
D.M. Nejad
EM Lab Drug Applied Research Center and Development Complex, Tabriz University of Medical Sciences, Tabriz, Iran
J.S. Rad
Department of Histology, Tabriz University of Medical Sciences, Tabriz, Iran
L. Roshankar
Department of Histology, Tabriz University of Medical Sciences, Tabriz, Iran
M. Karimipor
Department of Histology, Tabriz University of Medical Sciences, Tabriz, Iran
A.A. Ghanbari
Department of Histology, Tabriz University of Medical Sciences, Tabriz, Iran
A. Aazami
EM Lab Drug Applied Research Center and Development Complex, Tabriz University of Medical Sciences, Tabriz, Iran
M.R. Valilou
Department of Veterinary Medicine, Islamic Azad University, Shabestar Branch, Shabestar, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 15 | Page No.: 1929-1934







ABSTRACT
The present study was designed to explore the spermatogenic effect of intraperitoneal administration of thiotepa in mice. It is known the chemotherapy could affect testicular function and result in infertility. There are few reports on the side effect of thiotepa on spermatogenesis when are used as anticancer drug. Therefore, in the present study the effect of thiotepa on spermatogenesis was evaluated by electron microscopy. A total of 20 balb/c mice were used in this study. They were divided into control and thiotepa treated groups. Thiotepa was administrated as 2.5 mg kg-1 for 5 days. The mice were sacrificed after 5 weeks and testicular specimens were removed, fixed in boueins fixative and 2.5% Glutaraldehide then prepared for light and electron microscopic study. Light microscopy showed that the thickness of germinal epithelium was decreased in thiotepa treated group and many seminiferous tubules have lost germinal cells. Electron microscopy revealed that in experimental group several intercellular spaces appeared between spermatogonial and sertoli cells. The basement membrane became irregular and intercellular vacuoles were present in sertoli and spermatogonial cells. Nuclear chromatin was condensed and there were several vacuolated mitochondria. It was indicated that thiotepa affected testicular germinal epithelium by both cytotoxic effect and induction of apoptosis.
PDF Abstract XML References Citation
How to cite this article
D.M. Nejad, J.S. Rad, L. Roshankar, M. Karimipor, A.A. Ghanbari, A. Aazami and M.R. Valilou, 2008. A Study on the Effect of Thiotepa on Mice SpermatogenesisUsing Light and Electronic Microscope. Pakistan Journal of Biological Sciences, 11: 1929-1934.
DOI: 10.3923/pjbs.2008.1929.1934
URL: https://scialert.net/abstract/?doi=pjbs.2008.1929.1934
DOI: 10.3923/pjbs.2008.1929.1934
URL: https://scialert.net/abstract/?doi=pjbs.2008.1929.1934
INTRODUCTION
Todays, some of the malignancies such as Testicular tumors, acute Leukemia Lymphomas can be cured by surgery, chemo or radiotherapy. For young male patients, chemotherapy regimens that are toxic to testicular function have made infertility an important problem. When the cancer is controlled, quality of life, which often includes the ability to have a normal child, becomes a major issue. Therefore, the long term toxicity following therapeutic intervention and resulting quality of life becomes more important issue for chemotherapy treatments. Testicular dysfunction is amongst the most common long-term side effects of chemotherapy in men. Germinal epithelial damage resulting in oligo or azoospermia has long been recognized as a consequence of treatment with chemotherapeutic agents.
