
A. M. Diab
Department of Botany, Microbiology Section, Faculty of Science, Suez Canal University, Ismailia, Egypt
M.H. Abdel Aziz
Department of Botany, Microbiology Section, Faculty of Science, Suez Canal University, Ismailia, Egypt
S. A. Selim
Department of Botany, Microbiology Section, Faculty of Science, Suez Canal University, Ismailia, Egypt
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 7 | Page No.: 774-779







ABSTRACT
Thirteen multiple antibiotic resistant (MAR) gram-negative bacterial isolates from drinking water of Ismailia city were examined for the presence of plasmid DNA. Only six isolates were found to be plasmid-bearing isolates. One of these isolates, Aeromonas hydrophila, found to contain three plasmids with molecular sizes of 21.226, 3.530 and 1.375Kbp. Other isolates belonged to the genera Escherichia, Enterobacter (2 isolates), Pseudomonas and Salmonella which were found to contain only one plasmid each with varying molecular sizes. Curing and transformation protocols indicated that the ampicillin resistance of these isolates is plasmid-linked. Transformation coefficient, resistance expression within E. coli HB 101 and DH1 as well as restriction analysis of the plasmids were achieved.
PDF Abstract XML References Citation
How to cite this article
A. M. Diab, M.H. Abdel Aziz and S. A. Selim, 2002. Plasmid-Encoded Transferable Antibiotic Resistance in Gram-negative Bacteria, Isolated from Drinking Water in Ismailia City. Pakistan Journal of Biological Sciences, 5: 774-779.
DOI: 10.3923/pjbs.2002.774.779
URL: https://scialert.net/abstract/?doi=pjbs.2002.774.779
DOI: 10.3923/pjbs.2002.774.779
URL: https://scialert.net/abstract/?doi=pjbs.2002.774.779
INTRODUCTION
Antibiotic-resistant bacteria and antibiotics are discharged in various amounts in the environment as a result of the increasing and often indiscriminate use of antibiotics in medical, veterinary, and agricultural practices. River water is the main receptacle for these pollutants, as rivers are one of the major sources of water for human and animal consumption, this pollution may contribute to the maintenance and even the spread of bacterial antibiotic resistance (Halling-Sorensen et al., 1998 and Goni- Urriza et al., 2000). Most antibiotic- resistant microbes emerge as a result of genetic change and subsequent selection processes by antibiotics. The resistance factor may be chromosomal, that developed as a result of spontaneous mutations and extrachromosomal resistance (plasmid resistance). R factors are a class of plasmids that carry genes for resistance to one and often several antibiotics and heavy metals (Bibb, 1981; Fass and Barnishan, 1981; Richardson, 1982; Niemi et al., 1983; Smith and Amyer, 1984; Chang and Bolton, 1987; Sandt and Herson, 1991; Pathak and Gopal, 1994 and Brooks et al., 1995). Genetic material and plasmids in particular, can be transferred by transduction, transformation, conjugation and transposition mechanisms (Davies et al., 1980). The health concerns that antibiotic resistant (AR) and multiple-antibiotic resistant (MAR) bacteria present are amplified by the phenomenon of resistance transfer (Mckeon et al., 1995). Several studies showed that antibiotic resistance characteristics can be transferred to sensitive recipient organisms in the environment and DNA coding for antibiotic resistance may be conjugally transferred between similar microorganisms (Shaw and Cabelli, 1980; Ramteke et al., 1990 and Harnett et al., 1998). Thus an antibiotic-resistant bacterium in drinking water poses a serious public health problem.
The purpose of this study was to investigate any relation between the previously proved antibiotic-resistance of many gram-negative bacteria isolated from drinking water in Ismailia city (Diab et al., 2000) and the presence of plasmids. The contribution degree of these plasmids, if there is any, in antibiotic-resistance and the characters of these plasmids including transferability and expression capacity were also targeted.
MATERIALS AND METHODS
Bacterial isolates, plasmids and phage DNA: The bacterial isolates, plasmids and phages DNA used in this study and their sources are listed in Table 1.
