
Adam Torkamanzehi
University of Sistan and Baluchistan, Zahedan, Iran
Urs Kuhnlein
Department of Animal Science, McGill University, Quebec, Canada
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 4075-4080







ABSTRACT
This study analysed mitochondrial phosphoenol-pyruate carboxykinase (PEPCK-M) gene as a candidate QTL for egg production traits in chickens. Single Strand Conformational Polymorphism (SSCP) of a 300 bp DNA fragment, from exon 9 of samples from an egg laying North American commercial White Leghorn stock, revealed a total of 6 different single strand conformers, indicative of 3 alleles. Subsequent DNA sequencing found a total of 4 base changes in this fragment between these alleles (called A1, A2 and A3) when compared to the reference sequence published online. The A1 allele had one transition mutation of T to C at position 1700. The A2 allele had accumulated three transition mutations: T to C at position 1578, A to G at position 1647 and T to C at position 1650. Transition mutation of T to C at position 1578 of the A2 allele results in the loss of an AccI site, hence, producing a de novo RFLP. Analysis of 358 female individuals from this strain showed that the population is highly polymorphic at this site. The effect of PEPCK-M genotypes at this site, namely AccI -/-, AccI +/- and AccI +/+, was tested on three traits, age at first egg, egg production rate and egg number. Least square analysis showed that exon 9 RFLP significantly affects age at first egg (p<0.05). Egg production rate and egg number traits were not affected by different genotypes at this position. The data also indicates an over-dominance effect for the associated trait.
PDF Abstract XML References Citation
How to cite this article
Adam Torkamanzehi and Urs Kuhnlein, 2007. Restriction Fragment Length and Single Strand Conformational Polymorphisms in Chicken Mitochondrial Phosphoenol-Pyruate Carboxykinase Gene and its Association with Egg Production. Pakistan Journal of Biological Sciences, 10: 4075-4080.
DOI: 10.3923/pjbs.2007.4075.4080
URL: https://scialert.net/abstract/?doi=pjbs.2007.4075.4080
DOI: 10.3923/pjbs.2007.4075.4080
URL: https://scialert.net/abstract/?doi=pjbs.2007.4075.4080
INTRODUCTION
Animal breeding is concerned mainly with the aggregate effects of all genes causing variation in the trait on interest. A complete description of a gene affecting a quantitative trait (QTL) should take into account individual properties of the genes including gene frequencies, magnitude of the gene effect and the mode of gene action. While classical Mendelian and conventional population and quantitative, genetics cannot determine individual properties of a QTL because of the intervening non-genetic variation, molecular genetics' techniques have introduced methods to assist identification of QTLs and estimation of their effects (Sing et al., 1988; Soller and Beckmann, 1988; Lande and Thompson, 1990; Falconer and Mackay, 1996; Lynch and Walsh, 1998; Darvasi, 1998). One such approach is the candidate gene approach. There are two main criteria for this approach. First, enough information must be available on the function and structure of the relevant gene. Second, the gene should potentially be associated with the character(s) in question. Once these criteria are met the gene can be searched for molecular polymorphisms and their possible associations with the phenotypes of interest. Apart from the initial benefits to better understanding of the quantitative genetics theory, identification of individual QTLs may lead to useful practical applications in animal breeding. It could improve accuracy of selection and reduce generation interval, especially when the heritability of the trait is low, or the trait is sex-limited. Several evidences of association between DNA polymorphism at candidate genes and various quantitative traits have already been reported across species (Cowan et al., 1990; Fotouhi et al., 1993; Kuhnlein et al., 1997; Mackay and Langley, 1990; Pirchner, 1988; Rothschild et al., 1995; Yao et al., 1996; Fitzpatrick et al., 2005; Nie et al., 2005).