Spermatogenesis is influenced negatively by many factors such as radiation therapy and cytotoxic drugs used for treatment of different cancers (Codrington et al., 2007; Howell and Shalet, 2005; Howell and Shalet, 2001; Liu et al., 2007; Meistrich et al., 2003). Recently, the thiotepa is used for treatment of various malignancies including breast, ovary, bladder cancers and Hodgkin and non-Hodgkin lymphoma (Bdul-Hai et al., 2007; Biron et al., 2008; Burgues et al., 2007; Tokuda et al., 2008). The previous studies on the side effect of chemotherapeutic drugs such as procarbazine, cyclophosphamide and chlorambucil showed long-term (up to years) azoospermia after treatment (Howell and Shalet, 2005; Sieniawski et al., 2008). There are a few reports on the side effect of thiotepa on the spermatogenesis when they are used as anticancer drugs, in the literature (Khil`kevich and Kurilo, 1992). Tumors are characterized by division, which is no longer controlled as it is in normal tissue. Cancer cells no longer have the normal checks and balances in place that control and limit cell division. The process of cell division, whether normal or cancerous cell, is through the cell cycle. The cell cycle goes from the resting phase, through active growing phases and then to mitosis (division). The ability of chemotherapy to kill cancer cells depends on its ability to halt cell division. So chemotherapy is most effective at killing cells that are rapidly dividing. Unfortunately, chemotherapeutic agents can not differentiate between the cancerous and the normal cells. It has been shown that one of the mechanisms in cell destruction following chemotherapy is apoptosis (Andriana et al., 2004; Bakalska et al., 2004; Habermehl et al., 2006; Hou et al., 2005; Sawhney et al., 2005). Some agents including genes factors, Testicular ischemia, heat stress, exposure to irradiation and toxic substances, could increase the rate of germ cell apoptosis in testis (Andriana et al., 2004; Bakalska et al., 2004; Habermehl et al., 2006; Hou et al., 2005; Liu et al., 2007; Sakallioglu et al., 2007; Sawhney et al., 2005). Thiotepa is a classical DNA directed alkylating agent currently used in high dose therapy approaches to the treatment of both hematopoietic and solid tumors (Bdul-Hai et al., 2007; Biron et al., 2008; Burgues et al., 2007; Tokuda et al., 2008). Thiotepa induces S-phase cell cycle arrest and cell death as a result of DNA cross-links or adduct S formation. Several studies have shown the harmful impact of thiotepa on the germinal epithelium of testis (Khil`kevich and Kurilo, 1992). These studies have not provided enough information about ultra-structural changes in germinal epithelium of testis and spermatogenesis. On other hand, the results of electron microscope in evaluating morphological pattern of nucleus, organelles, membrane and cell adhesion is more invaluable in comparison to light microscope. In the meantime, the knowledge about the ultra structural changes of germinal stem cells and spermatogenic cells following the administration of thiotepa can help to understand the mechanisms of action of thiotepa.
In the current study, the morphological pattern of nucleus, organelles, membrane and cell junction of spermatogenic and somatic cells of mice testis after treatment with thiotepa in evaluated.
MATERIALS AND METHODS
Animals: Twenty balb/c mice aging 6-8 weeks was used in this study. The mice were divided into two groups, control and thiotepa treated. Thioteapa was injected intraperitoneally for five days as 2.5 mg kg-1. The dosage was selected as reported by previous studies. After 5 weeks from beginning of treatment, all animals were sacrificed in both groups. The testes were removed from the abdominal cavity and separated from the epididymis with care by using a surgical blade and then testicular specimen from right testis were fixed in boueins fixative for 48 h and then processed for light microscopy. The paraffin sections stained with H and E and studied with light microscope.
The specimens from the left testis were fixed in 2.5% Glutaraldehyde (Pro. Sci. Tech. Au) for 12 h and washed with 0.2 M phosphate buffer and post fixed with 2% osmium tetroxide (Taab, UK) for 2 h. After Dehydration, Clearing and Infiltration they embedded in resin (Pro. Sci. Tech. Au) and sectioned using ultramicrotome (Richert-Jung, Au). Semithin stained with toluidine blue and studied with light microscope. The thin sections (80 nm thickness) were stained with uranyl acetate and lead citrate and were studied with Leo 906 transmission electron microscope (Leo, Germany).
RESULTS
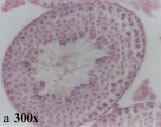
Light microscopy: Light microscopy revealed that seminiferous tubules have a thick germinal epithelium and their lumina contained numerous mature sperms. Different stages of spermatogenesis (12 stages) could be recognized based on morphological criteria. Interstitial spaces contained connective tissue and lydig cells (Fig. 1a).
