
N. Amirmozafari
Department of Microbiology, School of Medicine, Iran University of Medical Sciences, Tehran, Iran
F. Ghazi
Iran University of Medical Sciences, Tehran, Iran
A. Mostafazadeh
Department of Microbiology and Immunology, Cellular and Molecular Research Center, Babol University of Medical Sciences, Babol, Iran
A. Mostafaie
Department of Protein Biochemistry, Medical Biology Research Center, Kermanshah University of Medical Sciences, Kermanshah, Iran
R. Rajabnia
Department of Microbiology, Iran University of Medical Sciences, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 2 | Page No.: 188-194







ABSTRACT
Heat shock protein (hsp) is highly conserved, that serves a wide range of function in protein folding and transport. It protect from various type of stress including heat shocks. However, it is well known that the virulence of B. melitensis is more than B. abortus, but there is not any strong evidence to verify it. For this purpose, in refer to potent antigenicity of hsps in various infectious as well as some hsp molecules act as potent activator of macrophage (danger signal), we hypothesized that difference in virulence between B. abortus and B. melitensis may be originated from difference in pattern of response to heat shock induced by high degree of fever that usually present in brucellosis. To this end, five B. abortus and five B. melitensis strains isolated from cows and human, were subjected to 39, 40 and 42 ° C heat shocks. The bacterial whole cell proteins were extracted and resolved by SDS-PAGE. Western blotting was used to detect antibody production against the extracted bacterial proteins especially hsp60 in both control and patient sera. SDS-PAGE gels revealed protein bands mainly in the range of 10-100 kDa. The amounts of a 60 kDa protein band (hsp60) was significantly enhanced following heat shock at 42 ° C in relation to the unheated cells in both bacterial species. The heat shock responses in B. abortus and B. melitensis point to the higher production of a 60 kDa protein (hsp60) in both bacterial species, especially in B. abortus. It seems that, lower hsp60 production by B. melitensis would induce a relatively much lower immune response against the bacterium leading to its greater virulence potentials; the sera from Brucellosis patients reacted with several of these cell derived protein bands in western blots, none of which were reactive with sera from healthy individuals. The western blot protein bands showed striking differences. This observation points to the immunogenic properties of hsps, specially the overwhelming response to hsp-60. Therefore, hsp-60 can be a good antigenic candidate for engineering subunit vaccine against Brucella, as well as for ELISA test development.
PDF Abstract XML References Citation
How to cite this article
N. Amirmozafari, F. Ghazi, A. Mostafazadeh, A. Mostafaie and R. Rajabnia, 2008. Comparison of Heat Shock Response in Brucella abortus and Brucella melitensis. Pakistan Journal of Biological Sciences, 11: 188-194.
DOI: 10.3923/pjbs.2008.188.194
URL: https://scialert.net/abstract/?doi=pjbs.2008.188.194
DOI: 10.3923/pjbs.2008.188.194
URL: https://scialert.net/abstract/?doi=pjbs.2008.188.194
INTRODUCTION
Brucellosis is an important and widespread zoonotic disease which is caused by the genus of Brucella. Brucella melitensis and Brucella abortus are the two most common causative agents of Brucellosis in both human and cattle (Yang, 1995; Corbel, 1997). The pathogenicity of B. melitensis is generally higher than B. abortus for human. Brucellae are facultative intracellular and gram negative coccobacilli which usually survive within phagocytic cells such as macrophages (Lin and Ficht, 1995; Baloglu et al., 2000). They do not possess spores or elaborate any known exotoxins (Jawets et al., 2003). Their pathogenicity is the result of a multi-factorial phenomenon. Environmental growth and intracellular conditions of the bacteria can significantly influence cellular proteins level expression. Host-parasite interactions during natural infections will expose the bacteria to many physical and biological stresses (Lin et al., 1992). Phagocytosed and intracellular bacteria survive by adopting themselves to the extreme conditions of lysosome and intracellular environmental changes. This adaptation will prevent bacterial lysis by the intracellular defense mechanisms of the phagocytic cells. When Brucella sp. are exposed to elevated temperatures or heat shocks, they began to synthesize heat shock proteins that not only can help the bacteria to survive high temperatures; but also, can play a potentiating role in the pathogenesis of the bacteria. Heat shock proteins (hsps) produced by different bacteria are highly conserved and probably allow adaptation of the producing bacteria to many stressful conditions. Some of these hsps, members of Gro EL family, are readily recognized by the host immune system during infections and the ensuing immune response can have a protective role in the host. The most important member of the Gro EL hsp family is a 62 kDa protein (hsp 62) (Yilan et al., 1998).
