
F. Chamandosti
Department of Biology, Faculty of Science, Teacher Training University of Tehran. No. 49, Mofatteh Ave, Tehran, Iran
A. Majd
Department of Biology, Faculty of Science, Teacher Training University of Tehran. No. 49, Mofatteh Ave, Tehran, Iran
S. Mehrabian
Department of Biology, Faculty of Science, Teacher Training University of Tehran. No. 49, Mofatteh Ave, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 2 | Page No.: 302-306







ABSTRACT
In this study a simple and efficient procedure was developed for inducing callus and plant regeneration using hypocotyl segmsents of Canola (Brassica napus L.). The combinations and concentrations of differents plant growth regulators had a critical effect for callus formation with various colour and potential of differentiation. Three morphologically distinct types of calli were induced: 1) yellow calli were induced on MS medium supplemented with variou concentrations of 2, 4-D and 2 mg L-1 BA and presence of somatic embryos in first stages of development was showed by histological studies in these calli. white calli were induced on MS medium supplemented with into 1-2 mg L-1 IBA and 1 mg L-1 BA and MS medium with only 2 mg L-1 BA. These calli were organogen. So that differentiated to shoots, then shoots rooted and whole plants were obtained. And dark brown calli that induced on media with out of 2 mg L-1 IBA. These calli died and did not differentiated.
PDF Abstract XML References Citation
How to cite this article
F. Chamandosti, A. Majd and S. Mehrabian, 2006. In vitro Plant Regeneration from Callus of Cotyledons in Canola (Brassica napus L.). Pakistan Journal of Biological Sciences, 9: 302-306.
DOI: 10.3923/pjbs.2006.302.306
URL: https://scialert.net/abstract/?doi=pjbs.2006.302.306
DOI: 10.3923/pjbs.2006.302.306
URL: https://scialert.net/abstract/?doi=pjbs.2006.302.306
INTRODUCTION
Brassica crop species and their allies are important source of edible roots, stems, leaves, buds and inflorescences, as well as of edible or industerial oil, condiments and forage (Gomez-Campo, 1999). Brassica napus L. is one of the three major oilseed Brassica species that cultivated in Iran due to having edible and agricultural desirable specification, therefor cultivation of this important and economic crop has been increased in recent decade in Iran. Plant genetic transformation and gene cloning are becoming important tools in crop improvement. However development of efficient and reproducible tissue culture regeneration protocol is the first step in utilizing the power and potential of this new technology (Vencatachalam et al., 1999). A vegetative propagation system based on adventitious shoot production could be integrated with genetic engineering (Bergmann and Moon, 1997). Using the hypocotyl regeneration system is a simple and rapid method for production of Canola via tissue cultuer. Microspore culture and plant regeneration of Brassica napus L. this important economic crop, has been investigated extensively because of its high frequency of androgenesis from isolated late unicnucleate microspors and early binuceate pollen .culture protocol by this method have been progressively optimized over many years (Tian et al., 2004). In this case somatic embryogenesis take place from embrypgenically predetermined cells (von Arnold et al., 2002). But in this study a simple and rapid method was developed for inducing callus and plant regeneration in Canola.
MATERIALS AND METHODS
This study conducted in Developmental Biology Laboratory in Faculty of Sceince, Teacher Training University, Tehran, Iran in October 2004-August 2005.
Seed germination and explant preparation: Mature seeds of Brassica napus L. were obtained from Oilseed Research and development Company. Iran. They were surface- sterilized in commerical sodium hypochlorite solution (5% available chlorine) for 8 min, followed by four to five times rinses with sterile distilled water and then germinated on MS medium (Murashige and Skoog, 1962) free from plant growth regulators under 16 photoperiod at 200 μm mol–2s–1. Hypocotyl segments 2-5 mm in length were excised from 14-days-old aseptically grown green seedling for preparation of explants.
Media and culture conditions: The basal medium tested in these experiments were MS, 0.5 MS (MS salts, MS vitamins) and B5 (B5 salts, B5 vitamins). All of media containing 3% sucrose and 0.1% agar and supplemented with combinations of auxins (2, 4-D and IBA) and cytokinin (BA). MS medium free from PGRs was also used as control. All of media were mixed and adjusted to pH 5.8 prior to autoclaving at 120°C for 20 min. All cultures incubated at 24±2°C in a incubator under dark condition and 200 μm mol–2 s–1 for callus induction and growth of shoot regenerated, respectively.
Callus induction: Hypocotyl segments were placed on MS, 0.5 MS and B5 media with various concentrations of 2, 4-D (2 -10 mg L–1) with 2 mg L–1 BA. 1-4 mg L–1 IBA plus 1 mg L–1 BA and 2 mg L–1 BA alone also were used. All of media complemented with 500 mg L–1 casein hydrolyset. After 20 days presence of callus (yes or no) and general quality (based on colour and organogenesis or embryogenesis) were recorded.