In this study we selected mitochondrial phosphoenol-pyruate carboxykinase (PEPCK-M) as a candidate gene in chickens. PEPCK enzyme regulates a key step in de novo synthesis of glucose (Utter and Kurahashi, 1954; Nordlie and Lardy, 1963). There are two isozyme forms of the enzyme in vertebrates. A cytosolic form (PEPCK-C) and a mitochondrial form (PEPCK-M). However, both enzymes are encoded by distinct nuclear genes (Granner et al., 1983). In mammals PEPCK-C is expressed in a number of tissues, with highest expression occurring in the liver, kidney cortex and white and brown adipose tissue (Nizielski et al., 1996; Chakravarty et al., 2005). In mice, homozygous for PEPCK-C deletion (PEPCK-C -/-) becomes severely hypoglycemic by day two after birth and then dies due to profound hypoglycemia (Hakimi et al., 2005). In chicken, mitochondrial PEPCK is the sole form in the liver and constitutes 60% of the PEPCK in the kidney (Watfood et al., 1981). The isozymes of PEPCK have nearly identical catalytic properties, however, the protein structure of the two enzymes are distinct. The cDNA from chicken mitochondrial PEPCK is 3571 bp long, which codes for a 67 kDa protein. It also contains an untranslated region of 1.6 kb at the 3' end (Weldon et al., 1990). The genomic structure of the gene is still unknown, but it is suggested to be at least 16 kb long (Weldon et al., 1990; Hanson et al., 1994).
Given the important role of PEPCK in energy metabolism it could be considered a potential candidate gene for production traits, where the development and evolution of such characters are highly dependent on energy metabolism. The present study describes DNA variation at the chicken mitochondrial PEPCK gene and its association with egg production traits.
MATERIALS AND METHODS
DNA samples and the origin of chickens: DNA was extracted from chicken blood samples from strain 7 and 7R as described elsewhere (Kuhnlein et al., 1989). Strain 7 had been established by crossing 4 commercial North American White Leghorn stocks at the Center for Food and Animal Research (CFAR, Agriculture Canada) in 1958 and kept by random mating without selection with an effective population size of 457. Samples analyzed in this report were form the generation in 1993. Strain 7R is a subpopulation of strain 7 which had undergone one generation of selection for some immune traits.
PCR amplification: Sequence and position of the primers used to amplify two fragments from exon 9 of the chicken mitochondrial PEPCK are shown in Table 1. PCR reactions were carried out in 25 μL volume containing 100 ng of genomic DNA, 0.5 μM of each primer, 1x PCR buffer [10 mM Tris-HCl (pH 9.0), 1.5 mM MgCl2 and 50 mM KCl], 200 μM dNTP and 0.625 units of Thermus thermophilus (Tth) DNA Polymerase (Pharmacia). Thirty five cycles of amplification was performed at 94° Cχ 60 sec, 62° Cχ 80 sec and 72° Cχ 90 sec, after an initial denaturation at 95° C for 3 min.
Single Strand Conformation Polymorphism (SSCP): One microliter of PCR product was mixed with 15 μL of loading buffer (95% formamide, 20 mM EDTA, 0.05% bromophenol blue and 0.05 xylene cyanol), denatured at 100°C for 5 min and cooled on ice for 5 min. Samples were then loaded on 15% non-denaturig acrylamide gels [acrylamide:bis-acrylamide (49:1); 12χ10χ0.35 cm] and electrophoresed in 1x Tris-borate (pH 8.3)-EDTA buffer using a vertical minigel apparatus (Bio-Rad) for 24 h at 10 volts cm‾1. After electrophoresis at room temperature gels were separated from the glass plates and silver stained.
DNA cloning: DNA samples which showed polymorphism by SSCP were chosen for cloning and sequencing. One microliter of fresh PCR product was ligated and cloned using TA cloning kit (Invitrogen) according to the supplier’s instructions. Transformed colonies were cultured and recombinant plasmid DNA were extracted using standard methods (Sambrook et al., 1989). Presence of the inserts was confirmed by restriction enzyme digestion and agarose gel electrophoresis. Sequencing was carried out by dideoxy-chain termination method (Sanger et al., 1977) using T7 sequencing kit (Pharmacia).
