
Jwanny E. W.
Biochemistry Department, National Research Center, Tahrir St, Dokki, Cairo, Egypt
S. T. El-Sayed
Biochemistry Department, Faculty of Science, Ain Shams University, Egypt
A. M. Salem
Biochemistry Department, Faculty of Science, Ain Shams University, Egypt
A. N. Shehata
Biochemistry Department, National Research Center, Tahrir St, Dokki, Cairo, Egypt
Pakistan Journal of Biological Sciences
Year: 2001 | Volume: 4 | Issue: 3 | Page No.: 271-276







ABSTRACT
Chitinase, laminarinase A and laminarinase B were extracted and purified from Egyptian sugar beet (Beta vulgaris) leaves. Chitinase showed higher heat stability than laminarinase A and laminarinase B when heated at 50°C for 60 min. Chitinase activity was quite stable in water than in buffer, while laminarinase A and laminarinase B activities increased when incubated with 0.1M citrate-phosphate buffer at pH 6.0 to 7.0 for 30 min. The Km values of chitinase, laminarinase A and laminarinase B were 0.2, 0.27 and 0.074% at pH`s 4.5, 4.5 and 6.5 using colloidal chitin, laminarin A and laminarin B as substrates respectively. Chitinase enzyme was activated by 0.75 mM - mercaptoethanol by 1.6 times, while laminarinase A enzyme was activated by 1.0 mM of CuCl2, FeSO4 and EDTA with 2.35, 1.38 and 1.57 times respectively. Laminarinase B enzyme was activated by 1.0 mM Zn SO4 and K2SO4 with 1.3 and 1.18 times respectively. Chitinase, laminarinase A and laminarinase B enzymes have an endo-splitting type of activity. They were able to inhibit the growth and to lyse cell walls of Aspergillus oryza and A. flavus either alone or in combination through the degradation of chitin and laminarin.
PDF Abstract XML References Citation
How to cite this article
Jwanny E. W., S. T. El-Sayed, A. M. Salem and A. N. Shehata, 2001. Characterization and Antifungal Evaluation of Chitinase and
Laminarinases from Sugar Beet Leaves. Pakistan Journal of Biological Sciences, 4: 271-276.
DOI: 10.3923/pjbs.2001.271.276
URL: https://scialert.net/abstract/?doi=pjbs.2001.271.276
DOI: 10.3923/pjbs.2001.271.276
URL: https://scialert.net/abstract/?doi=pjbs.2001.271.276
INTRODUCTION
Nowadays production, purification and characterization of plant enzymes with biological importance have been reported for using in many fields. Chitinase (EC.3.2.1.14) and laminarinase (endo-β-1,3-glucanase, EC.3.2.1.6) are present in wide variety of plants including vegetative parts and seeds (Deloire et al., 1997), invertebrates and vertebrates (Han et al., 1997) and in microorganisms (Porfireva et al., 1997).
In the past decade, chitinases and laminarinases have been subjected to much attention since their obvious substrates chitin and β-1,3 glucan, are the major cell wall components of many plant pathogenic fungi (Bartnicki-Garcia, 1968). Most studies showed that chitinases and laminarinases of plants are endotype and contain lysozyme activity (Mauch et al., 1988). They have great ability to inhibit the growth of many fungi, pathogenic or non pathogenic, comparing with those isolated from microorganisms (Roberts and Selitrennikoff, 1988). The inhibition being caused by iysis of growing hyphal tips through degradation of the cell wall (Ariolo et al., 1992; Benhamou et al., 1993a). More experiments confirmed that these enzymes also have an antifungal potential in vivo (Benhamou et al., 1993b). This investigation has been directed towards the characterization of the purified chitinase and laminarinases from sugar beet leaves. It also describes the specificity, action pattern and the antifungal activity of the prepared enzymes on some Aspergillus species.
MATERIALS AND METHODS
Enzymes preparation and assay: Extraction, purification and assays of chitinase, laminarinase A and laminarinase B from sugar beet leaves were done according to the methods described in the previous papers (Salem et al., 1999; EI-Sayed et al., 2000). The prepared pure enzymes were used for studying their physicochemical properties.