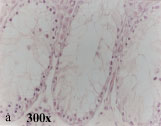
In the treated group, as compared to the control group, the spermatogenic epitheliums were reduced in thickness and in some cases the tubules had only sertoli cells and there were no mature sperms in any tubule. Therefore recognition of different stages of spermatogenesis was impossible. There were also several vacuoles between germinal cells in this group (Fig. 2a). Spermatogenic Index (SI) in control group was 3.01 ± 0.1 and in treated group it reduced to 0.32 ± 0.01 which is significant (p = 0.001).
Electron microscopy: Electron microscopy revealed that in control group, spermatogonia was rested on the basement membrane and had round or oval nuclei and small mitochondria and less developed endoplasmic reticulum (Fig. 1b). The sertoli cells could easily be distinguished regarding their large nuclei with one or two deep indentation and large nucleolus. Sertoli cells were rested on the basement membrane and the junction between two adjacent sertoli cells was obvious.
This structure has important role in normal spermatogenesis by forming Blood Testes Barrier (BTB) and is composed of three elements (Fig. 1b, c).
| • | Merged plasmalemma of adjacent sertoli cells in the center. |
| • | Dilated endoplasmic reticulum in both sides. |
| • | Bundles of filaments that are sandwiched between plasmalemma and endoplasmic reticulum. |
 |  |
 |  |
 |  |
| Fig. 1: | Light and electron micrographs from mouse testes in control group |
| a: | A cross section of seminiferous tubule showing germinal epithelium with different cell type, H and E staining Spermatogonium and sertoli cell on basement membrane. Junction between two sertoli cells (blood testes barrier) (arrow head) Blood testes barrier with higher magnification. Mp: merged plasma membrane, Ac: actin filaments, Er: endoplasmic reticulum Spermatids at different stages of spermatogenesis Sperm head. Ectoplasmic specialization (arrow head) Ectoplasmic specialization with higher magnification. Er: endoplasmic reticulum, Ac: actin filaments, Mp: merged plasmalemma |
The spermatogenic cells, including round spermatids, could be seen at various stages of spermatogenesis such as Golgi and cap stages (Fig. 1d).
In control group, spermatids were embedded in the sertoli cells and formed Ectoplasmic Specialization (ES) which is an indication of final stages of spermatogenesis. ES composed of three segments.
| • | Merged membrane of spermatid and sertoli cells. |
| • | Dilated endoplasmic reticulum in the sertoli cell. |
 |  |
 |  |
 |  |
| Fig. 2: | Light and electron micrographs from mouse testes in thiotepa treated groups |
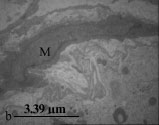
| a: b: c: d: e: f: | A cross section of seminiferous tubule. Note germinal cell depletion Spermatogonia is separated from basement membrane. The basement membrane is wavey. A contracted myoid cell is shown in the figure (M) Spermatogonial cells are separated from sertoli cell and basement membrane by large vacuoles Spermatogonium contain large intracytoplasmic vacuoles (V) Blood-Testes-Barrier is thinner and irregular in comparison to control group An apoptotic cell with crescent like chromatin condensation (arrow head) Vacuolated mitochondria and a deformed cilium (lower right corner) in a spermatogonic cell and sperm |
| • | Bundles of actin filaments that are sandwiched between sertoli plasmalemma and endoplasmic reticulum of sertoli cell (Fig. 1e, f). ES has important role in spermatogenesis especially in converting of round spermatid to mature spermatozoa. |
Study of testis in the treated group showed that spermatogonia and sertoli cells were separated from basal lamina and neighboring cells by large spaces. There were also large vacuoles in the cytoplasm of sertoli and spermatogonia cells. Basement membrane was morphologically irregular and wavey. Myoid cells in the boundary tissue were thickened and condensed (Fig. 2b, c).
BTB in this group showed a dramatic irregularity and appeared thinner than in the control group (Fig. 2d).
