
D. C. Ang
Department of Biotechnology, Faculty of Food Science and Biotechnology, Universiti Putra Malaysia,
43400 Serdang, Selangor, Malaysia.
S. Abd-Aziz
Department of Biotechnology, Faculty of Food Science and Biotechnology, Universiti Putra Malaysia,
43400 Serdang, Selangor, Malaysia.
H. M. Yusof
Department of Biotechnology, Faculty of Food Science and Biotechnology, Universiti Putra Malaysia,
43400 Serdang, Selangor, Malaysia.
M.I. A. Karim
Department of Biotechnology, Faculty of Food Science and Biotechnology, Universiti Putra Malaysia,
43400 Serdang, Selangor, Malaysia.
A. Ariff
Department of Biotechnology, Faculty of Food Science and Biotechnology, Universiti Putra Malaysia,
43400 Serdang, Selangor, Malaysia.
K. Uchiyama
Department of Biotechnology, Graduate School of Engineering,
Osaka University, 2-1 Yamadaoka, Suit a, Osaka 565, Japan
S. Shioya
Department of Biotechnology, Graduate School of Engineering,
Osaka University, 2-1 Yamadaoka, Suit a, Osaka 565, Japan
Pakistan Journal of Biological Sciences
Year: 2001 | Volume: 4 | Issue: 3 | Page No.: 266-270







ABSTRACT
Rice α-amylase and glucoamylase produced by S. cerevisiae YKU 107 and YKU 131, respectively were partial purified using anion exchange chromatography and characterised. The optimum active conditions for -amylase was at pH 5.0 and temperature of 50°C. It was capable in hydrolysing more than 85% of potato, rice, cassava and sago starch, however, it could only hydrolysed 64% of corn starch. Glucoamylase was optimally active at pH 5.5 and temperature of 55°C. Glucoamylase could hydrolyse more than 70% of cassava, sago and potato starch, it also hydrolysed more than 65% of corn and rice starch. Both of the enzymes were not severely affected by the changes of ionic strength of acetate buffer at molarity lower than 0.20 M.
PDF Abstract XML References Citation
How to cite this article
D. C. Ang, S. Abd-Aziz, H. M. Yusof, M.I. A. Karim, A. Ariff, K. Uchiyama and S. Shioya, 2001. Partial Purification and Characterisation of Amylolytic Enzymes Obtained from Direct Fermentation
of Sago Starch to Ethanol by Recombinant Yeast. Pakistan Journal of Biological Sciences, 4: 266-270.
DOI: 10.3923/pjbs.2001.266.270
URL: https://scialert.net/abstract/?doi=pjbs.2001.266.270
DOI: 10.3923/pjbs.2001.266.270
URL: https://scialert.net/abstract/?doi=pjbs.2001.266.270
INTRODUCTION
The conversion of biomass to ethanol has been paid significant attention as a alternative sources of fuels since the oil crises of 1970s. The renewable resources that can be used in ethanol fermentation are sugar bearing materials, starches and cellulose. Traditionally, ethanol fermentation is relied on sugar-rich substrate, mainly sugarcane, because of their carbohydrate is in fermentable sugar form. However, sugarcane is expensive and not continuously available because it is a seasonal crop that required high-quality agricultural land for its growth (De Moraes et al., 1995). Thus, there are great economic advantages to extent the substrate range of the ethanol-fermenting microorganism so that the ethanol may be produced from, mainly starchy crops (Marzola and Bartholomew, 1979; Coombs, 1984; Hacking, 1987; Rosillo-Calle et al., 1992). However, the ethanol-fermenting microorganisms, such as S. cerevisiae and Zymomonas mobilis are lack of amylolytic enzymes and unable to directly convert the starch into ethanol. Therefore, it is necessary to breed an ethanol-fermenting microorganism by a genetic manipulation, which can directly ferment starch materials into ethanol.
The yeast Saccharomyces cerevisiae, the main microorganism used for alcoholic fermentation, is widely used as a host cell for foreign gene products due to the abundance of information that available following by the early development of recombinant DNA techniques for S. cerevisiae (Hinnen et al., 1978; Beggs, 1978). On the other hand, yeast is an eukaryotic cell that has an ability to produce mature foreign protein from plants or animals. Lately, studies had also focused on genetically modification of S. cerevisiae for α-amylase and glucoamylase expression (Steyn and Pretorious, 1990; Uchiyama et al., 1995; Lee et al., 1998). The main purpose of genetic modification of S. cerevisiae was to realise the one-step bioconversion of starch to ethanol, in order to minimise the production costs of fermentation ethanol.
