
Marwa A. Masoud
Department of Pharmacology, National Organization for Drug Control and Research (NODCAR), Giza, Egypt
Omar A. Ahmed-Farid
Department of Physiology, National Organization for Drug Control and Research (NODCAR), P.O. Box 29, Giza, Egypt
LiveDNA: 20.31381
Hanan A. Rizk
Department of Pharmacology, National Organization for Drug Control and Research (NODCAR), Giza, Egypt
Journal of Pharmacology and Toxicology
Year: 2020 | Volume: 15 | Issue: 2 | Page No.: 44-57







ABSTRACT
Background and Objective: D-galactose induced neurotoxicity is widely known model for survey aging and related oxidative neurotoxicity and memory impairment. The present study was conducted to explore the possible role of Linseed Oil (LO) and Coenzyme Q10 (CoQ10) as compared with standard neuroprophylactic Ginkgo biloba extract to protect hippocampal cells against toxic effects induced by D-galactose (D-gal). Materials and Methods: The rats (50 males) were divided randomly into five groups, 10 rats for each, the 1st normal control, the 2nd served as aging treated with D-gal, the 3rd treated with Ginkgo biloba plus D-gal, the 4th group treated with linseed oil plus D-gal, the 5th group treated with Coenzyme Q10 plus D-gal. The experiment lasted for 6 weeks then behavioral tests were examined and then all animals were decapitated for biochemical, neurochemicals, histopathology and immunohistochemistry analysis. Results: Administration of D-gal for 6 weeks significantly impaired behavioral test, oxidative defense, decreased endogenous antioxidant, impaired neurotransmitters contents, increased acetylcholinesterase, inhibited neurogenesis, increased 8-hydroxy-2-deoxyguanosine, decreased ATP and increased apoptotic factors as compared to normal group. Six weeks of linseed oil and coenzyme Q10 treatments significantly improved neurobehavioral alterations, brain derived neurotrophic factor and apoptosis as compared to D-gal. Conclusion: The present study concluded that dietary intake rich with D-gal disrupt neuronal cell structure in accordance with aging which accumulate most neuronal aberration and treating with natural substances may be beneficial in normal aging troubles. In addition, CoQ10 showed best protective effect after 6 weeks of treatment compared with LO and GB which normalized most selected paramours.
PDF Abstract XML References Citation
Received: November 27, 2019;
Accepted: January 20, 2020;
Published: March 15, 2020
How to cite this article
Marwa A. Masoud, Omar A. Ahmed-Farid and Hanan A. Rizk, 2020. Neuromodulatory Influences of Nutraceuticals on D-galactose Induced Hippocampal Neuronal Metabolic Dysfunction Model. Journal of Pharmacology and Toxicology, 15: 44-57.
DOI: 10.3923/jpt.2020.44.57
URL: https://scialert.net/abstract/?doi=jpt.2020.44.57
DOI: 10.3923/jpt.2020.44.57
URL: https://scialert.net/abstract/?doi=jpt.2020.44.57
INTRODUCTION
Aging is a progressive biological activity associated with multiple structural and biochemical alteration in biological system in addition to cognitive impairment1. Reactive Oxygen Species (ROS) induced DNA injury which is the main cause of aging2,3. There are multiple animal models for aging research such as; D-galactose (D-gal) model is convenient and can be resemble to natural aging researches. D-galactose metabolized normally in the body by D-galactokinase and galactose-1-phosphate uridyl transferase. However, chronic administration of D-gal can be converted into hydrogen peroxide leading to the formation of a superoxide anion4. Administration of D-gal shows aging symptoms such as; impairment of spatial learning, memory, decline in cognitive functions, neuroinflammation and increase apoptosis5,6. Hippocampus is responsible for learning, memory function and age-related neurodegeneration7. Therefore, dietary supplements and foods rich in natural antioxidants were shown to be beneficial in aging disorders8.