In the present study, B. melitensis and B. abortus strains isolated from human and cows were exposed to different heat shocks. Total bacterial protein profiles of the shocked and the control B. melitensis and B. abortus isolates were analyzed by sodium dodecyle sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Hsp60 of shocked and un-shocked bacteria were identified by western blotting using anti-hsp60 specific antibodies. The acute phase sera of human patients, who were naturally infected with Brucella sp., were isolated for western blotting and serological tests such as wright, combs wright, 2ME, Brucella IgG and Brucella IgM were performed for each sera sample. The presence of anti-hsp60 antibodies and other anti-Brucella protein antibodies in the acute phase sera of human patients and control sera were analysed by western blots.
MATERIALS AND METHODS
Isolation and culture of bacterial strains: This study were performed in Microbiology Department, School of Medicine, Iran University of Medical Sciences and Health Services, Tehran, Iran in 2006. Five strains of B. abortus and five strains of B. melitensis were isolated from cows and human, respectively and were cultured on Brucella agar plates. Five milliliters blood samples were taken from each individual and promptly cultured in Brain Heart Infusion (BHI) broth at 37°C under 5% CO2-air. The media were incubated for 4 week, during which 5 sub-cultures (days 3, 7, 14, 21 and 28) were made onto solid medium Brucella agar containing 5% sheep blood. The Brucella agar plates were incubated at 37°C under 5% CO2-air for 48-72 h. Any colony seen was subjected to a gallery of biochemical tests such as Gram stain, Methyl red, Vogous Proskauer, TSI, urease, oxidase, catalase tests and growth on media containing different concentration of basic fuschin and thionine.
Serum samples: Two milliliters of serum samples were taken from brucellosis patient and control group and stored at -20°C. The serological tests were performed for all the sera.
Heat shocks: The bacterial colonies were transferred into 4 different tubes containing Brucella broth and incubated with shaking at 37°C until the optical density (at 600 nm) reached to 1.0 (Gomes et al., 2000). After incubation, the tubes were placed at 37, 39, 40 and 42°C for 3 h. The bacterial suspensions were centrifuged at 5000 x g for 30 min. The pellets were washed twice with normal saline and once with 10 mM Tris-base buffer (pH 7.5). The washed cells were resuspended in 2 mL Tris-base buffer and subsequently, 10 mL of cold acetone was added to each tube on ice. The tubes were stored at -20°C for 5-7 days, after which they were centrifuged at 5000 x g for 30 min. The supernatants were decanted and the killed bacteria were allowed to dry and then stored at -20°C.
Protein extraction: The bacterial proteins were extracted according to the method of Rosenbakh and his Colleques (Abdolalizadeh et al., 2002) with some modifications. The bacterial cells were washed with Tris-base-saline buffer (20 mM Tris-base, 100 mM NaCl; pH 7.5). The wet bacterial pellets (0.16 g) were resuspended in 640 μL of TSB (Tris buffered saline). PMSF (Phenyl Methane Sulfonyl Fluoride) and EDTA (Ethylene Diamine Tetra Acetic acid) were added each at final concentration of 1 mM. Subsequently, 4.8 mL of lysis buffer (Tris-base buffer containing 2% SDS, 10% glycerol and 0.7 M 2ME at pH 7.2) was added to each tube and incubated at 60°C for 4 h. After cooling the tubes to room temperature, lysosyme (1 mg for each 100 mg of dry weight) was added and incubated at 37°C overnight. The cell suspensions were plated in boiling water for 10 min and then centrifuged at 30000 x g for 30 min. The supernatant was divided into aliquots and kept at -20°C.
Electrophoresis: SDS-PAGE was performed in 13.5% resolving and 5% stacking polyacrylamide slab gels (Abdolalizadeh et al., 2002). The bacterial protein extracts were mixed with equal volume of SDS-PAGE sample buffer and incubated for 10 min at 100°C. Fifteen microliter of each sample was loaded into each well. Electrophoresis was performed at constant voltage (150 volts) and the gels were stained with Coomassie brilliant blue. The stained gels were stored in 10% acetic acid solution.