In this study, attempts were made to characterise the amylolytic enzymes produced by the recombinant yeasts, S. cerevisiae YKU 107 (expressing α-amylase) and S. cerevisiae YKU 131 (expressing glucoamylase) as a preliminary study before utilising the yeast to directly produce ethanol from starch.
Materials and Methods
Strains and media: In this study, the recombinant yeasts, YKU 107 and YKU 131 were kindly provided by Department of Biotechnology, Graduate School of Engineering, Osaka University, Japan. The temperature-sensitive mutants of Saccharomyces cerevisiae for the PHO regulatory system, YKU 76 (MAT a ura3-52 trp1 lue2 his3 pho 80-69ts) was use as host strain. S.cerevisiae YKU 107 is a transformant of YKU 76 with rice α-amylase expression vector, p739 (Uchiyama et al., 1995). S. cerevisiae YKU 131 is a transformant of YKU 76 with glucoamylase expression vector, pKU 122 (Lee et al., 1998). Saccharomyces cerevisiae strain YKU 107 and YKU 131 was maintained on a selective agar, respectively. Inoculum was prepared by germinating the cells from the selection agar in a 500-mL flask containing 100 mL of selection medium, incubated at 30°C for 16 hrs. For enzymes purification, 15 mL of the inoculum was added to 150 mL of media containing 2% sago starch, 2% peptone and 1% yeast extract in a 500-mL flask. The organisms were culture at 30°C for 2 days on a rotary shaker at 180 rpm.
Partial Purification of α-Amylase from Strain YKU 107
Step 1. Acetone precipitation: The culture was centrifuged at 4000 rpm for 15 minutes at 4°C, acetone that previously chilled to -20°C was added dropwise at 4°C to 100 mL of supernatant fluid with continuous stirring to the final proportion of 1:1. The solution was left at 4°C for 24 hours. The resultant precipitate was collected by centrifugation (10,000g for 15 minutes) at 4°C.
Step 2. DEAE-Cellulose column chromatography: The acetone-precipitated protein was dissolved in 0.05 M acetate buffer, pH 5.3 and concentrated by ultrafiltration membrane before applied to a DEAE-Cellulose column (2.5x40cm) equilibrated previously with 0.05 M acetate buffer, pH 5.3. Fractionation was carried out using the same buffer containing 0.2 M of NaCl at flow rate 30mLh-1. The protein fractions of 5 mL per tube were collected and the fractions given reading at OD280 were collected for α-amylase activity assay. The fractions containing α-amylase activity were combined and dialysed against with 1 L distilled water for 5 hours and concentrated by freeze-drying. The concentrated enzyme solution was used for enzyme characterisation.
Partial Purification of Glucoamylase from Strain YKU 131
Step 1. Ammonium sulphate precipitation: The culture was centrifuged at 4000 rpm for 15 minutes at 4°C, 55.1 g of powdered ammonium sulphate was added slowly to 100 mL of supernatant fluid with constant stirring at 4°C to the final ammonium saturation of 75%. The protein precipitation was collected by centrifugation (10,000g for 15 minutes) at 4°C.
Step 2. DEAE-Cellulose column chromatography: The protein collected was re-dissolved in 0.05 M of sodium phosphate buffer (pH 7.3). The solution was then applied to a DEAE-cellulose column (2.5 x 40 cm) equilibrated with the same buffer. The protein was eluted with the same buffer containing 0.3 M NaCl at a flow rate of 30 mLh-1. The protein fractions of 5 mL per tube were collected and the fractions given reading at OD280 were taken for glucoamylase activity assay. The fractions containing glucoamylase activity were pooled and dialysed against 1 L distilled water for 5 hours and concentrated by freeze-drying. The concentrated enzyme solution was used for enzyme characterisation.
Analytical Procedure
α-Amylase activity: The substrate for the α-amylase reaction (Thomsen, 1983) was prepared by adding 0.2 g soluble starch to 100 mL of boiling 50 mM sodium acetate buffer (pH 5.9) and the solution was cooled to 40°C. The iodine reagent was made by diluting 1 mL stock solution (0.5% I2 in 5% KI) in 500 mL deionised water containing 5 ml of 5 M HCl. The assay consisted of incubating 200 μL enzyme solution with 1 mL of starch solution at 40°C for 10 minutes. To stop the reaction, an aliquot (200 μL) was added to 5 mL iodine solution and the degradation of starch was measured at OD620 against an appropriate blank. One unit of α-amylase activity was defined as the quantity of enzyme required to hydrolyse 0.1 mg starch in 10 minutes, at 40°C, when 2.0mg starch was present at the start of the reaction.