Stability of the enzyme at different pH: Small aliquots of the purified enzymes (chitinase, laminarinase A and laminarinase B) were incubated with 0.1 M acetate (pH 2.5 to 5.0) and citrate-phosphate (pH 5.5 to 7.0) buffer at 9°C for 30 min. Then, the remaining relative activities were assayed.
Heat stability: Small aliquots of the purified enzymes were preheated at different temperatures, 30, 40, 50 and 60°C for varying time intervals (15,30,45 and 60 min). The remaining enzymatic activities were then assayed.
Effect of some activators and inhibitors on enzymatic activities: This effect was done by preincubation of each of K2SO4, CuCl2, CaCl2, MgCl2, CoCl2, BaCl2, CuSO4, ZnSO4, Hg(NO3)2, FeSO4 and AgNO3 with the purified enzyme at concentration of 1.0 mM for 30 min at 9°C before determination of enzyme activity. EDTA, cysteine and amercaptoethanol were also preincubated with the purified enzyme at concentration of 1.0 mM, While pchloromercuribenzoate (PCMB) at concentration of 2×107 mM was also incubated with the purified enzyme at 9°C for 30 min before determination of the enzymes activities. The effect of different concentrations (0.5 to 2.0) of amercaptoethanol on chitinase, of CuCl2, FeSO4 and EDTA on laminarinase A and of K2SO4 and ZnSO4 on laminarinase B also were estimated.
Michael's constant: Apparent Michael's constants were determined according to Lineweaver and Burk (1934) using chitin, laminarin A and laminarin B as substrates.
Substrate specificity: Solutions of some 13-D-glucan of different linkage types were prepared at concentrations of 0.25, 0.5 and 1.0%. Soluble chitosan was prepared from flaked chitosan as described by Davis and Eveleigh (1984). The soluble chitosan was dissolved in distilled H2O to form 1.0% solution. These substrates were incubated separately with chitinase, laminarinase A and laminarinase B for activities measurements.
Analysis of enzymes reaction products: The end products of incubating chitinase, laminarinase A and laminarinase B with chitin, laminarin A and laminarin B respectively were examined by paper chromatography according to the method of Wilson (1959). Chromatograms were developed by descending chromatography. The solvent system pyridin:n-butanol:water (9:15:6) was used. Reducing sugars on the chromatograms were detected with aniline phthalate reagent.
Antifungal activity of chitinase, laminarinase A and laminarinase B: A number of standard Aspergillus species namely, Aspergillus flavus NRRL 627, 3676 and 2210 and Aspergillus oryza NRRL 3502, were used throughout this study. Cultures were kindly provided from the collection of U.S. Department of Agriculture Research Service, California. The biological effects of the purified enzymes were determined by measuring their inhibitory effect on the growth of fungi and their lysing effect on the cell wall of fungi.
Inhibition of fungi growth: Glucose peptone medium composed of 20 g glucose, 10 g peptone and 20 g agar in 1.0 liter distilled H2O was used. Antifungal activity was determined under sterile conditions by detecting zone of inhibition (Roberts and Selitrennikoff, 1986) and by evaluating the decreasing of dry weight of the fungus in the liquid medium at the end of the culture (Yoshida et al., 1987).
Cell wall lytic activity: Aspergillus species cell walls were prepared according to the method of Kisser et al. (1980). The mycelium of one day old shaken culture on glucose-peptone medium was used.
Method A: In three test tubes, enzyme preparations (chitinase, laminarinase A and laminarinase B) were added separately to the cell wall suspension in acetate (0.1 M, pH 4.5) and in citrate-phosphate buffer (0.1 M, pH 4.5, 6.5) respectively. The reaction mixtures were incubated at 40°C for 3 hr. Assay for reducing sugars were determined according to the methods described by Smogyi (1952) and Nelson (1944).
International milliunits (ImU) of the lytic activity was defined as the quantity of enzyme, which release 1 nmole of reducing sugar from fungi cell wall per min. at 40°C.
Method B: Cell wall suspension giving an optical density of about 0.3 to 0.7 at 660 nm against distilled H2O was used. Enzyme preparations were added to the cell wall suspension either in acetate (0.1 M, pH 4.5) for chitinase or in citrate-phosphate buffers (0.1 M, pH 4.5 and 6.5) for laminarinase A and laminarinase B respectively. The reaction mixtures were incubated at 40°C for 2 hr. The decrease in turbidity of the cell wall suspension was measured at 660 nm. One unit of lytic activity is defined as the quantity of enzyme which reduces the initial optical density by 10% per hr. (Tsujisaka et al., 1973).