Immunoblotting: Western blotting was preformed according to the method of Towbin (Mostafaie, 1999) using transfer buffer (Tris-base 25 mM, glycin 192 mM and methanol 15% with pH 8.3) and PVDF membrane. A total of 10 sera collected from the patients with brucellosis who had wright titers of ≥1:320 and 2ME titers of ≥1:160 were used. PVDF membranes were rinsed with methanol for 5 min and then used for transfer. Transfer of protein bands from SDS-PAGE gel into PVDF membrane was carried out at 75 mA for 30 min, 100 mA for 30 min, 200 mA for 90 min and finally 300 mA for 90 min. Following transfer, the membranes were blocked in PBS containing 0.5% tween20, then washed three times PBS containing tween20 0.05% (PBS-T). The membranes were incubated with anti-hsp60 antibody (1/4000) or human sera (1/50) for 90 min at room temperature. After washing of the membranes four times with PBS-T, the secondary antibody conjugated with HRP (1/4000 for anti-human Igs and 1/4000 For anti-rabbit IgG) was added for 90 min. After washing four times with PBS-T, the membranes were incubated in enzyme substrate solution (3, 3-Diaminobenzidine and H2O2) until reactive bands developed. The membranes were finally washed with distilled water, dried and kept in dark.
RESULTS
In the present study, five strains of B. abortus and five strains of B. melitensis were isolated from cows and human suffering from brucellosis, respectively. In addition, ten sera with Wright titers of = 1:320 and 2-ME titers of = 1:160 collected from patients with brucellosis. All sera were positive for Brucella IgG and Brucella IgM in ELISA tests.
The isolated and mass cultured bacteria were placed under heat shocks at 39, 40 and 42°C. Thereafter the extracted proteins from the heat-shocked as well as the non shocked controls were resolved by SDS-PAGE (Fig. 1).
SDS-PAGE pattern of B. abortus (lane a and b) was composed of protein bands mainly with molecular mass in the range of 10-100 kDa. The major protein groups were in the range of 30-75 and 14 -20 kDa. The amounts of a 60 kDa protein band (hsp60) was significantly enhanced following heat shocks compared to controls.
SDS-PAGE pattern of B. melitensis (lane c and d) also indicated protein bands in the range of 10-100 kDa. The most significant protein groups of these isolates were in the range of 45-75 and 14-30 kDa. In the pattern of heat shocked bacteria in 42°C, not only a few new proteins bands appeared, but also there were a generalized increase in the level of most protein band. A 78 kDa and another protein band in the range of 40-45 kDa were present only in the control non-shocked cells and not in any of the heat-shocked bacteria. The amounts of a 60 kDa protein band (hsp60) was significantly enhanced following heat shocks in relation to the unheated cells. The results also indicated that heat shock responses induce more expression of a 60 kDa protein (hsp60) in B. abortus than in B. melitensis.
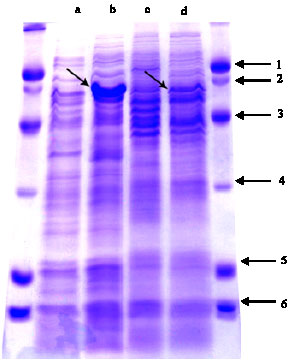 | |
Fig. 1: | SDS-PAGE pattern of extracted proteins from the heat-shocked (42°C) and non shocked control B. abortus (a and b, respectively) and B. melitensis (c and d, respectively). The right and left sides is protein size marker on the basis of kDa |
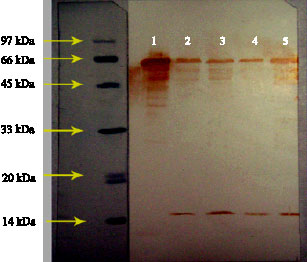 | |
Fig. 2: | Reaction of pure Escherichia coli hsp60 (1), shocked and un-shocked B. melitensis (2 and 3, respectively), un-shocked and shocked B. abortus (4 and 5, respectively) with anti-hsp60 specific antibody |
Immunoblotting of Escherichia coli hsp60 and the extracted proteins from heat shocked and control B. abortus and B. melitensis with polyclonal anti-hsp60 antibody showed that this antibody detect homologous proteins with similar molecular mass (Fig. 2). Furthermore
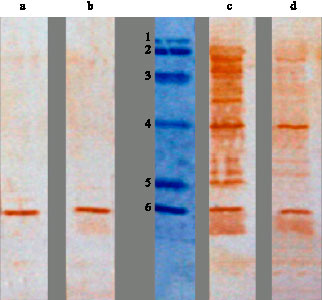 | |
Fig. 3: | Reactions of shocked B. abortus protein extracts with sera from healthy subjects (a, b) and brucellosis patients (c, d). The middle lane is protein size marker (78, 66, 45, 29, 18.5, 14.5 kDa, respectively) |
these results indicated that heat treatment induce hsp60 in the two bacterial species (lane 2 and 5), although response of B. abortus is more intense than B. melitensis. These results correlate considerably with the results of SDS-PAGE and verify the effect of heat on induction of hsp60. As immunoblotes of B. abortus and B. melitensis showed, anti-hsp60 antibody react with an additional protein band with molecular mass of 15-16 kDa with unknown specificity.