Glucoamylase activity (Bon, 1991) was determined using 2 mL of culture supernatant with 18 mL of 60 mM maltose as substrate (pH 4.4 using acetate buffer) incubated at 40°C. Samples (0.5 mL) were taken from the reaction mixture at certain intervals time (10, 20 and 30 minutes) and the reaction was stopped by adding 0.5 mL of 0.05 M Tris/HCl buffer (pH 9) and then kept in an ice bath until the measurement of glucose concentration. The glucose produced from the reaction was determined using a glucose (Trinder) 100 (Sigma Diagnostics, USA). Glucoamylase activity was expressed as μmole of glucose liberated per minute per mL of broth supernatant (μmole.min-1.mL-1) or unit per mL (U.mL-1).
Protein concentration was estimated by the modified Lowry method described by Hartree (1972) using bovine serum albumin (BSA) as a standard. The amount of protein in each sample was obtained using the BSA standard curves.
Starch concentration was determined colorimetrically (Smith and Reo, 1948) on the basis of iodine starch complex colour. An aliquot of sample was prepared by mixing 100 μL of supernatant from a culture broth with 60 μL of 0.1 M phosphate buffer (pH 7.2) 200 μL of 5 M NaCl, 40 μL of 1 M HCl and 50 μL of iodine reagent. 10 mL of distilled water was added and the absorbance of the colour was read at OD625 on a spectrophotometer.
Results
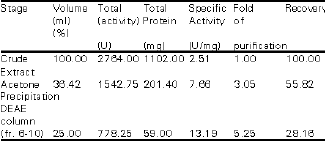
Partial Purification of α-Amylase from S.cerevisiae YKU 107 The typical purification profile of the α-amylase is shown in Table 1. The acetone precipitated crude extract when passed through the DEAE-cellulose column showed a major protein peak (fraction 6-10) that corresponded to the α-amylase activity. The elution profile of the extracellular α-amylase is shown in Figure 1. The enzyme was partial purified 5.25-fold with a yield of 28.16% to a specific activity of 13.19 units per gram protein.
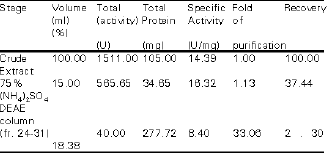
Partial Purification of Glucoamylase of S. cerevisiae YKU 131 Table 2 shows the typical purification profile of the glucoamylase. The ammonium sulphate precipitated crude extract when passed through the DEAE-cellulose column showed a major protein peak (fraction 24 - 31) that corresponded to the glucoamylase activity. The elution profile of the glucoamylase is shown in Figure 2. The enzyme was purified 2.30-fold with a recovery of 18.38% to a specific saccharifying activity of 33.06 U/gram protein.
Properties of α-amylase and glucoamylase
pH Optimum and Stability: Effect of pH on the activity and stability of S.cerevisiae YKU 107 α-amylase as well as S.cerevisiae YKU 131 glucoamylase are shown in Figure 3 and Figure 4 respectively. The effect of pH on the activity of the both enzymes was determined under standard assay conditions except for the changes of pH, using 0.05 M acetate buffer (pH 3.0 – 5.5) and 0.05 M phosphate buffer (pH 6.0 – 9.0). The effect of pH on the stability of α-amylase and glucoamylase was obtained by incubating the enzyme solutions at 35°C with various pH values for 24 hours. The buffers used were the same to those for enzyme activity determination. After each incubation time, the residual activity was estimated by standard assay procedure. The optimum pH of α-amylase was 5.0 and the enzyme was stable at pH between 4.5 and 6.0 with residual activity around 90% and above. The optimum pH of glucoamylase activity was pH 5.5. The enzyme was stable at pH between 4.0 and 6.0 with residual activity higher than 90%.
Effect of ionic strength: The effect of ionic strength on the α-amylase and glucoamylase activities was determined under standard assay conditions except for the changes of ionic strength of acetate buffer (0.02-0.40 M). Figure 5 shows that both of the enzymes optimally active at ionic strength 0.05M and did not severely affect by the ionic strength at the range of 0.05-0.20 M, with relative activity higher than 90%.