RESULTS
This study is a part of a comprehensive effort which aims to prepare pure enzymes having the ability to inhibit fungal growth and also degrade fungal cell walls. Previously, some plants extracts were obtained through extensive screening study to find chitinase, laminarinase A and laminarinase B activities (Salem et al., 1999; EI-Sayed et al., 2000). Of all the active plants extracts, Egyptian sugar beet leaves were found to be the most considerable producer of these enzymes in the screened plants.
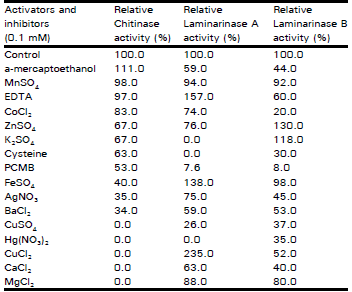
| Table 1: | Effect of some activators and inhibitors on enzymes activities |
 | |
Physicochemical properties of the prepared enzymes: The data presented in Fig. 1 shows the effect of different pH's on enzymatic stability. In absence of substrates, chitinase activity deceased by about 12 to 20.7%, when the pure chitinase enzyme was preincubated with 0.1M, acetate (pH 3.5 to 5.5) buffer for 30 min. at 9°C. It is more stable in water than in buffer. Laminarinase A and laminarinase B activities increased by 5 to 22 and 1.0 to 30% respectively when the pure enzymes were preincubated with 0.1 M acetate and citrate- phosphate buffer (pH 2.5 to 7.0) at 9°C for 30 min.
The results presented in Fig. 2 indicate the effect of different temperatures on enzymatic activity. In absence of substrates, chitinase enzyme showed marked thermostability at 60°C for 30 min with 22% loss of its activity. It looses 80% of its activity when heated at 60°C for 30min. Laminarinase A lost 28 and 33% of its activity when heated at 30 and 40°C for 15min. while results showed that laminarinase B lost 17 to 42% of its activity when heated at 30 and 40°C respectively for 15 min. Complete inactivation of activities of laminarinase A and laminarinase B were observed when heated at 50 and 60°C for 60 min.
The Arhenius activation energy for chitinase and laminarinase A and laminarinase B were 14.6, 13.6 and 9.8 Kcal. respectively. Similarly, the Arhenius activation energy for the enzyme catalyzed hydrolysis of laminarin was found to be 9.7 Kcal (Huotari et al., 1968).
The effects of various cations on enzymes activities were tested (Table 1). Complete inhibition of chitinase enzyme was noticed with some heavy metals CuSO4, Hg (NO3)2, CuCl2, CaCl2 and MgCl2, while a-mercaptoethanol is considered as activator as it increased the activity by 1.11 fold. When the effect of different concentrations (0.5 to 2.0 mM) of a-mercaptoethanol on chitinase was estimated, 0.75 mM concentration was found to stimulate the chitinase activity optimally by 1.6 fold.
Laminarinase A enzyme activity was increased by CuCl2, EDTA and FeSO4 (2.35, 1.57 and 1.38 times respectively), while laminarinase B activity was increased by ZnSO4 and K2SO4 (1.3 and 1.18 times respectively).