Isolation of Plasmid DNA: LB broth plus appropriate antibiotics in culture tubes with individual bacterial colonies were shaken at 37oC overnight. Cell pellets are resuspended in 100μL alkaline extraction with solution I composed of 50mM glucose, 25mM tris (pH 8.0) and 10mM.
| Table 1: | Bacterial isolates, plasmids, phage DNA and their sources |
 | |
| * Isolated, identified and confirmed as AR and/or MAR. (Diab et al., 2000) | |
EDTA, combined with fresh 20% SDS, solution II by combining 1mL of 2M NaOH, 0.5mL 20% SDS, and 8.5mL H2O. One hundred and fiftyμL of ice-cold solution III [3M KOAC were added, brought to pH 5.5 by adding glacial acetic acid] to each tube. A white precipitate of denatured proteins and cell debris is formed. Two hundredμL phase separation mixture of phenol chloroform (1:1) was added to each tube. Closed microfuge tubes were inverted several times to mix the phases well. The samples were centrifuged for 1minute in a microfuge. Using a pipett or a Pasteur pipet, the upper (aqueous) phase was transferred to new microfuge tubes. Precipitated the DNA using 300μl cold isopropanol to each tube. Fresh TE [50mM Tris (pH 8.0); 20mM NaCl and 5mM EDTA] plus digestion of RNA using RNase by adding 20μL (5 mg/mL) RNase in TE buffer was prepared. Five μL of 3M NaOAC and 125μL cold ethanol to DNA were added. Samples can be stored at –20 oC indefinitely. The pellets were resuspended in 40μL of 1mM Tris and 0.1mM EDTA, pH 7.5 (Sambrook et al., 1989).
Concentrations and purity of DNA: Concentrations and purity of DNA were estimated spectrophotometrically using Spectro-22, Labo.Med. Inc., USA. The concentration of DNA was determined at 260nm. The purity of DNA was calculated by the ratio of absorbance at 260nm and 280nm (Sambrook et al., 1989).
Agarose gel electrophoresis of the plasmid DNA: Agarose gel electrophoresis was carried out using the tris-borate EDTA buffer (TBE). Gels were prepared by adding 1% agarose and 5μL ethidium bromide (stock solution of 10mg/mL) to TBE buffer as described by Hammad and Dora (1993).
Restriction analysis: Pure DNA sample (3FL) was added to 12FL deionized water, 1FL endonuclease, and 2FL of the high saltdigestion buffer 1M NaCl, 500mM Tris-HCl (pH 7.5). 100mM MgCl2.6H2O, and 10mM DTT loading buffer (5FL) was added and agarose gel was run at 90V for 2h. A control plasmid sample and a reference lambda plasmid treated with Hind III, were run in the same gel (Sambrook et al., 1989). Hind III (Sigma Production, St. Louis, USA), EcoR I and BamH I (Roche Diagnostics GmbH, Mannheim, Germany) were used.
Curing and transformation protocols: Serial dilutions of acridine orange were used for curing (Ramteke et al., 1990).Two different ampicillin-senstive and plasmid-free E. coli strains; DH1 and HB 101 were used as recipient cells in the transformation experiments using chlorides mixture method (Sambrook et al., 1989).
RESULTS
Plasmid profiles of the gram-negative bacterial isolates under study were determined. Only 6 out of 13 isolates were found to contain plasmids (Table 2 and Fig. 1).
| Table 2: | Gram-negative bacterial isolates screened for the presence of plasmids. |
 | |
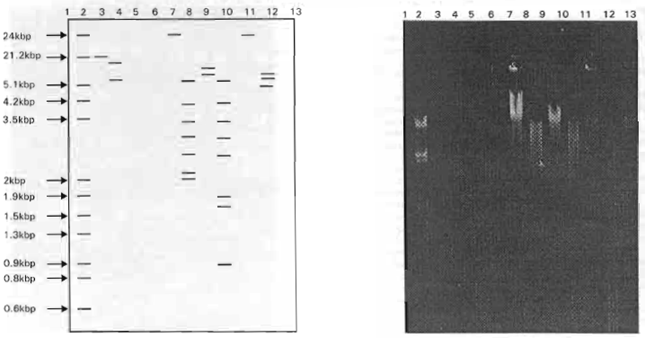
No plasmids could be detected for other seven isolates (Fig. 1). One isolate Aeromonas hydrophila was found to harbor three plasmids (multiplamidic). Molecular sizes of the detected plasmids ranged from 1.3kbp in Aeromonas hydrophila to 24kbp in Pseudomonas aeruginosa (Fig. 2). Concentration and degree of purity of the plasmid DNA(S) were as in Table 3.