Restriction Fragment Length Polymorphism (RFLP): Ten microliters of the PCR products were digested with 1.7 units of AccI endonuclease at 37°C overnight and analyzed on 2% agarose gel. Genotypes were identified by comparison of bands from digested samples with undigested and/or marker controls.
Traits and statistical analysis: Traits analyzed included age at First Egg (AFE), egg production (hen day percent), egg number at 273 to 385 days and egg number at 386 to 497 days.
Chi-square test was used to check for Hardy-Weinberg equilibrium for the exon 9 AccI -RFLP in the sample population from strain 7.
| Table 1: | Sequence and position of primers used for PCR amplification |
 | |
Association between the marker genotypes and production traits were analyzed using a one way analysis of variance linear model.
RESULTS
Single Strand Conformation Polymorphism (SSCP): Single Strand Conformation Polymorphism (SSCP) of the amplified 300 bp fragment from exon 9 of the mitochondrial PEPCK revealed a total of 3 alleles (henceforth called alleles A1, A2 and A3). However, homozygotes were only present for the two of the three alleles (A1 and A2), in a total of 72 individuals from strains 7 and 7R. The 3rd allele (A3) occurred at a much lower frequency compared to the other two and only in one of the two possible heterozygote combinations (A1A3). Both A1 and A2 alleles produced clearly distinct single strand conformers, but the band from A3 allele overlapped with the band from A2 allele, unless electrophoresis was prolonged for at least 36 h. Table 2 shows observed frequencies of the SSCP genotypes and alleles in random samples from strains 7 and 7R.
Sequence variation at exon 9: Individuals from two predominant homozygous SSCP genotypes, namely A1A1 and A2A2 were cloned and sequenced, as described before. The third SSCP allele (A3) was not sequenced, given the low frequency and unavailability of the homozygotes. A total of 4 base changes were found in the exon 9 of the mitochondrial PEPCK gene between these alleles and the published cDNA sequence (Weldon et al., 1990). The A1 allele had one transition mutation of T to C at position 1700 (Fig. 1a). This causes a mis-sense mutation, changing isoleucine to threonine at this position. It was not possible to verify this mutation as this change was not amenable by restriction enzyme digestion. The A2 allele had accumulated three transition mutations: T to C at position 1578 (Fig. 1b), A to G at position 1647 and T to C at position 1650 (Fig. 1c). Transition of T to C at position 1578 of the A2 allele results in the loss of an AccI site (changing GT/CT to GT/CC) at this position, hence creating a readily detectable RFLP (Fig. 2).
Restriction Fragment Length Polymorphism (RFLP): The 300 bp fragment from exon 9 of the mitochondrial PEPCK was extended from both ends to cover 401 bp of the region, using the second set of primers (Table 1). This fragment facilitates genotypic analysis by agarose gel electrophoresis. A total of 358 female individuals from strain 7 were amplified. PCR products were digested with AccI enzyme and genotypes were analyzed as described (Fig. 2). Table 3 gives the observed genotypic and allelic frequencies for this RFLP in strain 7. As it is clear form this table, the population is in equilibrium with respect to this RFLP.
