 h and declined to 7.1 at 72 h. The rise in pH has been attributed to the abundant production of ammonia during the fermentation due to protein hydrolysis and deaminase activity. The increase in pH would encourage the growth of Bacillus sp. which have been found to grow well at pH 7.0 to 8.0 (Odunfa and Oyeyiola, 1985). The drop in pH to 7.1 at 72 h could be attributed to the fact that B. subtilis and B. licheniformis use ammonia as nitrogen source (Odunfa, 1986b). However, Odunfa and Oyeyiola (1985), Njoku and Okemadu (1989) and Ogueke and Aririatu (2004) found that pH rose throughout the fermentation from 5.0-5.7 at 0 h to 7.9-8.7 after 3-5 days of fermentation.
The temperature of fermentation was observed to increase from about 30.8 to 34.5-38.5°C within the first 24-36 h of fermentation and decreased gradually afterwards to 30-32.5°C at the end of fermentation (Odunfa and Oyeyiola, 1985; Njoku and Okemadu, 1989). Thus ugba fermentation is exothermic. This initial increase in temperature has been attributed to the intense metabolic activities of the microorganisms (period of maximum microbial activity) and represents the most active and important period of the fermentation. This is because enzyme studies (Njoku and Okemadu, 1989) have revealed that the α-amylase, proteolytic and lipolytic enzyme activities attained their maximum levels at 24-36 h of fermentation. Thus it could be the enzymes already produced rather than the presence of the microorganisms that continued the fermentation later.
Moisture content was also found to increase throughout the period of fermentation (52-56.90% to 71.20-73%) (Odunfa and Oyeyiola, 1985; Njoku and Okemadu, 1989; Ogueke and Aririatu, 2004). The increase in moisture is believed to be due to the hydrolytic activities of the microorganisms. However, the high moisture level has been suggested to predispose the product to rapid spoilage (Odunfa and Oyeyiola, 1985; Ogueke and Aririatu, 2004).
Njoku and Okemadu (1989) detected α-amylase, proteolytic and lipolytic enzymes from the start of ugba fermentation. These enzymes attained their maximum levels at 24-36 h. They suggested that this could be assumed to be the period of maximum microbial activity. The initial enzyme activity detected could be due to the activity of the natural microflora of the oilbean which developed particularly during the soaking of the cooked beans. Njoku and Okemadu (1989) therefore suggested that it could well be that fermentation began much earlier during the soaking of the sliced beans. Some workers (Enujiugha et al., 2002, 2004) have demonstrated that the raw seeds contain both α-amylase and lipase. They observed that the specific activity of the purified α-amylase from the raw and fermented seeds were 0.037 mL-1 min-1 and 0.88 mL-1 min-1, respectively. They also claimed that these enzymes complement the bacterial enzymes during fermentation. However, since the seeds were boiled for several hours before fermentation they could not have contributed to the fermentation as the boiling must have inactivated them. The proteinase enzyme is considered the most important enzyme in ugba fermentation. Njoku and Okemadu (1989) detected a sevenfold increase in the level of amino nitrogen while Enujiugha (2003) also observed a steady increase in the level of amino nitrogen from 1.23 to 13.68 mg Ng-1 DM after 72 h of fermentation. Only a two fold increase in reducing sugar was found, while the activity of lipase was minimal compared to the other two enzymes (Njoku and Okemadu, 1989). This agrees with the report of Achinewhu (1986) that fermentation has no appreciable effect on the fatty acid content of P. macrophylla. The minimal activity of the lipase could be attributed to the effect of NaCl that is usually added during fermentation. Enujiugha et al. (2004) in their study of the lipase activity in dormant seeds of African oilbean seeds observed that the activity of lipase isolated from the seeds were inhibited up to 36% by NaCl. However, they found that presence of Ca2+ increased the activity of the enzyme by 64%.
Optimization of ugba fermentation: Several workers have conducted studies on ways of optimizing the production of Ugba. Most of the studies have been on the modification of some parameters that affect the fermentation such as temperature and Relative Humidity (RH), the use of starter cultures and immobilized cells in the fermentation process. Isu and Njoku (1998) the influence of temperature, relative humidity and microenvironment on the natural fermentation of Ugba. Their results suggested that fermentation may have been faster at 80% relative humidity. They suggested that the traditional method of wrapping with leaves of Mallotus oppositifolium presumably produced warmth and a humid environment while limiting the accessibility of air to the fermenting substrate. Thus the 80% RH created a more humid environment than the atmospheric RH of 74.5% which was more favourable for the fermentation process. They also found that a temperature of 35°C was most suitable for the fermentation generating a peak amino-nitrogen content of 19.6 mg N/100 g dry matter within 48 h. Thus reducing fermentation time by 24 h. as against the 72-96 h. in the traditional fermentation process.