Plasmid curing in combination with MIC determination revealed that 100% of the isolates resistance to ampicillin is plasmid linked except for Pseudomonas aeruginosa and Salmonella choleraesuis where 25% and 50% of their resistance, respectively, were found chromosomal. E. coli DH1 that is plasmid-free and ampicillin sensitive strain, transformed with each of the isolated plasmids, in separate trails while E. coli HB 101 strain was not. Strain DH1, received the plasmid, and expressed the ampicillin resistance to MIC(S) very close to that of the original donor isolates except for two species Pseudomonas aeruginosa and Salmonella choleraesuis (Table 3). Althuogh the plasmids showed transformation coefficients ranging from 0.0009 for isolate Salmonella choleraesuis to 1.2 for isolate Enterobacter aerogenes (Table 3) yet they were unstable.
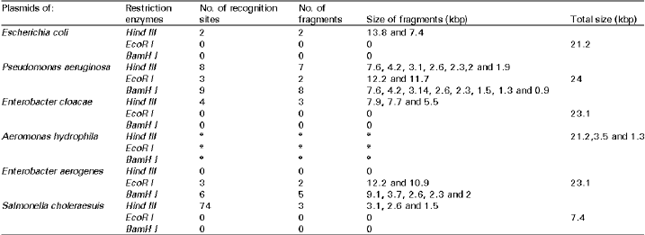
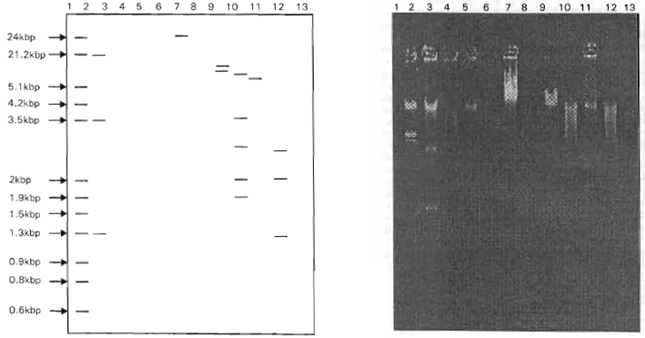
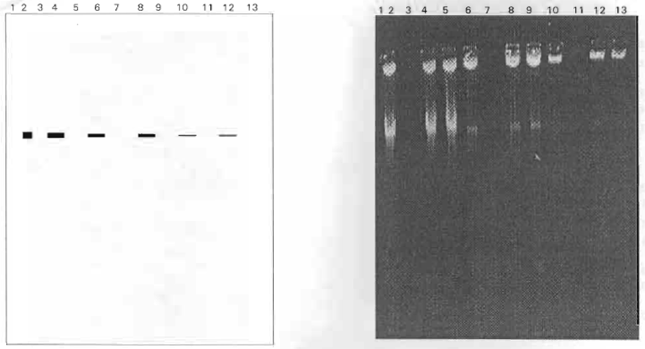
Digestion of different isolated plasmids, singly, with Hind III, EcoR I and BamHI are given in Figs. 3, 4, 5 and 6. Number of recognition sites, number of fragments and the approximate molecular size of restricted fragments are shown in Table 4.
