
Simeen Mansoor
Department of Genetics, University of Karachi, Karachi-75270, Pakistan
Farzana N. Naqvi
Department of Genetics, University of Karachi, Karachi-75270, Pakistan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 10 | Page No.: 1550-1552







ABSTRACT
A study of wall bound invertases in different tissues of mung bean cultivar has been undertaken. In young and mature leaves, acid as well as alkaline invertase activity was prominent. Senescent leaves showed the presence of alkaline invertase only. During flowering and fruiting, flowers and pods both posses acid invertase activity at initial stages of development whereas closed flower and mature pods showed the presence of alkaline invertase. Present results revealed that the young tissues of mung bean required acid invertase activity for their growth and development where hexoses were rapidly utilized with rapid sucrose hydrolysis. In mature and senescent tissues the appearance of alkaline invertase indicate the involvement of enzymes in the storage of sucrose. During present investigations, isozymes of invertases were studied by eluting them in sucrose solution of their respective pH optima after performing 7.5% polyacrylamide gel electrophoresis. Each tissue appears to possess wall bound invertases with predominance of a different activity in each case.
PDF Abstract XML References Citation
How to cite this article
Simeen Mansoor and Farzana N. Naqvi, 2000. Changes in Wall Bound Invertase Activity in Young and Mature Tissues of Mung Bean (Vigna radiata). Pakistan Journal of Biological Sciences, 3: 1550-1552.
DOI: 10.3923/pjbs.2000.1550.1552
URL: https://scialert.net/abstract/?doi=pjbs.2000.1550.1552
DOI: 10.3923/pjbs.2000.1550.1552
URL: https://scialert.net/abstract/?doi=pjbs.2000.1550.1552
INTRODUCTION
β-Fructofuranosidse fructohydrolase (EC 3.2.1.26) or invertase enzyme is responsible for the hydrolysis of sucrose into glucose and fructose (Ouelhazi et al., 1992). It often occurs in multiple forms deferring in pH optima, isoelectric points and subcollular locations in many tissues (Pressey, 1994). Wall bound or soluble fractions may contain acid, alkaline or neutral invertases. Acid enzyme catalysis the irreversible cleavage of sucrose to glucose and fructose (Isla et al., 1999), while alkaline enzyme hydrolysed sucrose in cell of storage organs (Dey, 1985). Neutral invertase catalysis the hydrolysis of terminal non-reducing β-fructofuranoside residues in β-D-fructofuranosides like sucrose (Vorster and Botha, 1998). This paper is an attempt to perform comparative study of the cell wail invertases from different tissues of mung bean at various developmental stages.
MATERIALS AND METHODS
Seeds of cv. 19-19 of mung bean (Vigna radiate) were obtained from Pakistan Agricultural Research Council, Islamabad. Seeds were sown in the field in randomized complete block design with three replications. Third leaf was collected from young, blooming, mature and senescent. Bud and closed flower (preanthesis) were taken as two different developmental stages of flower, whereas immature, young and mature pods were the three different stages of pod.
From these tissues, wall bound fraction was obtained by the method of Vattuone et al. (1981). pH optima from these tissues was detected by the method of Prado et al. (1980). Then 7.5% polyacrylamide gal electrophoresis (PAGE) was performed according to the method of Maurer (1971) with minor modifications. After PAGE, each gel was cut into tan pieces of 0.5 cm. Each piece was then placed in 1 ml of 0.1 M acetate buffer and 0.2 M sucrose solution to elute enzyme in the solution. These tubes were shaken vigorously and incubated overnight at 4°C. Invertase activity (Erg sugar/mg proteins/30 minutes) was estimated by measuring the amount of reducing sugars after enzymatic hydrolysis of sucrose by the method of Nelson (1944). Graph was plotted, each peak will represent an isozyme.
RESULTS AND DISCUSSION
A major concern of investigators interested in cellular differentiation is to discern the biochemical steps that accompany the development of cells. The isozymic pattern of plant organs and tissues may be a direct reflection of particular genes that are operating at a given stage of growth and differentiation (Trinh et al., 1981).