| Table 2: | SSCP genotypic and allelic frequencies in strain 7 and 7R |
 | |
| Table 3: | AccI genotypic and allelic frequencies in strain 7 |
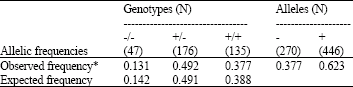 | |
| *: X2[d. f. = 1] = 0.76 (p = 0.40) | |
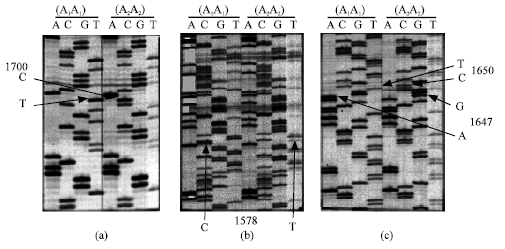 | |
| Fig. 1: | The sequencing gels, after autoradiography, showing the four base changes in the 300 bp fragment from the exon 9 of the mitochondrial PEPCK gene. (a) Transition mutation of T to C at position 1700 in the A1 allele. (b) Transition mutation of T to C at position 1578 in the A2 allele. This mutation results in the loss of an AccI site (changing GT/CT to GT/CC) and creates a readily detectable RFLP. (c) Transition mutation of T to C at position 1650 and A to G at position 1647 in the A2 allele (c) |
| Table 4: | Comparison of least square means for egg production traits from different genotypes |
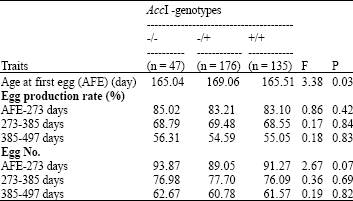 | |
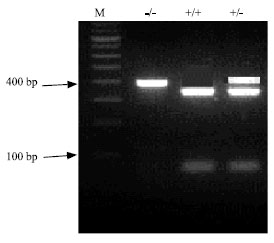 | |
| Fig. 2: | PCR products of the 401 bp fragment from the exon 9 of chicken PEPCK-M after digestion with 1.7 units of AccI enzyme and electrophoresis on a 2% agarose gel. M shows bands from a 100 bp DNA marker |
Trait association: The effect of PEPCK genotypes (-/-, +/- and +/+) was tested on three traits in strain 7-age at first egg, egg production rate and egg number-all of which affect egg production in chickens. Egg production and egg number traits each consisted of 3 records at different laying intervals. Table 4 compares least square means for these traits. Analysis showed that exon 9 RFLP significantly affects age at first egg. Apart from a nearly significant difference (p = 0.07) in egg number in the period of age at first egg and 273 days, egg production rate and egg number traits were not affected by different genotypes at this position.
DISCUSSION
Numerous reports have shown association between polymorphisms at growth hormone, growth hormone receptor, cytosolic PEPCK and several other candidate genes, with production or disease resistance traits in chickens (Fotouhi et al., 1993; Kuhnlein et al., 1997; Parsanejad et al., 2002, 2003; Nie et al., 2005; Dunn et al., 2004).
Taking the chicken mitochondrial PEPCK as another candidate gene we present evidence of a rapidly detectable DNA variation in this gene and association with egg production in an egg layer strain of White Leghorn.
SSCP analysis the 300 bp fragment from exon 9 revealed 3 alleles with only two of the alleles (A1 and A2) observed in heterozygote and homozygote forms. The 3rd allele (A3) was only observed as heterozygotes with A1 (the most common allele). Given the low frequency of this allele more samples have to be analysed in order to observe possible homozygotes for this allele.
From the four base changes, revealed by sequencing in the exon 9 of the mitochondrial PEPCK gene, a transition of T to C at position 1578 of the A2 allele creates a de novo AccI site, which leads to an easily detectable RFLP (Fig. 1 and 2). Similarly, another transition (T to C) at position 1700 of the A1 allele causes a mis-sense mutation, changing isoleucine to threonine. This variation was not amenable by restriction enzyme digestion, therefore further clarification of this mutation is necessary.
RFLP analysis of female individuals form strain 7 showed that the AccI genotypes are in Hardy-Weinberg equilibrium (Table 3). Furthermore, both alleles are maintained at intermediate frequencies, an indication of the absence of natural selection or selection in favour of heterozygotes. Further elucidation of these matters might seem useful, as the latter, if present, may indicate heterosis for fitness or fitness associated traits.