In this group, a lot of apoptotic cells were observed, they were recognized based on their crescent like chromatin condensation (Fig. 2e). Spermatogenic cells had also several damaged organelles such as vacuolated mitochondria and abnormal axonem (Fig. 2f). Due to destructive effect of thiotepa different spermatogenic cells and stage of spermatogenesis was rarely observed.
DISCUSSION
In the present study, the effect of thiotepa, as an anticancer drug, on germinal epithelium has been investigated. Present results showed that the thickness of germinal epithelium decreased in the thiotepa treated group. This is in agreement with the findings of other researchers that chemotherapy could affect spermatogenesis (Codrington et al., 2007; Howell and Shalet, 2005; Howell and Shalet, 2001; Khil`kevich and Kurilo, 1992; Meistrich et al., 2003; Sieniawski et al., 2008).
We also showed the presence of several spaces between the spermatogonial and sertoli cells and between them and basal lamina. In this regard, it has been shown that sertoli-germ cell adhesion enhances the survival of germ cells and cadherins mediated adhesion between sertoli and germ cells in vitro (Newton et al., 1993). Cadherins as an important molecular system controls interaction between sertoli and germ cells and promotes the survival of germ cells. It has thought that cadherin based adhesion generates intracellular signaling cascades that control cell survival, migration and maturation (Honecker et al., 2004). In our TEM study both BM and myoid cells were altered structurally in experimental group. BM in the treated group was irregular and had some swirls. Similar findings were observed in irradiated rats (Sawada and Esaki, 2003) and after efferent ligation (Richardson et al., 1998). The swirls may be due to contraction of myoid cells or reduction of tubular diameter. In this study the BM was continuous even in the absence of germ cells. It means that germ cells are probably not involved in BM production. It has been shown that during tubular injury, for survival and regeneration of stem cells, the BM is produced (Sawada and Esaki, 2003). Testicular BM is synthesized by both sertoli and myoid cells (Skinner et al., 1985). In our TEM study it appeared that the thickness of BM is increased in thiotepa treated group. Increasing of BM thickness similarly occurs in: aging (Siu and Cheng, 2004), efferent ligation (Richardson et al., 1998) and after irradiation (Sawada and Esaki, 2003). This alteration either is due to increasing of production by sertoli or myoid cells or reduction of proteolysis rate in ECM. In efferent ligation the gene expression of laminin is changed in sertoli cells and their protein synthesis is enhanced. The nucleolus of myoid cell in thiotepa treated groups were appeared shorter and had a contractile form in comparison with control group. Myoid cells contain abundant actin filaments which are distributed in the cells in a species specific manner. The arrangement of actin filaments in the cells changes during postnatal development and the distribution of spermatogenesis such as cryptorchidism. Myoid cells also have myosin, desmin and actinin (Maekawa et al., 1996). In other words they are kind of smooth muscle cells (Virtanen et al., 1986). Several substances such as prostoglandins, oxytocin, TGF-β have been suggested to affect the contraction of these cells (Maekawa et al., 1996). These finding suggest that probably after treatment with thiotepa, damaging of epithelium stimulates secretion of some factors such as oxytocin or prostaglandins and cause myoid cells contraction. Alteration in BTB could affect normal spermatogenesis and may result in transient or long-term spermatogenesis defect. Microtubular arrangement in the sperm flagella (axonem) is important for energy production and sperm motility. Any changes, in the arrangement of flagellar microtubules may affect motility and result in subfertility. The changes, such as separation of spermatogonial cells from sertoli cells and basal lamina could also be considered as preapoptotic sign. Other finding such as nuclear condensation, chromatin margination and mitochondrial vacuolization are additional evidence for appearance of apoptotic cells. The cells with apoptotic feature were numerous in experimental group than the control group. In support of this finding there are some reports showing that anticancer drugs could induce apoptosis in germinal epithelium (Andriana et al., 2004; Bakalska et al., 2004; Habermehl et al., 2006; Hou et al., 2005; Sawhney et al., 2005). It is also known that chemotherapy agent, especially alkylating agents, tend to act on cells that have a high mitotic activity such as spermatogonial cells (Codrington et al., 2007; Howell and Shalet, 2001, 2005). On the other hand, apoptosis has a critical role on the removal of damaged spermatogonial cells to prevent the formation of abnormal sperms (Cai et al., 1997). It is also shown that spermatocytes that fail to complete their mitotic division are removed by apoptosis (Print and Loveland, 2000). It appears that thiotepa as an alkylating agent induces apoptosis of spermatogenic cells.