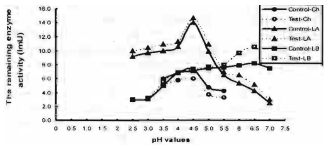 | |
| Fig. 1: | Effect of different pH values on chitinase (Ch.), Laminarinase A (LA) and Laminarinase B (LB) stability. Test: enzyme activity after preincubation with buffer for 30 min, control: enzyme activity withour preincubation with buffer |
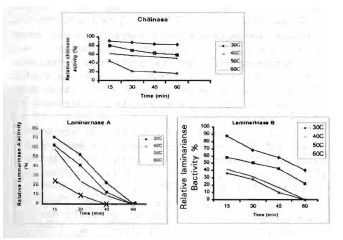 | |
| Fig. 2: | The effect of different temperatures on chitinase, laminaarinase A and laminariiase B stability at different times |
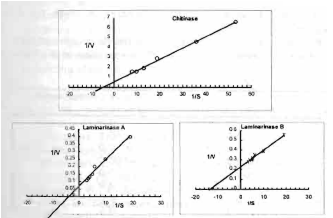 | |
| Fig. 3: | Lineweavear-Burk plot for hydrolysis of chitin, laminarin A and laminarin B by chitinase, laminarinase A and laminarinase B respectively |
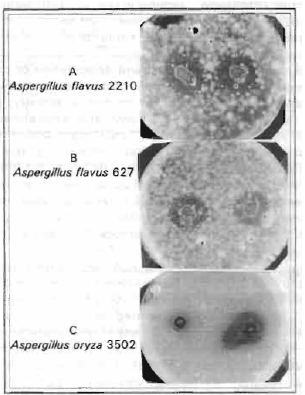 | |
| Fig. 4: | Antifungal activities in vitro of the purified three enzymes, chitinase and laminarinases (A and B) against Aspergillus flavus 2210 (A), A. Flavus 627 (B) and A. oryza 3502 (C) |
The effect of different concentration (0.5 to 2.0 mM )of CuCl2, EDTA and FeSO4 on laminarinase A and ZnSO4 and K2SO4 on laminarinase B also were estimated. The best metals concentration for laminarinase A and laminarinase B activation was found to be 1.0 mM.
Absorption spectrum of the three enzymes in 0.01 M phosphate buffer, pH 7.0 was measured at 220 to 350 nm, The enzymes showed maximum absorbancy at wave length 275 nm. The ratio of absorbency at 280 nm to 260 nm were 1.05, 1.02 and 1.08 for chitinase, laminarinase A and laminarinase B respectively.
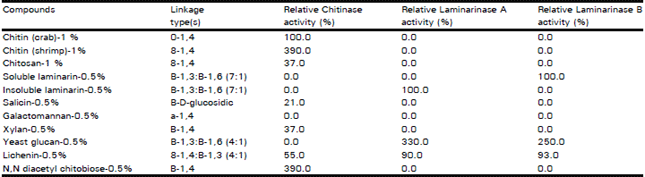
The purified chitinase enzyme was found to have hydrolytic effect on β-1, 4 linkage (chitin from crab and shrimp, chitosan, salicin, xylan, lichenin) but lacking hydrolytic activity towards β-1, 3, β-1, 6 and α-1, 4 linkage (Table 2). Chitosan was degraded to 37% that of chitin, since chitosan is only partially deacetylated. The purified chitinase was also found to have hydrolytic effect on N, N' diacetylchitobiose. On the other hand, the purified laminarinase A and laminarinase B enzymes showed strong hydrolytic action on β-1, 3 linkage (laminarin A and laminarin B, yeast glucan and lichenin) and failed to hydrolyze a or β-1, 4 linkages.
The Michael's constant (Km) was 0.2% for chitinase, 0.27% for laminarinase A and 0.074% for laminarinase B using chitin, laminarin A and laminarin B as substrates respectively (Fig. 3). The Km of laminarinase B is relatively low which indicates the presence of high affinity between the enzyme and the substrate. Laminarinase from wheat grain has higher Kn, 0.35% (Luchsinger et al., 1963). The Vmax were 2.27, 15.87 and 9.5 U for chitinase, laminarinase A and laminarinase B respectively.
Chromatographic analysis of the products released upon incubation of chitin or laminarin A or laminarin B with the purified chitinase, laminarinase A and larninarinase B respectively revealed the presence of several oligosaccharides and small amounts of the monomers.
Inhibition of fungal growth and degradation of the isolated fungal cell walls by chitipase, larrinerinase A and laminarinase: To test for antifungal activity, aliquots of the prepared chitinase (1.17 mU) and Ifirninerinase A (3.17 ImU) and larninarinase B (2.11 mU) were added to a well in four agar plates containing germinating four different fungal spores. After 24 hr, distinct inhibition zones developed around the well (Fig. 4). The growth of the two strains Aspergillus flavus 2210 and 627 was strongly and moderately inhibited respectively, while Aspergillus oryza 3502 was slightly inhibited but Aspergillus flavus 3676 was not affected.