The use of immobilized cells of Bacillus sp. in the fermentation process has also been carried out by some workers. Isu and Ofuya (2000) studied the use of pure cultures of Bacillus subtilis attached to cowpea and maize granules in the fermentation process. These workers monitored changes in pH, amino-nitrogen and protease activity which they said were the fermentation indicators. They found out that in comparison with the natural fermentation changes in these indicators were more pronounced in the fermentation carried out with the immobilized cells (Table 4). For example the protease activity increased from 4.5 to 27.65 mg N/min for the immobilized cells as against 10.5 mg N/min produced by the natural fermentation.
The use of the immobilized cells resulted in the reduction of the fermentation time to 48 h. as compared to 96 h. for the natural fermentation process. The authors attributed the increased activity observed with the immobilized cells to the increase in cell density per unit reactor and enhanced cell wall permeability and metabolism. Their sensory evaluation studies also showed that the products from immobilized cell fermentation were well accepted. The cultures were also stable and viable for 6 months on the granules of cowpea.
The use of starter cultures of B. subtilis and the spores in association with cowpea granules were studied by Isu and Abu (2000). They observed that the viability of the cells in association remained stable at 94.5% for 6 months at 30°C and up to 10 months at 4°C while the viability of the spores in association remained at 96% for up to 10 months at both 4 and 30°C. They also observed that the indicators of fermentation were more pronounced than in the natural fermentation and fermentation was completed within 48 h.
The use of such cultures for fermentations can be applied to the indigenous technologies of the developing countries. This type of starter culture will enhance process standardization and uniform product quality (Isu and Ofuya, 2000). It will also eliminate the chances of contamination by potential food poisoning and other disease causing and spoilage microorganisms.
| Table 4: | Effect of different starter cultures on ugba fermentation variables |
 | |
| aIncrease within 48 h; bIncrease within 72 h; cIncrease within 96 h | |
Nutritional value of ugba: Table 5 shows the mineral and vitamin content of the seeds. The vitamin content of the seeds is low while they are poor sources of calcium and phosphorus (Duke, 1981). Odoemelam (2005) has also shown that the seeds contain sodium (236.2 ppm) and potassium (181.3 ppm). The contents of niacin and riboflavin have been found to decrease during fermentation. Mineral content also decreased during fermentation while no phosphorus could be found in ugba (Duke, 1981).
However, since ugba is usually eaten with fish or added as a condiment to soup containing animal proteins, much of the needed calcium and protein may be obtained from these sources (Odunfa, 1986a).
The major sugars found in the seeds are stachyose, galactose and fructose while saponins constitute about 2.1% of the seeds (Achinewhu, 1983). These saponins when hydrolysed would yield glucose, arabinose, rhaminose, oleanolic acid and hederagenin. The content of these carbohydrate decreased significantly as fermentation time increased (Monago et al., 2004).
However, Enujiugha (2003) has shown that fermentation for 72 h slightly increased the crude protein and ash contents of ugba. The amino nitrogen increased steadily from 1.23 mg N g-1 DM prior to fermentation to 13.68 mg N g-1 DM after 72 h of fermentation. He also found that the principal fatty acid linoleic acid increased from 60.68 to 67.57% of the total fatty acids while oleic acid decreased from 26.95 to 22.59%. Palmitic acid and other saturated fatty acids in the seed oil were also slightly affected by the fermentation. However, Onwuliri et al. (2004) found that fatty acid concentrations did not change appreciably with processing and fermentation. There was also accumulation of formic acid, acetic, lactic and butyric acids and got to 0.20, 0.18, 0.35 and 0.41 mg g-1 respectively after 72 h of fermentation.
| Table 5: | Mineral and vitamin content of unfermented and fermented ugbad |
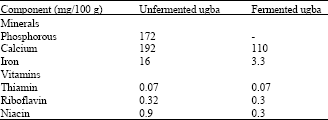 | |
| dDuke (1981) | |
Isichei and Achinewhu (1988) studied the nutritive value of African oil bean seeds. The seeds were high in energy with a slight difference between the gross energy value of unfermented and fermented oil bean seeds. They also stated that the results obtained from the estimated protein energy ratio (pe%) and net dietary protein calorie percent (NDpCal%) showed that the two processed forms of the seed have the potential to satisfy human protein and energy requirements.