The shocked and un-shocked bacterial protein extracts that resolved by SDS-PAGE and transferred to PVDF membranes, were incubated with sera from brucellosis patients as well as healthy subjects. The sera from brucellosis patients reacted with several protein bands of B. abortus with molecular mass 14, 29 and 60 kDa (Fig. 3, 4). The 60 kDa band was the most significant and showed strong reactions with all patients sera. The mentioned protein bands showed week or no reactivity against sera from healthy individuals.
Furthermore, reactions of B. melitensis proteins with patients and control sera indicated that sera from patients but not from the healthy individuals strongly react with proteins extracted from heat-shocked bacteria in the positions 10, 60 and 100 kDa (Fig. 5, 6). The result indicates that none of these bands showed up in the reactions between control sera with protein extracted from un-shocked B. melitensis. The reactions between patients and controls sera with un-shocked B. melitensis extracted proteins are shown in Fig. 6. Many new protein bands
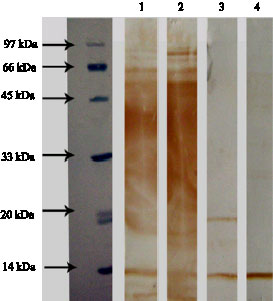 | |
Fig. 4: | Reactions of un-shocked B. abortus proteins with sera of brucellosis patients (1, 2) and healthy subjects (3, 4) |
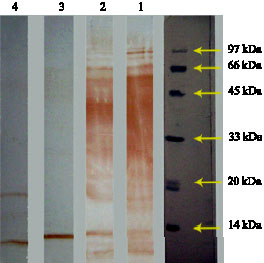 | |
Fig. 5: | Reactions of shocked B. melitensis protein extracts with sera of brucellosis patients (1, 2) and healthy people (3, 4) |
show up in the interaction of un-shocked bacterial proteins with patients sera that are absent in reaction with control sera. A common 60 kDa band is seen in reactions between shocked B. melitensis proteins with all patients` sera. This band is less frequently detected in the reaction between un-shocked bacterial proteins with the patient`s sera.
The heavily stained smear seen in the upper and middle sections of the blots correlates to the presence of high levels of anti Brucella LPS antibodies in patient`s sera. A 14 kDa band is also detected in the immunoblots of all samples, both patients and healthy control.
 | |
Fig. 6: | Reactions of un-shocked B. melitensis protein extracts with sera of brucellosis patients (1, 2) and healthy people (3, 4). |
Significant differences in protein bands were not detected following electrophoresis of 39 and 40 heat shocked cells in relation to the unheated bacteria.
DISCUSSION
In order to find the effect of heat stress on expression of hsp60, five strains of B. abortus and five strains of B. melitensis were isolated from cows and human, respectively and shocked at 39, 40 and 42°C. Total bacterial protein profiles of shocked and un-shocked bacteria were resolved by SDS-PAGE and analyzed against anti-hsp60 specific antibody and the patient`s sera by immunobloting.