When 0.67 ImU chitinase and/or 4.2 ImU laminarinase A and/or 2.33 ImU laminarinase B were incubated with spore of Aspergillus flavus 2210 in fungal liquid medium at 37°, the dry weight declined by (22 to 75%) in 24 hr (Table 3). The decrease percent of dry weights by chitinase or by laminarinase A or by laminarinese B enzymes represents antifungal activities of the enzymes.
The results of fungal cell wall lysis by chitinase, laminarinase A and laminarinase B either alone or in combination are shown also in Table 4. The extent of wall hydrolysis was measured by assaying released reducing sugars and by determining the increase of turbidity. Results demonstrated that purified plant chitinase, laminarinase A and laminarinase B are capable of liberating reducing sugars from the fungal cell walls and exhibited lysoiyme activity.
DISCUSSION
Recently, there has beep much interest in production of antifungal enzymes from natural sources. The prepared pure chitinase from sugar beet leaves is more stable in water than in acetate buffer, while the prepared pure laminarinase A and laminarinase B showed high stability when kept in the citrate-phosphate buffer.
Chitinase is more stable than laminarinase A and laminarinase B when heated up to 50°C for 60 min. Similarly, chitinase from carrot roots was relatively stable when heated at temperatures below 60°C (Zhang et al., 1996), while Laminarinase from soybean and Nicotiana glutinosa leaves, were inactivated at 65°C (Moore and Stone, 1972a; Keen and Yoshikawa, 1983).
The complete inhibition of chitinase with some heavy metals CuSQ4, Hg (NO3)2, CuCl2, CaCl2 and MgCl2 and laminarinase A with K2SO4 and Hg (NO3)2 could be related to the creation of the cation to chitin or laminarin A subkratesin turn cleavage or hydrolysis of substrates by the enzymes could be inhibited. These results agree with those of stomach eel and cabbage stem chitinases and of Nicotiana glutin larninarinase (Moore and Stone, 1972a; Kono et al., 1990; Chang et al., 1996). The activation of chitjnase b y gt-mercaptoethanol, of laminarinase A by CuCl2, EDTA and FeSO4 and of laminarinase B by ZnSO4 and K2SO4 may be due to the facility of the formation of enzyme substrate complex.
Spectrum patterns of the prepared enzymes from sugar beet leaves are typical protein profile and quite similar to amylases, enzymes from beet root and turnip leaves (El-Sayed, 1994).
The purified chitinase enzyme was found to have hydrolytic effect on β-1, 4 linkage and on the other hand the purified laminarinase A and laminarinase B enzymes showed strong hydrolytic action on β-1, 3 linkage. Science chitosan is partial deagetylated, the small degradation of chitosan (37% that of chitin) by chitinase may be due to stretches of polysaccharide chains that conserve the acetyl groups (Molano et al., 1979). The presence of chitobiase activity with the prepared pure chitinase was not surprising, since chitinase and chitobiase activities oftenly are purified together and separation of the two enzymes was difficult. This result was coinciding with those of Wadsworth and Zikakis (1984).
The purified three enzymes behave as endo-type and free of exo activity. These results were in agreement with endochitinase from bean leaves and carrot roots (Boller et al., 1983; Zhang et al., 1996). Endolaminarinase also could be isolated from Nicotiana glutinosa leaves (Moore and Stone, 1972b) and from grape vine seeds (Clarke and Stone, 1962). However, exochitinase activity has been reported from number of plants (Boller et al., 1983). An endohydrolytic activity would appear to be more suitable for the postulated defense function than an exohydrolytic activity, since the former could destroy the polymer fabric of pathogen cell wall more effectively.
The data presented herein demonstrates that the purified chitinase, laminarinase A and laminarinase B have strong inhibitory effect on growth of Aspergillus oryza and A. flavus.