Feeding of rats with unfermented (UOB) and the fermented (FOB) seeds resulted in weight loss (-0.82 g and -0.11 g, respectively) (Table 6). However, the average daily intake by the rats was higher for the fermented (5.06 g) than the unfermented (4.72 g). The unfermented and fermented seeds produced a negative protein efficiency ratio (PER) in rats. The protein digestibility was also low. Although, the oil bean seeds are rich in protein (Achinewhu, 1982), they suffer source nutritional drawback as they could not promote nor maintain growth of rats. The poor performance has been attributed to the presence of toxic components in the seeds which impair protein utilization (Isichei and Achinewhu (1988). Mbadiwe (1978) attributed the poor nutritional quality to the presence of growth-depressing factors. Onwuliri et al. (2004) have shown that the seeds contain some anti-nutritional factors which included cyanide, phytate, tannin and oxalate. The raw seeds were found to contain the highest concentrations of all the anti-nutritional factors except oxalate with the highest concentration (937.5 mg/100 g) in the boiled seeds. However, they observed a progressive reduction in the level of all the anti-nutritional factors at the different stages of processing and fermentation. The fully fermented ugba had a reduction of 73.49% for cyanide, 79.41% for tannin, 76.92% for oxalate and 45.98% for phytate. Akindahunsi (2004) studied the effect of salting, soaking before cooking and fermentation on the proximate, anti nutritional and mineral content of the bean seeds. They significantly decreased protein content by 10.5, 9.9 and 8.0%, respectively. However, the energy levels increased from 312.5 kcal mol-1 in raw seeds to 450.9, 440.5 and 405.9 kcal mol-1, respectively after treatments. The zinc levels increased while Mg, Na and K levels decreased.
Toxicology of ugba: The unfermented oil bean seeds contain a number of anti nutritional and toxic factors.
| Table 6: | Weight gain, feed intake, PER feed efficiency ratio and digestibility of test samples fed to the rats for 28 days*f |
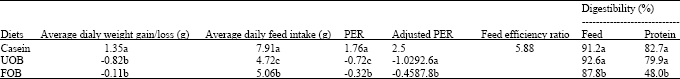 | |
| UOB: Unfermented oil bean. FOB: Fermented oil bean. *Only means followed by different letters within columns differ significantly (p<0.05). fIsichei and Achinewhu (1988) | |
Achinewhu (1983), showed the presence of saponins while Duke (1981) reported the presence of a poisonous alkaloid, paucine in the oilbean seeds. Mbadiwe (1979) reported the presence of caffeoylputrescine, a growth depressant. However, hemagglutinnins were not found in the oil bean seeds (Toms and Western, 1971). The presence or absence of these toxic substances in the fermented beans has not been investigated. It is, however, believed that these substances are eliminated during the processing and fermentation of the seeds, especially during the soaking, where they can leach out into the water used for soaking. Other anti-nutritional factors in the beans have been shown to reduce progressively during processing and fermentation (Onwuliri et al., 2004). Ruiz-Teran and Owens (1999) have also shown that such substances are leached out during soaking in soya bean tempeh production. However, Akindahunsi (2004). observed that salting and soaking before cooking and fermentation did not have any effect on the level of tannins while the level of phytate increased. Thus these may be responsible for the poor performance of the bean seeds during feeding studies in rats by Isichei and Achinewhu (1988). However, there has not been any reported case of health problems resulting from the consumption of ugba over the years.
Although saponins have been reported to be toxic, they may be beneficial since they have been found to lower plasma cholesterol. Monago et al. (2004) have shown that ugba fermented for up to four days decreased the level of plasma cholesterol in rats, the rate of decrease increasing with the time (days) of fermentation. Thus consumption of the well fermented product promotes health. Chidozie (2006) has shown that administration of the fermented seeds as a food supplement have greatly reduced the risk of cancer and some tobacco related diseases and cancer patients who had regular fermented oil bean seeds as food supplement showed marked improvements in regaining quality health.