Comparing the protein profiles of the heat-shocked versus un-shocked bacteria showed a general increase in protein expression levels in the heat-shocked samples; especially in a 60 KDa protein band (hsp60) in 42°C shocked bacteria. This is in agreement with similar study preformed by Lin et al. (1992). In this study, they observed a significant increase in the level of the 60 kDa protein following heat-shocking of B. abortus. The present study which was conducted on clinical isolates of B. melitensis and B. abortus, did not show any significant protein level expression differences in bacteria shocked at 39 and 40°C in comparison with the un-shocked cells. In the Lin et al. (1992) study, maximum hsp proteins levels were produced in B. abortus cells shocked at 42-46°C. The duration of bacterial heat shock used in this study was 3 h as the Lin investigation found the maximum heat shock response occurring after 2-3 h. Besides cell wall antigen such as the outer membrane antigen, there are some other cellular proteins that can play important immunologic roles during disease process (Lindler et al., 1996). These antigens can act as important candidate for serological diagnosis of brucellosis and/or subunit vaccine development (Angel et al., 2003; Juliana et al., 2007). The levels of hsp proteins are significantly increased under heat shock (Kaufmann, 1990). Some of these hsps are readily recognized by the host immune system during natural infections. The immunogenicity of the shocked and un-shocked bacterial proteins against the acute phase sera of human patients, who were naturally infected with Brucella, were analyzed by western blots.
This study aims to elucidate the immunogenicity of the heat-shocked and un-shocked bacterial proteins, especially hsp60, against patients and control sera. The study`s long term objective is to find a suitable antigenic candidate for specific serological tests as well as for subunit vaccine studies.
Highly pure E. coli hsp60 and polyclonal anti-hsp60 antibody were used to specifically detect hsp60 protein expression levels in the B. melitensis and B. abortus. Pure hsp60 protein and also shocked and un-shocked extracted B. melitensis and B. abortus protein samples which resolved and blotted, were incubated with polyclonal anti-hsp60 antibody. As the results showed (Fig. 2), the antibody reacted much more strong with the 60 kDa protein extracted from the heat shocked B. melitensis and B. abortus isolates in relation with those recovered from the un-shocked bacteria. This indicates an increase level of hsp60 production under heat shock conditions. The reaction of the anti-hsp60 antibodies with a 14 kDa protein band is probably due to the general cross-reaction of the polyclonal antibodies.
The heat-shocked bacterial protein reactions with patients sera showed striking differences with those of control sera reaction. This result points to the immunogenic nature of hsp60 and the relatively strong immune response induced by the human immune system during natural infections.
As it is concluded from SDS-PAGE and Immunoblotting results, hsp60 production in B. abortus following heat shocks was significantly greater than B. melitensis. We initially thought that the greater virulence of B. melitensis relative to B. abortus, was probably due to the higher in vivo induction of hsp60 following stress shocks. But paradoxically, we observed higher hsp60 production by B. abortus relative to B. melitensis. It thus can be postulated that the higher levels of hsp60 production by B. abortus and the enhanced antigenic properties of hsp60, which leads to higher anti-hsp60 antibodies can in effect counteract the higher hsp60 production and subsequently the pathogenicity of B. abortus. In other words, the increased hsp production by B. abortus can induce a potent immune response which will in effect help to restrict bacterial virulence. On the other hands, lower hsp60 production by B. melitensis would induce a relatively much lower immune response against the bacterium leading to its greater virulence potentials.
As Wei et al. (1999) has indicated, the mammalian hsp60 acts as a danger signal for innate immune system such as macrophages; therefore, another reason for lower pathogenicity of B. abortus compared to B. melitensis may be the role of Brucella hsp60 as a danger signal. hsp60 Receptors on the surface of macrophages has been designated as toll-like receptor-4. After binding of hsp60 to these receptors, cytokines such as TNF-α, IL-1 and IL-6 are released. T-helper-2 (Th2) cell is shifted to Th1 by TNF-α. Th1 cells play a major role in Cell Mediated Immunity (CMI) which is very effective against intracellular bacteria. Therefore, low production of hsp60 produces less TNF-α leading to more pathogenicity. On the other hands, high production of hsp60 produces higher TNF-α leading to activation of CMI and therefore, lower pathogenicity. At the result, the pathogenicity of B. melitensis by producing lower levels of hsp60 is higher than B. abortus.
The routine serological tests commonly used for sero-diagnosis of animal and human brucellosis, is based on determination of anti-LPS antibody levels. These antibodies persist at relatively high titers for long period of times, even after full patients recovery (Abdolalizadeh et al., 2002; Mostafaie et al., 2005). Unfortunately, these serological tests have many cross-reactions with other Gram negative bacteria (Abdolalizadeh et al., 2002; Mostafaie et al., 2005). Because of these problems, researchers have long sought to develop more specific non-LPS based serological tests directed against proteinatious component of the bacteria. Therefore, hsp60 can potentially be used as a suitable candidate for specific ELISA tests and subunit vaccine development (Naoko et al., 2002).