Shoot formation and plant regeneration: The proliferated white calli after approximately 1 month of culture shooted on MS media supplemented with 1 mg L–1 IBA plus 1 mg L–1 BA , 2 mg L–1 BA and 2 mg L–1 IBA with 1 mg L–1 BA. (previous media for induction of white calli) These shoots were transfered to MS medium without PGRs and with NAA at 1-2 mg L–1 and IBA at 1-2 mg L–1 for rooting. The percentage of rooted shoots was scored 20 days of culturing.
Light microscopy: Calli were fixed in FAA (formalin: glacial acetic acid: ethanol, (10: 5: 35) for 16 h, dehydrated in a grated series of alcohol where after they were embedded in paraffin wax. Serial sections 8 mm thick were cut and stained with hematoxylin and eosin (Martoja and Martoja, 1967).
Exprimental design, data collection and analysis: Expriments were set up in Completely Randomized Design and repeated three times. Each treatment has 20 replications. Observation on the number of explants forming shoots were recorded. Data were subjected to SD and ANOVA test.
RESULTS
Callus induction: Callus formation from ends of hypocotyls segments after two weeks of culture was visible. After 20 days in culture explants developed into three types of calli which differed in colour and potential of organogenesis. Despites of basal salts and streght of media, concentration and kind of PGRs play an important role in induction of these three types of calli. In fact all three basal media showed approximately equal effect on induction of three morphologically distinct types of calli (data not shown). White calli was only obtained in media with 1-2 mg L–1 IBA plus 1 mg L–1 BA and 2 mg L–1 BA alone. Yellow calli was only induced i1n media with various concentrations of 2,4-D with 2 mg L–1 BA. Histological studies presence of somatic embryos confirmed in medium with 10 mg L–1 2, 4-D (Fig. 1 and 2). In media with out of 2 mg L–1 IBA all of calli were dark brown and practically died.
Shoot formation: As previously mentioned proliferated calli of the three types divided based on colour and potential for shoot and root formation. The effects of both types of calli (white and yellow, dark brown calli in initial steps of experiments died) and media on morphogenic responses were significant. White calli that produced on MS media with 1mg L–1 IBA and 1 mg L–1 BA and 2 mg L–1 BA alone and 2 mg L–1 IBA with 1 mg L–1 BA showed a high frequency of shoot regeneration (Fig. 3 and 4). Within the second of subculture small, light patches emerged on the surface of white calli, these developed further into multiple shoots after 14 days of culture. As Table 1 shows, in medium with 2 mg L–1 BA the means number of shoots/explants was the heighest (89±0.27%). Also in media with 1 mg L–1 IBA and 1 mg L–1 BA and 2 mg L–1 IBA and 1 mg L–1 BA the means number of shoots/explants was good (56±0.16% and 58±0.19%, respectively). This table also explains that in media with out of 2 mg L–1 IBA and 1 mg L–1 BA no shoot formation was seen and as we described previously in these media formed dark brown calli. In medium with 6 mg L–1 2, 4-D and 2 mg L–1 BA some parts of yellow calli produced a very small leaves that accounted as shoot with least percentage (0.15±0.013).
| Table 1: | Caulogenic respons of hypocotyl explants of B. napus cultured on MS medium |
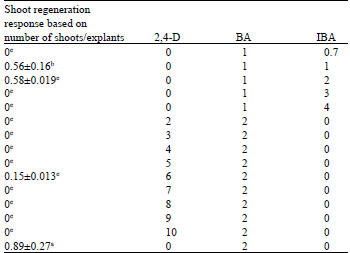 | |
Means±SD; means for each experiment marked with same letter do not differ significantly (p<0.01) | |
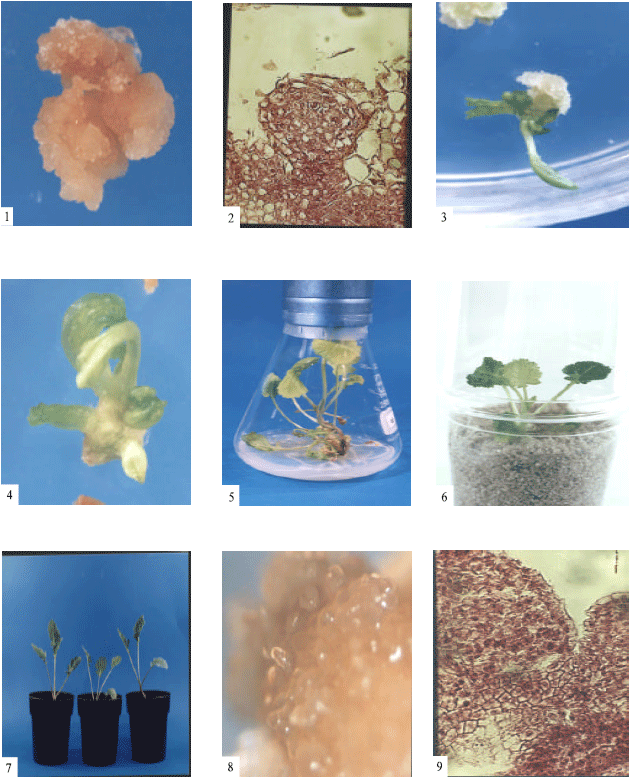 | |
| Fig. 1- 9: | Callus induction and plant regeneration in Brassica napus L. 1: Yellow calli that induced in medium with 10 mg L–1 2,4-D and 2 mg L–1 BA, 2: Longitudinal section of embryo on medium with 10 mg L–1 2,4-D and 2 mg L–1 BA at globular stage (bar200X), 3: Shoot formation from white calli on medium containing 1 mg L–1 IBA and 1 mg L–1 BA, 4: Shoot formation from white calli on medium containing 2 mg L–1 BA, 5: Rooted adventitious shoots (in medium with 2 mg L–1 BA) after 2- weeks cultured on MS medium with 1 mg L–1 IBA. 6 and 7: In vitro raised plants established in soil, 8: Elongated finger like projection was produced on callus surface that obtained in medium with 2 mg L–1 BA (arrowheads), 9: Longitudinal section of elongated finger like (bar400X) |
Rooting and plant regeneration: Regenerated shoots, were transferred to MS media with various concentrations of NAA and IBA and medium lacking growth regulator to evaluate their potential for root formation. Only in medium with 1 mg L–1 IBA rooting was observed. In fact in this medium regenerated shoots displayed the shortest time of root initiation and the hightest frequency of shoots with rooting (90%). Two weeks after transfer regenerated shoots produced 8-9 root and whole plantlets were obtaine Whole plantlets adapted to dry environment and successfully poted (Fig. 5-7).