In the present investigations, various tissues of mung bean were taken from different developmental stages and wall bound invertase activity (μg sugar/mg proteins/30 minutes) was observed in buffer of different pH (3.0-8.5). invertase isozymes at respective pH optima were detected.
pH Optima of Various Tissues: During present work, young leaf possessed maximum invertase activity at acidic pH and alkaline pH (Fig. 1). Leaf at blooming stage possessed acidic and neutral invertase. Leaves at mature stage had acidic and alkaline invertase, whereas senescent leaf contained neutral and alkaline invertase activity with no acid invertases, according to Isla et al. (1988) in the absence of alkaline invertase, neutral invertase take over the role of acid soluble invertase. It was noted that acid invertase persist within young, blooming and mature stages of leaf whereas there was appearance and disappearance of alkaline and neutral invertases. Presence of soluble invertase with pH optima of 7.2 in mature sugarcane stem tissue was also reported by Vorster and Botha (1998).
Bud and closed flower showed highest peak at acidic and at alkaline pH (Fig. 2). Immature pod has maximum invertase activity at acidic pH . Young and mature pod possessed only alkaline invertase (Fig. 3). During present studies it was observed that acid invertase was present at early stages of development in leaves, flowers and pods, whose presence indicated the hydrolysis of sucrose into glucose and fructose for carbon and energy source (Krishnan et al., 1985). In sugar cane, a high acid invertase in young stem with low sucrose concentration was also reported by Venkataramana and Naidu (1993). As matured tissues posses either no or very low acid invertase activity and the absence of acid invertase activity may indicate sucrose storage, also reported by (Isla et al., 1988).
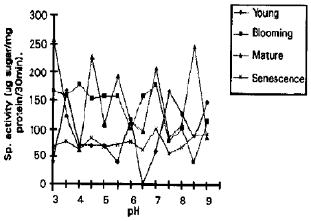 | |
| Fig. 1: | Invertase activity, pH profile at different stages of leaf in cv, '19-19' |
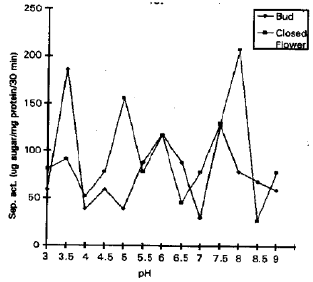 | |
| Fig. 2: | Invertase activity, pH profile at different stages of flower in cv, '19-19' |
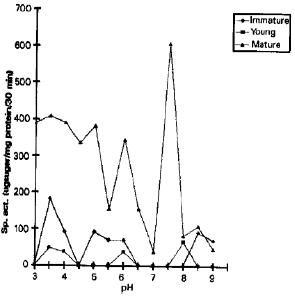 | |
| Fig. 3: | Invertase activity, pH profile at different stages of pod in cv, 19-19 |
 | |
| Fig. 4: | Invertase isozymes in leaves at their specific pH optima |
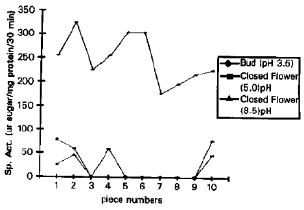 | |
| Fig. 5: | Invertase isozymes in flower at respective pH optima |
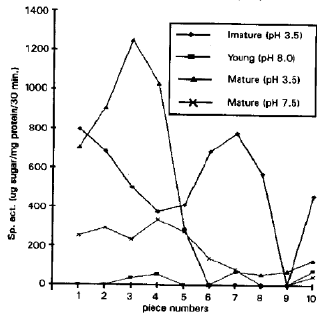 | |
| Fig. 6: | Invertase isozymes in pod at their respective pH optima |
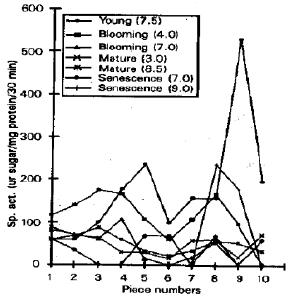
Invertase Isozymes at their Specific pH Optima: Another way of detecting invertase isozymes is to elute the enzyme in the sucrose solution from the gel pieces (after PAGE) and then the amount of sucrose hydrolysed by the enzyme could be detected, each peak will indicate an isozyme (Humphreys and Echeverria, 1980). Figure 4 showed that young leaf possessed acid and alkaline invertases however data of acid invertase was not reported. Young leaf contained lv1, lv2, lv5 lv6, lv8 and Iv10 alkaline isozymes. Leaf at blooming stage have lv1 to Iv9 acid isozymes and Iv1, lv2, lv3, Iv4, lv5, lv7, Iv8 and Iv9 neutral isozymes. Mature leaf had acid and alkaline enzyme activity in all ten pieces with different intensities. In senescent leaf all the ten neutral invertase isozyrnes of different quantities were present however only two alkaline isozymes i.e. Iv8 and Iv9 were present. In case of leaves, Iv1, lv2, Iv5 and Iv8 were present in all the stages.