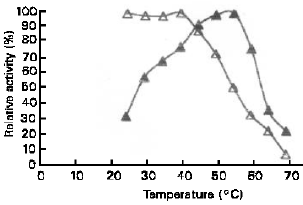
Effect of temperature: The effect of temperature on the activity of the α-amylase and glucoamylase was determined under standard assay condition except for the changes of temperature (25–70°C). In order to study the effect of temperature on the stability of the enzymes, α-amylase and glucoamylase were incubated at various temperatures (25 –70°C) for 1 hour at pH 5.9 and 4.4, respectively, and then rapid cooling in ice-bath.
| Table 1: | Partial purification of S. Cerevisiae YKU 107 α-amylase |
 | |
| Table 2: | Partial purification of S. Cerevisiae YKU 131 glucoamylase |
 | |
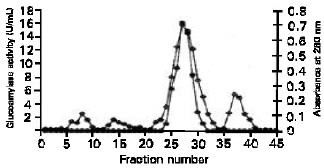 | |
| Fig. 1: | The elution profile of S. Cerevisiae YKU 107α-amylase from DEAE-cellulose column. (●) α-amylase activity; (◊) Absorbance at 280 nm |
After each incubation, the residual activity was estimated by standard assay procedure. Figure 6 shows that optimum temperature of α-amylase was around 45-50 oC with heat stability up to 40 oC. On the other hand, the glucoamylase optimally active at temperature 50-55 oC and the heat stability was up to 45 oC. Figure 7 shows the effect of temperature on activity and stability of glucoamylase.
Hydrolysis of starches: The hydrolysis of various starches (sago, potato, rice, cassava and corn starch) by glucoamylase and α-amylase was investigated. For α-amylase, the incubation was performed at 40°C with 0.05 M acetate buffer at pH 5.0, containing 20 g/L of gelatinised starch and 23 U/mL of enzyme activity. After different time intervals of incubation, samples were taken and the residual starch remained was estimated.
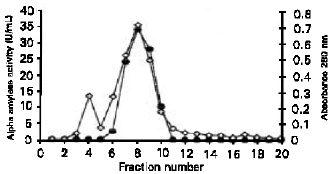 | |
| Fig. 2: | The elution profile of S. Cerevisiae YKU 131 glucoamylase from DEAE-cellulose column. (●) α-amylase activity; (◊) Absorbance at 280 nm |
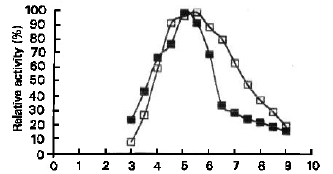 | |
| Fig. 3: | Effect of pH on S. Cerevisiae YKU 107 α-amylase activity ( ■ ) and stability ( □ ) |
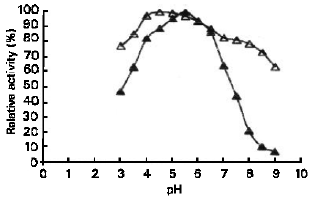 | |
| Fig. 4: | Effect of pH on S. Cerevisiae YKU 132 glucoamylase activity (▲) and stability (Δ) |
For glucoamylase, the incubation was performed at 40°C with 0.05 M acetate buffer at pH 5.5, containing 20 g/L of gelatinised starch and 16 U/mL of enzyme activity. After different time intervals of incubation, samples were taken for residual starch estimation.
Figure 8 shows that α-amylase hydrolysed the potato starch most rapid, followed by rice, cassava, sago and corn starch. There is no significant different in percentage of hydrolysis of potato, rice and cassava starch after 24 hours of incubation, which was approximately 89.50%.
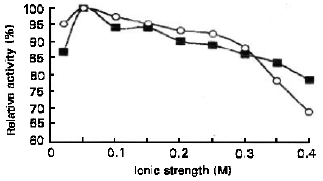 | |
| Fig. 5: | Effect of ionic strength (sodium acetate) at 40°C on the enzyme activity of α-amylase ( ■) and glucoamylase (○) |
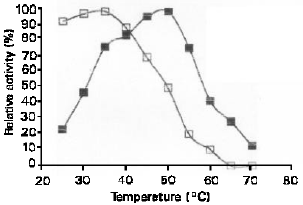 | |
| Fig. 6: | Effect of temperature on the activity (■) and stability (□ ) of S. Cerevisiae YKU 107 α-amylase |
 | |
| Fig. 7: | Effect of temperature on the activity (▲) and stability (Δ) of S. Cerevisiae YKU 131 glucoamylase |
The percentage hydrolysis of sago and corn starch after 24 hours of incubation were 86.30% and 64.63%, respectively.
Figure 9 shows the hydrolysis of various starches by S. cerevisiae YKU 131 glucoamylase. The glucoamylase hydrolysed the cassava starch most rapid, followed by sago, potato, corn and rice starch.