Abdolalizadeh et al. (2002) and Mostafaie et al. (2005) studied the reaction between immunogens of B. abortus with human, gout and rabbit sera. Mostafaie (1999) looked at the interactions between major outer membrane proteins of B. abortus (S19 strain) and B. melitensis (16M strain) with human patients sera. In another study by Mostafaie et al. (2000), the interaction between the Brucella LPS with human sera was investigated. Angel et al. (2003) have used the superoxide dismutase gene of B. abortus in order to create a DNA vaccine that was capable of inducing protective immunity in mice.
In none of these studies, the interactions of bacterial hsps with human sera were investigated. Therefore, this is probably the first report of Comparison of Heat Shock Response in Brucella abortus and Brucella melitensis and interaction of human patient sera with the hsp60 in both bacteria.
ACKNOWLEDGMENTS
The authors like to thank: Microbiology department, School of medicine, Iran university of medical science, Tehran, Iran; Cellular and molecular research center of Iran University of Medical Sciences, Tehran, Iran; Medical biology research center of Kermanshah University of Medical Sciences, Kermanshah, Iran; and Microbiology and Immunology department of Babol University of Medical Science.
REFERENCES
- Oñate, A.A., S. Cespedes, A. Cabrera, R. Rivers and A. Gonzalez et al., 2003. A DNA vaccine encoding Cu-Zn superoxide dismutase of Brucella abortus induces protective immunity in BALB/c mice. Infect. Immun., 1: 4857-4861.
CrossRefDirect Link - Baloglu, S., T.E. Toth, G.G. Schurig, N. Sriranganathan and S.M. Boyle, 2000. Humoral immune response of BALB/c mice to a vaccinia virus recombinant expressing Brucella abortus GroEL does not correlate with protection against a B. abortus challenge. Vet. Microbiol., 76: 193-199.
CrossRefPubMedDirect Link - Corbel, M.J., 1997. Brucellosis: An overview. Emerg. Infect. Dis., 3: 213-221.
CrossRefPubMedDirect Link - Gomes, A.P.T., A. Cloeckaert and M.S. Zygmunt, 2000. Characterization of heat, oxidative and acid stress responses in Brucella melitensis. Infect. Immunol., 68: 2954-2961.
Direct Link - Cassataro, J., K.A. Pasquevich, S.M. Estein, D.A. Laplagne and A. Zwerdling et al., 2007. A DNA vaccine coding for the chimera BLOSmp31 induced a better degree of protection against B. ovis and similar degree of protection against B. melitensis than review. 1 vaccination. Vaccine, 25: 5958-5967.
PubMed - Kaufmann, S.H.E., 1990. Heat shock proteins and the immune response. Immunol. Today, 11: 129-136.
CrossRef - Lin, J., L.G. Adams and T.A. Ficht, 1992. Characterization of the heat shock response in Brucella abortus and isolation of the genes the GroE heat shock proteins. Infect. Immunol., 60: 2425-2431.
PubMed - Lin, J. and T.A. Ficht, 1995. Protein synthesis in Brucella abortus induced during macrophage infection. Infect. Immunol., 63: 1409-1414.
Direct Link - Lindler, LE., T.L. Hadfield, B.D. Tall, N.J. Snellings and F.A. Rubin et al., 1996. Cloning of Brucella melitensis group 3 antigen gene encoding omp28. Infect. Immunol., 64: 2490-2499.
Direct Link - Mostafaie, A., 2005. Immunogens of Brucella abortus (S19) identified by two-dimensional gel electrophoresis and immunobloting. Iran J. Med. Sci., 30: 10-15.
Direct Link - Naoko, Y. K. Yokota, M. Mizuno, Y. Kawahara and M. Adachi et al., 2000. Antibody to heat shock protein can be used for early serological monitoring of Helicobacter pylori eradication treatment. Clin. Diag. Lab. Immunol., 7: 574-577.
Direct Link - Young, E.J., 1995. An overview of human brucellosis. Clin. Infect. Dis., 21: 283-289.
CrossRefPubMedDirect Link