They also are involved in lysis and in the dissolution of their cell walls, while in the case of combination of the three enzymes, greater effect was observed due to synergistic effect of the three enzymes.
| Table 2: | Substrate specificity of sugar beet leaves enzymes |
 | |
| Table 3: | Antifungal activities of Chitinase, Laminarinase A and Laminarinase B on Aspergillus flavus 2210 in liquid medium |
 | |
| Table 4: | Degradation of Aspergillus cell walls |
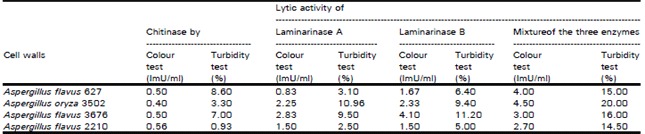 | |
| N.B. 1- Chitinase, laminarinase A and laminarinase B contain 0.67, 4.2 and 2.32 ImU/ml 2- The mixture of the three enzymes contains 0.8, 5.0 and 2.8 ImU/ml | |
This is related to the dissolution of the two major cell wall polymers (chitin and β-1, 3-glucan). These results are in agreement with those of Mauch et al. (1988) and Huynh et al. (1996). Two independent bioassays demonstrated an antifungal effect of sugar beet chitinase on Cercospoa beticola (Nielsen et al., 1993). However, the chitinase enzyme from carrot roots did not possess lysozyme activity but showed antifungal activity when tested against Trichoderma sp. (Zhang et al., 1996).
Several studies on the isolation and preparation of antifungal enzymes chitinase, laminarinases from microorganisms and plants have been investigated (Chang et al., 1996; Porfireva et al., 1997), but it was found that the effect of antifungal enzymes from plants is more potent in inhibition of growth of fungi than that from microorganisms (Park et al., 1992). These results may be due to that chitinase in plants are endochitinase which randomly hydrolyze internal β-1, 4-linkages of chitin releasing oligosaccharides of N-acetylglucsamine. Therefore, preparations of the purified chitinase, laminarinase A and laminarinase B from plants were carried out in this investigation. These enzymes may play a role in protecting plants against attack from pathogens fungi.
REFERENCES
- Ariolo, M., A. Ludwig, T. Boller and P. Bonfante, 1992. Inhibition of fungal growth by plant chitinases and β-1,3-glucanases. Protoplasma, 171: 34-43.
CrossRefDirect Link - Bartnicki-Garcia, S., 1968. Cell wall chemistry, morphogenesis and taxonomy of fungi. Annu. Rev. Microbiol., 22: 87-108.
CrossRefPubMedDirect Link - Benhamou, N., K. Broglie and I. Chet, 1993. Antifungal effect of bean endochitinase on Rhizoctonia solani: Ultrastructural changes and cytochemical aspects of chitin breakdown. Can. J. Microbiol., 39: 318-328.
PubMedDirect Link - Boller, T., A. Gehri, F. Mauch and U. Vogeli, 1983. Chitinase in bean leaves: Induction by ethylene, purification, properties and possible function. Planta, 157: 22-31.
CrossRefPubMedDirect Link - Chang, C.T., Y.L. Hsueh and H.Y. Sung, 1996. Purification and properties of chitinase from cabbage stems with roots. Biochem. Mol. Biol. Int., 40: 417-425.
PubMedDirect Link - Clarke, A.E. and B.A. Stone, 1962. β-1,3-glucan hydrolases from the grape vine (Vitis vinifera) and other plants. Phytochemistry, 1: 175-188.
CrossRefDirect Link - Deloire, A., E. Kraeva, M.C. Maura, E. Bonnet and J. Bierne, 1997. Immunodetection of chitinase-like and (β-1,3-glucanase-like proteins secreted in vitro by embryogenic and non-embryogenic cells of grapevines. Vitis, 36: 51-52.
Direct Link - El-Sayed, S.T., A.M. Salem, A.N. Shehata and E.W. Jwanny, 2000. Chitinase from leaves of Beta vulgaris and other higher plants. Pak. J. Biol. Sci., 3: 250-256.
CrossRefDirect Link - Han, B.K., W.J. Lee and D.H. Jo, 1997. Survey on the chitinolytic activity from some animals for the industrial utilization. Applied Biol. Chem., 40: 95-100.
Direct Link - Huotari, F.I., T.E. Nelson, F. Smith and S. Kirkwood, 1968. Purification of an Exo-β-D-(1 → 3)-glucanase from Basidiomycete species QM 806. J. Biol. Chem., 243: 952-956.
Direct Link - Huynh, Q.K., J.R. Borgmeyer, C.E. Smith, L.D. Bell and D.M. Shah, 1996. Isolation and characterization of a 30 kDa protein with antifungal activity from leaves of Engelmannia pinnatifida. Biochem. J., 316: 723-727.