DISCUSSION
The plasmid-linked resistance patterns of 13 gram negative ampicillin-resistant isolates were investigated. For six of them plasmids were observed and plasimd-linked resistance to ampicillin were confirmed with relatively high MIC(S). In recent studies plasimd-linked resistance, especially for pathogenic bacterial isolates, are still of critical importance (Bekowitz, 1995; Demain and Davies, 1999).
| Table 3: | Characterization of isolated plasmids including size, concentration, purity, transformation coefficient to E. coli DH 1, MIC(S) of the original and transformed isolates and curing (%). |
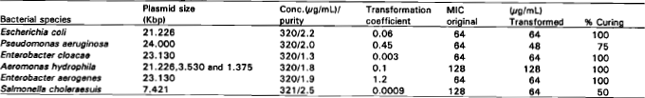 | |
| Table 4: | The restriction patterns of plasmids from 6 Gram negative bacterial isolates isolated from drinking water in Ismailia city and digested with Hind III, Ecor I and BamH I. |
 | |
| * Not Determined | |
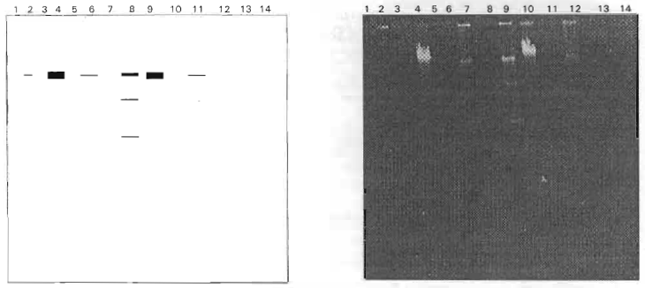 | |
| Fig. 1: | A brief description of the plasmids detected in studied bacterial isolates. Where: Lane 2: Escherichia coli* Lane 3: Acinetobater calcoaceticus Lane 4: Pseudomonas aeruginosa Lane 5: Citrobacter freundii Lane 6: Enterobacter cloacae* Lane 7: Hafinia alvei Lane 8: Aeromonas hydrophilla* Lane 9: Enterobacter aerogenes Lane 10: Morganella morganii Lane 11: Salmonella choleraesuis Lane 12: Citrobacter diversus Lane 13: Proteus vulgaris Lane 14: Proteus mirabilis * = plasmid bearing isolates |
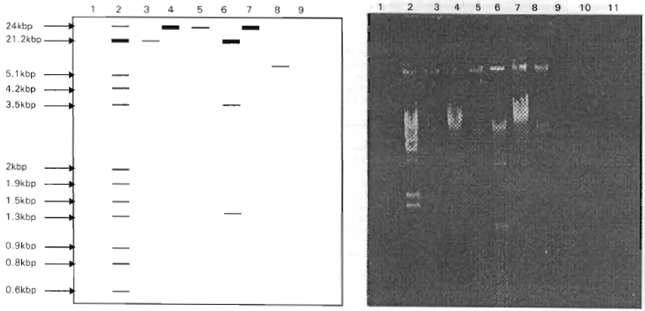 | |
| Fig. 2: | Molecular size of recovered plasmids against lambada (λ) DNA digested with Hind III+ EcoR I; as a reference, lane 2 pure plasmids preparation from Escherichia colil, lane 3, from Pseudomonas aeruginosa , lane 4, from Enterobacter cloacae, lane 5, from Aeromonas hydrophila, lane 6, from Enterobacter aerogenes and lane 7, from Salmonella choleraesuis, lane 8. |
 | |
| Fig. 3: | The restriction patterns of bacterial plasmids isolated from drinking water in Ismailia city and digested with Hind III, Ecor I and BamH I respectively, Where: Lane 2: Lambda (λ) DNA + Hind III + EcorR I Lane 3: Escherichia coli Lane 7 : Pseudomonas aeruginosa Lane 11: Enterobacter cloacae |
 | |
| Fig. 4: | The restriction patterns of bacterial plasmids isolatd from drinking water in Ismailia city and digested with Hind III, EcoR I and BamH I respectively. Where: Lane 2: Lambda (λ) DNA + Hind III + EcoR I Lane 3: Aeromonas hydrophila Lane 7: Enterobacter aerogenes Lane 11: Salmonella choleraesuis |
 | |
| Fig. 5: | The original pure plasmids from isolates Escherichia coli, Psedomonas aeruginosa and Enterobacter cloacae as a control, lanes 2,6 and 10, the same isolates cured from plasmids, lanes 3,7 and 11 and transformed plasmids with E. coli DH 1 of Escherichia coli, Pseudomonas aeruginosa and Enterobacter cloacae, lanes 4, 8 and 12 respectively. |
 | |
| Fig. 6: | The original pure plasmids from isolates Escherichia coli, Pseudomonas eaeruginosa and Enterobacter cloacae as a control, lanes2, 6 and 10, the same isolates cured from plasmids, lanes 3,7 and 11 and transformed plasmids with E. coli DH 1 of Aeromonas hydrophila, Eterobacter aerogenes and Salmonella choleraesuis, lanes 4, 8 and 12 respectively. |