CONCLUSION
Present results indicate that administration of thiotepa, as an anticancer drug in mice, destroys testicular germinal epithelium by both cytotoxic effect and induction of apoptosis.
ACKNOWLEDGMENT
The authors are thankful to Drug Applied Research Center, Tabriz University of Medical Sciences, for their financial support of the project.
REFERENCES
- Andriana, B.B., T.W. Tay, I.M.A. Awal, Y. Kanai, M. Kurohmaru and Y. Hayashi, 2004. An ultrastructural study on cytotoxic effects of mono (2-ethylhexyl) phthalate (MEHP) on testes in Shiba goat in vitro. J. Vet. Sci., 5: 235-240.
PubMed - Bakalska, M., N. Atanassova, Y. Koeva, B. Nikolov and M. Davidoff, 2004. Induction of male germ cell apoptosis by testosterone withdrawal after ethane dimethanesulfonate treatment in adult rats. Endocr. Regul., 38: 103-110.
PubMed - Bdul-Hai, A., L. Weiss, D. Ergas, I.B. Resnick, S. Slavin and M.Y. Shapira, 2007. The effect of high-dose thiotepa, alone or in combination with other chemotherapeutic agents, on a murine B-cell leukemia model simulating autologous stem cell transplantation. Bone Marrow Transplant., 40: 891-896.
PubMed - Biron, P., M. Durand, H. Roche, T. Delozier and C. Battista et al., 2008. Pegase 03: A prospective randomized phase III trial of FEC with or without high-dose thiotepa, cyclophosphamide and autologous stem cell transplantation in first-line treatment of metastatic breast cancer. Bone Marrow Transplant., 41: 555-562.
PubMed - Burgues, J.P., L. Gomez, J.L. Pontones, C.D. Vera, J.F. Jimenez-Cruz and M. Ozonas, 2007. A chemosensitivity test for superficial bladder cancer based on three-dimensional culture of tumour spheroids. Eur. Urol., 51: 962-969.
PubMed - Cai, L., B.F. Hales and B. Robaire, 1997. Induction of apoptosis in the germ cells of adult male rats after exposure to cyclophosphamide. Biol. Reprod., 56: 1490-1497.
CrossRefPubMedDirect Link - Codrington, A.M., B.F. Hales and B. Robaire, 2007. Exposure of male rats to cyclophosphamide alters the chromatin structure and basic proteome in spermatozoa. Hum. Reprod., 22: 1431-1442.
CrossRefPubMedDirect Link - Habermehl, D., B. Kammerer, R. Handrick, T. Eldh and C. Gruber et al., 2006. Proapoptotic activity of Ukrain is based on Chelidonium majus L. alkaloids and mediated via a mitochondrial death pathway. BMC Cancer, Vol. 6.
CrossRefDirect Link - Honecker, F., A.M. Kersemaekers, M. Molier, P.C. Van Weeren and H. Stoop et al., 2004. Involvement of E-cadherin and beta-catenin in germ cell tumours and in normal male fetal germ cell development. J. Pathol., 204: 167-174.
PubMed - Hou, M., D. Chrysis, M. Nurmio, M. Parvinen, S. Eksborg, O. Soder and K. Jahnukainen, 2005. Doxorubicin induces apoptosis in germ line stem cells in the immature rat testis and amifostine cannot protect against this cytotoxicity. Cancer Res., 65: 9999-10005.
CrossRefPubMedDirect Link - Howell, S.J. and S.M. Shalet, 2005. Spermatogenesis after cancer treatment: damage and recovery. J. Natl. Cancer Inst. Monogr., 34: 12-17.