Age at first egg, an egg production trait, was significantly affected by DNA variation at exon 9 (Table 4). Egg number had also a near significant association. This was expected as the two traits are negatively correlated. There is an indication of an over-dominance effect for both associated traits, which may be due to heterosis, as a result of natural selection favouring heterozygotes. It should be interesting to see if there is any indication of genetic homeostasis (Lerner, 1954) for these traits while the newly formed strain 7 Leghorn population has undergone relaxation of selection.
The cytosolic form of the PEPCK gene (PEPCK-C), carries out the same reaction as the PEPCK-M but is encoded by a different nuclear gene. In a different study (Parsanejad et al., 2003) we tested for the interaction between the two genes. The three AccI -RFLP genotypes were analyzed together with six PEPCK-C genotypes described previously (Parsanejad et al., 2002). Analysis indicated that the PEPCK-M genotype affected the trait egg weight among some of the PEPCK-C genotypes. There are also evidence of coselection of a PEPCK-M MspI-RFLP with Marek’s Disease (MD) resistance in White Leghorn chickens (Li et al., 1998).
Research is in progress to document further variation at mitochondrial PEPCK gene and association with other traits in chicken, including growth and disease resistance traits. Moreover, dissecting DNA variation at other coding and non-conding regions of the gene and analysis of their association with economically important QTL will help to determine if the PEPCK is indeed a QTL and/or its effect is large enough to be included in the indices for selecting the relevant trait(s).
The future for understanding the complex genetics of quantitative traits is becoming bright through ever increasing molecular genetics techniques, which allow identification of various genes right down to the coding sequence (Buitenhuis et al., 2005; Korstanje and Paigen, 2002). Some of these genes might be potential QTLs and therefore, could readily be analysed by the candidate gene approach for association with economically important traits in animals. Emergence of new information about the QTLs will increase our understanding of the nature of genetic variation in economically important traits and will help in its use in the most efficient, effective and sustainable way (Dunn et al., 2004; Nei et al., 2005).
ACKNOWLEDGMENTS
We thank D. Zadworney, S. Aggrey, L. Yao and Leny Volkov for valuable guidance in data analysis and molecular techniques. A. Torkamanzehi is a visiting scientist from the University of Sistan and Baluchistan, Zahedan, Iran. This research was supported by a grant to U. Kuhnlien.
REFERENCES
- Buitenhuis, A.J., T.B. Rodenburg, M. Siwek, S.J. Cornelissen and M.G. Nieuwland et al., 2004. Identification of QTLs involved in open field behavior in young and adult laying hens. Beha. Genet., 34: 325-333.
Direct Link - Chakravarty, K., H. Cassuto, L. Reshef and R.W. Hanson, 2005. Factors that control the tissue-specific transcription of the gene for phosphoenolpyruvate carboxykinase-C. Crit. Rev. Biochem. Mol. Biol., 3: 129-154.
Direct Link - Cowan, C.M., M.R. Dentine, R.L. Ax and L.A. Schuler, 1990. Structural variation around prolactin gene linked to quantitative traits in a elite Holstein sire family. Theor. Applied Genet., 79: 577-582.
Direct Link - Darvasi, A., 1998. Experimental strategies for the genetic dissection of complex traits in animal models. Nat. Genet., 18: 19-24.
Direct Link - Dunn, I.C., Y.W. Miao, A. Morris, M.N. Romanov, P.W. Wilson and D. Waddington, 2004. A study of association between genetic markers in candidate genes and reproductive traits in one generation of a commercial broiler breeder hen population. Heredity, 92: 128-134.
CrossRefPubMedDirect Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Fitzpatrick, M.J., Y. Ben-Shahar, H.M. Smid, L.E. Vet, G.E. Robinson and M.B. Sokolowski, 2005. Candidate genes for behavioural ecology. Trends Ecol. Evol., 20: 96-104.
Direct Link - Fotouhi, N., C.N. Karatzas, U. Kuhnlein and D. Zadworny, 1993. Identification of growth hormone DNA polymorphisms which respond to divergent selection for abdominal fat content in chickens. Theor. Applied Genet., 85: 931-936.