Transformed E. coli DH 1, initially expressed ampicilllin resistance at almost the same MIC of the original mother strain; the strainthat originally bear the plasmid, but gradually lose resistance resulting in ampicillin sensitive and plasmid free E. coli. Plasmid instability has been reported to be due to several reasons such as fragmentation, mutation or drop in copy number (Russell, 1998). It is also possible that a relationship between the nutritional conditions in which a host-plasmid system grown and the copy-number was inversely proportional to host cell growth rate (Caulcott et al., 1987). In other cases instability could be due to unacceptable metabolic load exerted by the plasmids on the host cell leading to high rates of plasmids loss (Diab, 1989). It could also be explained by being ecologically limited to their hosts (some sort of specificity) and thus could not adapt to exist for long in transformed E. coli (Hedges et al., 1985).
Due to limited number of endonucleases used in this study, restriction analysis could not proceed to beneficial points that originally supposed. But it is clear that one of the smallest plasmids, 7.4kbp, isolated from Salmonella choleraesuis showed the lowest ability, transformation coefficient of 0.0009, to transfer to the recipient strain E. coli DH 1. In contrast, the second largest plasmid, 23.1kbp, isolated from Enterobacter aerogenes showed high transferability (1.2). This may be in contrast with the idea of the smaller plasmid the higher the transferability of it to other cells (Bloom et al., 1995). The presence of certain genes on a plasmid may be responsible for such a behaviour (Chora, 1998).
The larger the plasmid the frequent the sequence at which endonucleases works on. This could be noticed when comparing the number of recognition sites and fragments of Pseudomonas aeruginosa and Enterobacter aerogenes plasmids (largest two plasmids isolated in this study) with others. For example digestion with EcoR 2 gave 2 fragments with both Pseudomonas aeruginosa and Enterobacter aerogenes while no fragments were noticed for other isolates with the same EcoR I.
The quite high number of fragments in Pseudomonas aeruginosa plasmid, 8 fragments, with BamH I may indicate the repeated sequence of G ATCC.
Detailed characterization of these plasmids is needed in future for more understanding about transferability, gene expression and stability.
REFERENCES
- Caulcott, C.A., A. Dunn, H.A. Robertson, N.S. Cooper, M.E. Brown and P.M. Rhodes, 1987. Investigation of the effect of growth environment on the stability of low-copy number plasmids in E. coli. J. Gen. Microbiol., 133: 1881-1889.
Direct Link - Goni-Urriza, M., M. Capdepuy, C. Arpin, N. Raymond, P. Caumette and C. Quentin, 2000. Impact of an urban effluent on antibiotic resistance of riverine Enterobacteriaceae and Aeromonas spp. Appl. Environ. Microbiol., 66: 125-132.
CrossRefPubMedDirect Link - Halling-Sorensen, B., S. Nors-Nielsen, P.F. Lanzky, F. Ingerslev, Holten, H.C. Luetzhoeft and S.E. Joergensen, 1998. Occurrence, fate and effects of pharmaceutical substances in the environment: A review. Chemosphere, 36: 357-393.
Direct Link - McKeon, D.M., J.P. Calabrese and G.J. Bissonnette, 1995. Antibiotic resistant gram-negative bacteria in rural groundwater supplies. Water Res., 29: 1902-1908.
CrossRefDirect Link - Niemi, M., M. Sibakov and S. Niemela, 1983. Antibiotic resistance among different species of faecal coliforms isolated from water samples. Applied Environ. Microbiol., 45: 79-83.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link