Flower at bud stage had Iv1 to lv10 acid isozymes, with no neutral or alkaline invertases. Closed flower showed both acidic and alkaline invertase activity, in which lv1, lv2, lv4 and lv10 acid isozymes and lv1, Iv2 and lv10 alkaline isozymes were present (Fig. 5). In flowers lv1, Iv2 and lv10 were present in both the developmental stages, Whereas Iv3, Iv5, Iv6, lv7, lv8 and Iv9 were specific to bud stage only.
Pods at immature stage possessed invertase activity only at acid pH where lv1 to Iv8 and lv10 were present. In young pod iv3, lv4 and lv10 alkaline isozymes were present with no neutral or acidic activity. Mature pod had both acid and neutral invertases, Iv1, Iv2, Iv3, lv4, lv5, lv7, lv8, lv9 and lv10 acid isozymes were present whereas Iv1 to Iv7 and lv10 alkaline invertases were present (Fig. 6). In pod, lv3 and lv10 were present in all stages.
In the present studies, expression of invertase isozymes was detected from various tissues at different devlopmental stages. It was noted that acid invertase was present in the young stages and persists for longer period. It was also observed that lv1, lv2 and lv10 is present almost in all the tissues at all the pH optima so we may considered it as house keeping isozyme.
REFERENCES
- Vorster, D.J. and F.C. Botha, 1998. Partial purification and characterization of sugarcane neutral invertase. Phytochemistry, 49: 651-655.
CrossRefDirect Link - Dey, P.M., 1985. Changes in the forms of invertase during germination of mung bean seeds. Phytochemistry, 25: 51-53.
CrossRefDirect Link - Humphreys, T. and E. Echeverria, 1980. Invertase and maltase in the free space of the maize scutellum. Phytochemistry, 19: 189-193.
CrossRefDirect Link - Isla, M.I., M.A. Vattuone, R.M. Ordonez and A.R. Sampietro, 1999. Invertase activity associated with the walls of Solanum tuberosum tubers. Phytochemistry, 50: 525-534.
CrossRefPubMedDirect Link - Isla, M.I., M.A. Vattuone, M.I. Gutierrez and A.R. Sampietro, 1988. Acid invertase from Tropaeolum leaves. Phytochemistry, 27: 1993-1998.
CrossRefDirect Link - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
CrossRefDirect Link - Ouelhazi, N.K., R. Ghrir, K.D. Le and F. Lederer, 1992. Invertase from Opuntia ficus-indica fruits. Phytochemistry, 31: 59-61.
CrossRefDirect Link - Prado, F.E., O.L. Fleischmacher, M.A. Vattuone and A.R. Sampietro, 1980. Cell wall invertases of sugar cane. Phytochemistry, 21: 2825-2828.
CrossRefDirect Link - Trinh, T.H., T. Gaspar, K.T. Thanh Van and J.L. Marcotte, 1981. Genotype, ploidy and physiological state in relation to isoperoxidases in Nicotiana. Physiol. Plant., 53: 153-157.
CrossRefDirect Link - Vattuone, M.A., F.E. Prado and A.R. Sampietro, 1981. Cell wall invertases from sugar cane. Phytochemistry, 20: 189-191.
CrossRefDirect Link - Venkataramana, S. and K.M. Naidu, 1993. Invertase-sucrose relationship in young and mature stem of sugarcane. Phytochemistry, 32: 821-822.
CrossRefDirect Link