Flavour components of ugba: Fermented African oilbean seeds have typical aroma and flavour. These are due to the various volatile compounds produced by the fermenting microorganisms in the course of their metabolism. Not much work has been done in this direction. However, Kabuo et al. (2007) studied the various flavour and aroma components present in the beans fermented with pure cultures of microorganisms isolated from ugba. The sample fermented with B. subtilis and B. licheniformis were found to produce the best ugba with its typical aroma and flavour. The compounds identified were ethyl state (3.60%), ethyl oleate (4.70%), ethyl linoleate (14.14%), ethyl phenol (6.94%) and ethyl phenol (6.94%) and ethyl octanoate (2.69%). The control (naturally fermented) contained ethyl benzoate (18.40%), ethyl carbonate (5.557%), methyl pentanone (1.67%) and ethyl octanoate (4.72%).
Packaging of ugba: Attempts have been made by some workers to effectively package the product and thus extend the shelf life of the product. Ogbulie et al. (1998) made attempts to package the product in low and high density polyethylene sachets and aluminium foil wraps as well as treatment with chemical preservatives such as 2% sodium chloride. However, none of the methods could extend the shelf life beyond 8 days. Mbata and Orji (2008) in their study applied a process of pasteurization at a temperature of 98-100°C for 30 min, which they said completely eliminated all the organisms present including the organisms used for the fermentation. This was able to extend the shelf life to 8 days. They also made attempts to package them in returnable and sterilizable bottles/cups. The containers were sterilized before use and the products pasteurized in the containers. These were able to keep for six weeks. The colour, taste, aroma, softness and other physicochemical properties of the product before and after keeping for six weeks compared favourably well with the locally produced ugba.
Enujiugha and Akanbi (2008) used conventional batch retort procedures. The sliced and fermented beans were canned in three different media (brine, refined ground oil and tomato sauce). The product was able to keep for 6 months at ambient temperature storage. Total viable counts after 6 months of storage were 9.3x103, 1.7x104, and 6.0x103 cfu g-1 in brine, refined groundnut oil and tomato sauce respectively while the free fatty acids content (g oleic acid) were 3.12, 2.54 and 3.98 respectively. The peroxide values obtained after storage were 11.63, 9.54 and 10.02 meq kg-1, respectively while the acid values (mg NaOH g-1 oil) were 6.43, 5.10 and 7.92, respectively. Sensory evaluation of the canned products showed that the groundnut-oil canned product was least acceptable in terms of aroma and overall acceptability although all the products showed increased softening and colour darkening with the prolonged storage.
Microbiological safety of ugba: Various microbiological studies conducted on ugba (Obeta, 1983; Odunfa and Oyeyiola, 1985; Ogueke and Aririatu, 2004) showed that food pathogens such as Clostridum perfringens, C. botulinum, Salmonella sp., Shigella sp. and Vibrio sp. have not been isolated from ugba. However, such bacteria as E. coli and Staphylococcus aureus have been isolated. These are bacteria capable of causing food infections/poisoning. But since the preparation of the delicacy or addition as condiment to soup involves heating, they will be eliminated during the process. Azubuine and Isu (2006) studied the fungal contamination of the fermenting product. They isolated Aspergillus flavus, A. niger, Penicillium chrysogenum and Fusarium sp. This poses a serious health risk as these are moulds that produce mycotoxins in foods. This calls for the observation of Good Manufacturing Practice (GMP) during the production. However, application of starter cultures and immobilized cells in the fermentation process will eliminate these possibilities of contamination with unwanted organisms.
However, their numbers decreased with increase in the number of days of fermentation. Thus the environment was not suitable for their growth and toxin production, especially with the increasing pH of the fermenting slices into the alkaline region.
REFERENCES
- Achinewhu, S.C., 1982. Composition and food potential of African oil bean (Pentaclethra macrophylla) and velvet bean (Mucuna uriens). J. Food Sci., 47: 1736-1737.
CrossRefDirect Link - Achinewhu, S.C., 1983. Protein quality of African oil bean seed (Pentaclethra macrophylla Benth). J. Food Sci., 48: 1374-1375.
CrossRef - Achinewhu, S.C., 1986. The effect of fermentation on carbohydrate and fatty acid composition of African oil bean seed (Pentaclethra macrophylla). Food Chem., 19: 105-116.
CrossRef - Akindahunsi, A.A., 2004. Physicochemical studies on African oilbean (Pentaclethra macrophylla) seed. J. Food Agric. Environ., 2: 14-17.
Direct Link - Duke, J.A., 1981. Handbook of Legumes of World Economic Importance. 1st Edn., Springer, New York, ISBN: 978-1-4684-8151-8, Pages: 346.