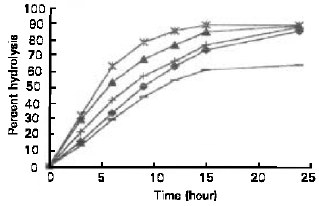 | |
| Fig. 8: | Hydrolysis of various starches by S. Cerevisiae YKU 107 α-amylase. (♦) Sago, (▲) Rice, (*) Potato, (+) Cassava, (–) Corn |
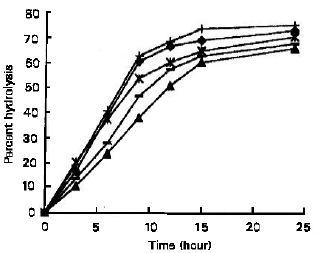 | |
| Fig. 9: | Hydrolysis of various starches by S. Cerevisiae YKU 131 glucoamylase. (♦) Sago, (▲) Rice, (*) Potato, (+) Cassava, (–) Corn |
After 24 hours of incubation, the percent hydrolysis of cassava, sago, potato, corn and rice starch were 75.73, 73.58, 71.01, 68.35 and 66.45 %, respectively.
DISCUSSION
For α-amylase, the optimum pH was 5.0, and the enzyme was stable at pH range between 4.5 and 6.0. Thus the pH should be controlled at 4.5 – 6.0 for the maximum enzyme activity. The glucoamylase was stable at pH 4.0 – 6.0, the stability decrease gradually with the increase of pH. The enzyme was optimally active at pH 5.5. The ionic strength at the range of 0.05 M to 0.20 M did not severely affect the activity of the both enzymes. Thus the pH in an uncontrolled fermentation process could be maintain by increasing the buffering capacity of the sodium acetate buffer at ionic strength lower than 0.2 M. Previous studies reported that S. cerevisiae could grow at pH 2.4 to 8.6 (Jones et al., 1981), therefore, the pH values selected for fermentation process would primarily depend on the optimum pH of the amylolytic enzymes.
Since the host strain of the recombinant yeast (YKU 107 and YKU131 strains) were temperature sensitive mutants, which growth optimally at 30°C. At temperature 30°C, the relative activity of glucoamylase and α-amylase was 57.73% and 47.60%, respectively. Previous studies shown that S. cerevisiae YKU 107 produced the α-amylase optimally at temperature 34°C (Ujiyama et al., 1995), while S. cerevisiae YKU 131 (produced glucoamylase), 35°C (Lee et al., 1998). At this optimum enzyme expressing temperature, relative activity of α-amylase and glucoamylase were approximately 76.94 % and 68.43 % respectively. Thus, controlling the temperature using optimum production strategy to enhance the amylolytic enzymes expression (Ujiyama et al., 1995; Lee et al., 1998), might be essential in maximising the starch utilisation during fermentation process.
The α-amylase and glucoamylase were capable in hydrolysing various types of starches at different rates and degrees. However, it is too soon to conclude which starch is the most suitable for ethanol production using the particular recombinant yeasts. This is because in fermentation process, the different viscosity of different starches might affect the growth and the amylolytic enzymes expression of the recombinant yeasts.
The pH that suitable in fermentation process by S. cerevisiae YKU 107 and YKU131 were 4.5 – 6.0 and 5.0 – 6.0 respectively. The optimum amylolytic enzyme production strategy might be important in maximising the ethanol production directly from starches. The α-amylase could hydrolyse more than 85% of the starches, except for corn, 64.63%. Glucoamylase was capable in hydrolysing more than 65% of the starches. In the view of the susceptibility of the starch to amylolytic enzyme hydrolysis, sago starch is compatible as carbon source for ethanol production by the recombinant yeasts.
ACKNOWLEDGEMENT
The authors are grateful to the National Biotechnology Directorate, the Ministry of Science, Technology and Environment, Malaysia for the financial support towards this research project under the “Top-Down” Biotechnology Program and to the Japan Society for the Promotion Science (JSPS Group 3) under the Large Scale Co-operative Research program in Biotechnology (JSPS-NRCT/DOST/LIPI/VCC).
REFERENCES
- Hartree, E.F., 1972. Determination of protein: A modification of the lowry method that gives a linear photometric response. Anal. Biochem., 48: 422-427.
CrossRefPubMedDirect Link - Rossillo-Calle, F., D.O. Hall, A.L. Arora and J.O.B. Carioca, 1992. Bio-Ethanol Production: Economic and Social Considerations in Failure and Success. In: Biotechnology: Economic and Social Aspects, E.J. Da Silva, C. Ratledge and A. Sasson (Eds.). Cambridge University Press, Cambridge, London, pp: 23-54.