Direct Link - Hakimi, P., M.T. Johnson, J. Yang, D.F. Lepage and R.A. Conlon et al., 2005. Phosphoenolpyruvate carboxykinase and the critical role of cataplerosis in the control of hepatic metabolism. Nutr. Metabol., 2: 33-44.
Direct Link - Hanson, R.W. and Y.M. Patel, 1994. P-enolpyruate carboxykinase: The gene and the enzyme. Adv. Enzymol. Relat. Areas Mol. Biol., 69: 203-281.
Direct Link - Korstanje, R. and B. Paigen, 2002. From QTL to gene: The harvest begins. Nat. Genet., 31: 235-236.
Direct Link - Kuhnlein, U., L. Ni, S. Weigend, J.S. Gavora, W. Fairfull and D. Zadworny, 1997. DNA polymorphism in the chicken growth hormone gene: Response to selection for disease resistance and association with egg production. Anim. Genet., 28: 116-123.
Direct Link - Lande, R. and R. Thompson, 1990. Efficiency of marker-assisted selection in the improvement of quantitative traits. Genetics, 124: 743-756.
Direct Link - Li, S., S.E. Aggrey, D. Zadworny, W. Fairfull and U. Kuhnlein, 1998. Evidence for a genetic variation in the mitochondrial genome affecting traits in White Leghorn chickens. J. Heredity, 89: 222-226.
Direct Link - Nie, Q., B. Sun, D. Zhang, C. Luo and N.A. Ishag et al., 2005. High diversity of the chicken growth hormone gene and effects on growth and carcass traits. J. Heredity, 96: 698-703.
CrossRefDirect Link - Nizielski, S.E., P.S. Lechner, C.M. Croniger, N.D. Wang, G.L. Darlington and R.W. Hanson, 1996. Animal models for studying the genetic basis of metabolic regulation. J. Nutr., 126: 2697-2708.
Direct Link - Nordlie, R.C. and H.A. Lardy, 1963. Mammalian liver phosphoenolpyruate carboxykinase activities. J. Biol. Chem., 238: 2259-2263.
Direct Link - Parsanejad, R., D. Zadworny and U. Kuhnlein, 2002. Genetic variability of the cytosolic phosphoenolpyruvate carboxykinase gene in white leghorn chickens. Poult. Sci., 81: 1668-1670.
Direct Link - Parsanejad, R., A. Torkamanzehi, D. Zadworny and U. Kuhnlein, 2003. Alleles of cytosolic phosphoenolpyruvate carboxykinase (PEPCK): Trait association and interaction with mitochondrial PEPCK in a strain of White Leghorn chickens. Poult. Sci., 82: 1708-1715.
Direct Link - Rothschild, M., C. Jacobson, D. Vaske, C. Tullge and L. Wang et al., 1996. The estrogen receptor locus is associated with a major gene influencing litter size in pigs. Proc. Natl. Acad. Sci. USA., 93: 201-205.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sanger, F., S. Nicklen and A.R. Coulson, 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A., 74: 5463-5467.
CrossRefPubMedDirect Link - Watfood, M., Y. Hod, Y.B. Chiao, M.F. Utter and R.W. Hanson, 1981. The unique role of the kidney in gluconeogenesis in the chicken. J. Biol. Chem., 256: 10023-10027.
Direct Link - Weldon, S.L., A. Rando, A.S. Matathias, Y. Hod and P.A. Kalonick et al., 1990. Mitochondrial Phosphoenol pyruate carboxykinase from the chicken. J. Biol. Chem., 265: 7308-7317.
Direct Link - Yao, J., S.E. Aggrey, D. Zadworney, J.F. Hayes and U. Kuhnlein, 1996. Sequence variations in the bovine growth hormone gene characterized by single-strand conformation polymorphism (SSCP) analysis and their association with milk production traits in Holsteins. Genetics, 144: 1809-1816.
PubMedDirect Link