DISCUSSION
Our results indicate that presence of 2,4-D in media produces yelow calli. There are many reports that show these type of calli are embryogenic based on microscopic observations and their regeneration capacity. (Kebebew et al., 1998; Zhang et al., 2001; Salunkhe et al., 1999; Luo et al., 2001; Neumann et al., 2000; Selles et al., 1999; Lee et al., 2002). Somatic embryogenesis includes several steps, first: initiation of embryogenic culturs by culturing the primery explant on medium supplemented with PGRs mainly auxin but often also cytokinin (von Arnold et al., 2002). Also in our expriments yellow calli did not changed their morphological characters even though they were maintained in culture for 5 months. On the other hand did not produce regenerated shoots and roots. And microscopic observations proved presence of structural embryonic. Calli induced in media (1-2 mg L–1 IBA and 1 mg L–1 BA) and 2 mg L–1 BA differentiated to shoots. Means of shoots regenerated numbers and streght of them in medium with 2 mg L–1 BA was higher. Dan and Reichert (1998) reported that hypocotyl explants excised from 7-days old seedlings after placement on a medium containing 5-10 μM bensyladenin produced adventitious shoots. Like Rudraswamy and Reichert (1998) explained that the numbers of microshoots produced per explant appeared more correlated to the concentration of BA in medium.
It is interesting that a few elongated finger-like projections could be observed on callus surface that obtained in medium with 2 mg L–1 BA. Histological studies showed that the somatic embryos had distinct developmental stages, suggesting that shoots were regenerated through or partly through somatic embryogenesis (Fig. 8 and 9) (Luo and Jia, 1998).
In this study we found that Brassica napus L. have potential of somatic embryogenesis, callus formation and regeneration of organs (shoot and root). Therefore we can create somaclonal variation in this important crop simplicity. Somaclonal variation has been reported to occur for a range of agronomic traits such as yield, protein content, salt tolerance and herbicide and disease resistane (Donovan et al., 1994). Somaclonal variation in Brassica napus L. need more investigations and have special importance (Sacristan, 1982).
REFERENCES
- Bergmann, B.A. and H.K. Moon, 1997. In vitro adventitious shoot production in Paulownia. Plant Cell Rep., 16: 315-319.
CrossRefDirect Link - Zhang, B.H., R. Feng, F. Liu and Q. Wang, 2001. High frequency somatic embryogenesis and plant regeneration of an elite Chinese cotton variety. Botanical Bull. Acada. Sin., 42: 9-16.
Direct Link - Donovan, A.M., R. Morgan, C. Valobra-Piagnani, M.S. Ridout, D.J. James and C.M.E. Garrett, 1994. Assessment of somaclonal variation in apple. I. Resistance to the fire blight pathogen, Erwinia amylovora. J. Hortic. Sci., 69: 105-113.
CrossRefDirect Link - Dan, Y. and N.A. Reichert, 1998. Organogenic regeneration of Soybean from Hypocotyl explants and progeny analysis. In Vitro Cell. Dev. Biol.-Plant, 34: 14-21.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Tian, H., C.Y. Yao and M.X. Sun, 2004. High frequency conversion of microspore-derived embryos of Brassica napus cv. Topas by supplemented calcium and vitamins. Plant Cell. Tissue Organ Cult., 76: 159-165.
CrossRefDirect Link - Von Arnold, S., I. Sabala, P. Bozhkov, J. Dyachok and L. Filonova, 2002. Developmental pathways of somatic embryogenesis. Plant Cell. Tissue Organ Cult., 69: 233-249.
CrossRefDirect Link