CrossRefPubMedDirect Link - Howell, S.J. and S.M. Shalet, 2001. Testicular function following chemotherapy. Hum. Reprod. Update, 7: 363-369.
PubMed - Khil'kevich, L.V. and L.F. Kurilo, 1992. The gametotoxic effect of antenatal exposure to thiotepa in mice. Ontogenez, 23: 401-406.
PubMed - Liu, G., P. Gong, L.R. Bernstein, Y. Bi, S. Gong and L. Cai, 2007. Apoptotic cell death induced by low-dose radiation in male germ cells: Hormesis and adaptation. Crit. Rev. Toxicol., 37: 587-605.
CrossRefPubMedDirect Link - Maekawa, M., K. Kamimura and T. Nagano, 1996. Peritubular myoid cells in the testis: Their structure and function. Arch. Histol. Cytol., 59: 1-13.
PubMed - Meistrich, M.L., G. Wilson, K.L. Porter, I. Huhtaniemi, G. Shetty and G.A. Shuttlesworth, 2003. Restoration of spermatogenesis in dibromochloropropane (DBCP)-treated rats by hormone suppression. Toxicol. Sci., 76: 418-426.
PubMed - Newton, S.C., O.W. Blaschuk and C.F. Millette, 1993. N-cadherin mediates sertoli cell-spermatogenic cell adhesion. Dev. Dyn., 197: 1-13.
CrossRefPubMedDirect Link - Print, C.G. and K.L. Loveland, 2000. Germ cell suicide: New insights into apoptosis during spermatogenesis. Bioessays, 22: 423-430.
PubMed - Richardson, L.L., H.K. Kleinman and M. Dym, 1998. Altered basement membrane synthesis in the testis after tissue injury. J. Androl., 19: 145-155.
PubMed - Sakallioglu, A.E., B.H. Ozdemir, O. Basaran, A. Nacar, D. Surena and M.A. Haberal, 2007. Ultrastructural study of severe testicular damage following acute scrotal thermal injury. Burns, 33: 328-333.
PubMed - Sawada, H. and M. Esaki, 2003. Electron microscopic observation of 137Cs-irradiated rat testis: production of basal laminae for germ cells, despite their absence. J. Elect. Microsc. (Tokyo), 52: 391-397.
PubMed - Sawhney, P., C.J. Giammona, M.L. Meistrich and J.H. Richburg, 2005. Cisplatin-induced long-term failure of spermatogenesis in adult C57/Bl/6J mice. J. Androl., 26: 136-145.
PubMedDirect Link - Sieniawski, M., T. Reineke, L. Nogova, A. Josting, B. Pfistner, V. Diehl and A. Engert, 2008. Fertility in male patients with advanced Hodgkin lymphoma treated with BEACOPP: A report of the German Hodgkin Study Group (GHSG). Blood, 111: 71-76.
PubMed - Siu, M.K. and C.Y. Cheng, 2004. Extracellular matrix: Recent advances on its role in junction dynamics in the seminiferous epithelium during spermatogenesis. Biol. Reprod., 71: 375-391.
PubMed - Skinner, M.K., P.S. Tung and I.B. Fritz, 1985. Cooperativity between Sertoli cells and testicular peritubular cells in the production and deposition of extracellular matrix components. J. Cell. Biol., 100: 1941-1947.
PubMed - Tokuda, Y., T. Tajima, M. Narabayashi, K. Takeyama and T. Watanabe et al., 2008. Phase III study to evaluate the use of high-dose chemotherapy as consolidation of treatment for high-risk postoperative breast cancer: Japan Clinical Oncology Group study, JCOG 9208. Cancer Sci., 99: 145-151.
PubMed - Virtanen, I., M. Kallajoki, O. Narvanen, J. Paranko, L.E. Thornell, M. Miettinen and V.P. Lehto, 1986. Peritubular myoid cells of human and rat testis are smooth muscle cells that contain desmin-type intermediate filaments. Anat Rec., 215: 10-20.
PubMed