CrossRefDirect Link - Enujiugha, V., C. Amadi and T. Sanni, 2002. Amylase in raw and fermented African oilbean seed (Pentaclethra macrophylla Benth). J. Eur. Food Res. Technol., 214: 497-500.
CrossRef - Enujiugha, V.N., 2003. Nutrient changes during the fermentation of African oilbean (Pentaclethra macrophylla Benth) seeds. Pak. J. Nutr., 2: 320-323.
CrossRefDirect Link - Enujiugha, V.N., F.A. Thani, T.M. Sanni and R.D. Abigor, 2004. Lipase activity in dormant seeds of the African oilbean (Pentaclethra macrophylla Benth). Food Chem., 88: 405-410.
CrossRef - Enujiugha, V.N. and C.T. Akanbi, 2008. Quality evaluation of canned fermented oil bean seed slices during ambient storage. Afr. J. Food Sci., 2: 54-59.
Direct Link - Fogarty, W.M. and P.J. Griffin, 1973. Production and purification of metalloproteases of Bacillus polymyxa. Applied Microbiol., 26: 191-195.
Direct Link - Isu, N.R. and H.O. Njoku, 1998. Studies on the influence of temperature, relative humidity and microenvironment on the natural fermentation of African oil bean seeds to Ugba. Plant Foods Hum. Nutr., 52: 337-351.
CrossRef - Kabuo, N.O., J.O. Uzuegbu, C.N. Ubbaonu and E.U. Onyeka, 2007. Effect of chemical compounds produced by microorganisms on organoleptic properties of ugba (Pentaclethra macrophylla Benth) during fermentation. Proceedings of the Annual Conference of Nigerian Institute of Food Science and Technology, Oct. 22-25, Abuja, Nigeira, pp: 175-176.
- Kar, A. and A.D. Okechukwu, 1978. Chemical investigations of the edible seeds of Pentaclethra macrophylla Benth. Plant Foods Hum. Nutr., 28: 29-36.
CrossRef - Mba, A.U., M.C. Njike and V.A. Oyenuga, 1974. The proximate chemical compsotion of the amino content of some Nigerian oil seeds. J. Sci. Food Agric., 25: 1547-1553.
PubMedDirect Link - Mbadiwe, E.I., 1978. Nutritional evaluation of seeds of Pentaclethra macrophylla. Plant Foods Hum. Nutr., 28: 261-269.
CrossRef - Mbajunwa, O.K., J.O. Akingbala, K. Mulongoy and G. Oguntimein, 1998. Starter culture evaluation for the production of ugba from African oil bean seed (Pentaclethra macrophylla). J. Sci. Food Agric., 77: 127-132.
Direct Link - Mbata, T. and M.U. Orji, 2008. Process optimization in the production and preservation of Ugba, a Nigerian fermented food. Int. J. Microbiol., 4: 2-6.
Direct Link - Odoemelam, S.A., 2005. Proximate composition and selected physicochemical properties of the seeds of African oil bean (Pentaclethra marcrophylla). Pak. J. Nutr., 4: 382-383.
CrossRefDirect Link - Ogueke, C.C. and L.E. Aririatu, 2004. Microbial and organoleptic changes associated with ugba stored at ambient temperature. Niger. Food J., 22: 133-140.
Direct Link - Okafor, J.C., U.C. Okolonkwo and M.A.N. Ejiofor, 1991. Evaluation of oils from fermented and unfermented seeds of the African oil bean tree Pentaclethra macrophyla. Int. Tree Crops J., 7: 95-101.
Direct Link - Onwuliri, V.A., I. Attah and J.O. Nwankwo, 2004. Anti-nutritional factors, essential and non-essential fatty acids composition of Ugba (Pentaclethra macrophylla) seeds at different stages of processing and fermentation. J. Biological Sci., 4: 671-675.
CrossRefDirect Link - Ruiz-Teran, F. and J.D. Owens, 1999. Fate of oligosaccharides during production of soya bean tempe. J. Sci. Food Agric., 79: 249-252.
CrossRefDirect Link - Sokari, T.G. and C.K. Wachukwu, 1997. Simple rapid processing of African oil beans for ugba production. Int. J. Food Sci. Technol., 32: 77-79.
CrossRef - Njoku, H.O. and C.P. Okemadu, 1989. Biochemical changes during the natural fermentation of the African oil bean (Pentaclethra macrophylla) for the production of Ugba. J. Sci. Food Agric., 49: 457-465.
CrossRef