CrossRefDirect Link - Keen, N.T. and M. Yoshikawa, 1983. β-1,3-endoglucanase from soybean releases elicitor-active carbohydrates from fungus cell walls. Plant Physiol., 71: 460-465.
CrossRefDirect Link - Kisser, M., C.P. Kubicek and M. Rohr, 1980. Influence of manganese on morphology and cell wall composition of Aspergillus niger during citric acid fermentation. Arch. Microliol., 128: 26-33.
CrossRefDirect Link - Kono, M., T. Matsui, C. Shimizu and D. Koga, 1990. Purification and some properties of chitinase from the stomach of Japanese Eel, Anguilla japonica. Agric. Biol. Chem., 54: 973-978.
CrossRefDirect Link - Lineweaver, H. and D. Burk, 1934. The determination of enzyme dissociation constants. J. Am. Chem. Soc., 56: 658-666.
CrossRefDirect Link - Mauch, F., B. Mauch-Mani and T. Boller, 1988. Antifungal hydrolases in pea tissue: II. Inhibition of fungal growth by combinations of chitinase and β-1,3-glucanase. Plant Physiol., 88: 936-942.
CrossRefPubMedDirect Link - Molano, J., I. Polacheck, A. Duran and E. Cabib, 1979. An endochitinase from wheat germ. Activity on nascent and preformed chitin. J. Biol. Chem., 254: 4901-4907.
PubMedDirect Link - Moore, A.E. and B.A. Stone, 1972. A β-1,3-glucan hydrolase from Nicotiana glutinosa: I. Extraction, purification and physical properties. Biochim. Biophys. Acta (BBA)-Enzymol., 258: 238-247.
CrossRefDirect Link - Moore, A.E. and B.A. Stone, 1972. A β-1,3-glucan hydrolase from Nicotiana glutinosa II. Specificity, action pattern and inhibitor studies. Biochim. Biophys. Acta (BBA)-Enzymol., 258: 248-264.
CrossRefPubMedDirect Link - Nielsen, K.K., J.D. Mikkelsen, K.M. Kragh and K. Bojsen, 1993. An acidic class III chitinase in sugar beet: induction by Cercospora beticola, characterization and expression in transgenic tobacco plants. Mol. Plant Microbe Interact, 6: 495-506.
PubMedDirect Link - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
CrossRefDirect Link - Porfireva, O., D. Yusupova, N. Zotkina, R. Sokolova and L. Gabdrakhmanova, 1997. Chitinolytic complex of Serratia marcescens and Peculiarities of its biosynthesis. Mikrobiologiya, 66: 347-353.
Direct Link - Roberts, W.K. and C.P. Selitrennikoff, 1986. Isolation and partial characterization of two antifungal proteins from barley. Biochim. Biophys. Acta (BBA)-Gen. Subjects, 880: 161-170.
CrossRefDirect Link - Roberts, W.K. and C.P. Selitrennikoff, 1988. Plant and bacterial chitinases differ in antifungal activity. J. Genet. Micobiol., 134: 169-176.
CrossRefDirect Link - Tsujisaka, Y., Y. Tominaga and M. Iwai, 1973. Taxonomic characters and culture conditions of a bacterium which produces a lytic enzyme on rhizopus cell wall. Agric. Biol. Chem., 37: 2517-2525.
Direct Link - Wadsworth, S.A. and J.P. Zikakis, 1984. Chitinase from soybean seeds: Purification and some properties of the enzyme system. J. Agric. Food Chem., 32: 1284-1288.
CrossRefDirect Link - Wilson, C.M., 1959. Quantitative determination of sugars on paper chromatograms. Anal. Chem., 31: 1199-1201.
CrossRefDirect Link - Yoshida, S., S. Kasuga, N. Hayashi, T. Ushiroguchi, H. Matsuura and S. Nakagawa, 1987. Antifungal activity of ajoene derived from garlic. Applied Environ. Microbiol., 53: 615-617.
PubMedDirect Link - Zhang, Y.Y., N.H. Haunerland and Z.K. Punja, 1996. Chitinase profiles in mature carrot (Daucus carota) roots and purification and characterization of a novel isoform. Physiol. Planta, 96: 130-138.
CrossRefDirect Link