Linseed “Flaxseed, Linum usitatissimum” has been the focus of increased interest in the field of diet and disease research due to its Omega-3 polyunsaturated fatty acids content. It is also the richest natural source of mammalian lignans that effectiveness is through an antioxidant effect9.
Nutritionally, linseed oils rich with α-linolenic acid (ALA) which considered an essential fatty acid used as dietary supplements10. Thus, may benefit human body as antiarrhythmic and neuroprotective functions11. Up to date, no studies focused on the neuroprotective role of linseed oil against D-gal induced aging alterations and neurotoxicity.
Coenzyme Q (CoQ, ubiquinone) is a unique lipid-soluble antioxidant that is generated in animals12. It is an essential compound of the mitochondrial electron transport chain and is therefore essential for the production of ATP13. It is an important controller of lifespan in normal aging14. It is chemically similar to vitamin K in its structure, but it is not considered a vitamin because it is synthesized in the body. Furthermore, CoQ10 is a powerful antioxidant capable of recycling and restore other antioxidants15 such as; vitamin E and vitamin C.
The present research was conducted to study dietary intake of natural substances that could be beneficial in normal aging troubles and to explore the potential role of linseed oil and coenzyme Q10 as compared with standard neuroprophylactic Ginkgo biloba extract to protect hippocampal cells against toxic effects induced by D-galactose.
MATERIALS AND METHODS
This study was conducted in September, 2017 in Animal house of National Organization for Drug Control and Research, Giza, Egypt.
Animals: The adult male Sprague-dawley rats weighing 280-300 g were used in the present study. The animals were obtained from animal house of NODCAR and caged in 5 separate cages with natural ventilation and illumination at about 20±2°C and 14 h light period and 65% humidity, animal feed standard basal diet (ad libitum) and free access of water. Animals were allowed one week for adaptation before treatments. Animal handling, tissue collection were follow the instructions of the guidelines of the Research Ethical Committee of the NODCAR.
Chemicals: D-galactose was purchased from Loba Chemie, India and used in dose of D-gal (120 mg kg–1) daily, sub-cutaneously (sc), for 6 successive weeks7. Standardized EGb 761 (Huisong, China) was used in dose of 150 mg kg–1 b.wt., according to Blecharz-Klin et al.16. Linseed oil (Isis Company for Food and Industries, Egypt) was used (LO, 1.5 g kg–1; p.o, for 6 weeks), according to Rajesha et al.17. Coenzyme Q10 (Arab Company for Pharmaceutical and Medicinal Plant; MEPACO, Egypt) was used (CoQ10, 10 mg kg–1; p.o, for 6 weeks), according to Emam et al.18.
Experimental design: The protocols of following methodologies were adopted according to the guidelines of the Institutional Animal Ethics Committee of NODCAR. All the experimental procedures were carried out in accordance with international guide lines for the care and use of laboratory animals and with accordance with standard guidelines19. The rats were subjected to 50 male adult Sprague-dawley were divided randomly into five groups, 10 rats for each, the 1st served as normal control the 2nd served as control aging treated with D-galactose (D-gal, 120 mg kg–1, subcutaneously) the 3rd treated with Ginkgo biloba (GB, 150 mg kg–1, p.o.) plus D-gal, the 4th group treated with Linseed Oil (LO, 1.5 g kg–1, p.o) plus D-gal, the 5th group treated with coenzyme Q10 (10 mg kg–1, p.o.) plus D-gal. The experiment lasted for 6 weeks then behavioral tests were examined and then all animals were decapitated for biochemical, neurochemicals and histopathology and immunohistochemistry analysis.
Behavioral assessments
Measurement of spatial memory and learning by Morris Water Maze (MWM): Morris water maze20 analyze its behavior via 2 parameters 1st is escape latency, which is the time it takes to find the platform, the 2nd measured during probe trials: the escape platform is removed and the mice or rats are allowed to search for it for a fixed time (often 60 sec). Variables were measured time and path length in quadrants, time near platform and platform crossings.
Grid test (Catalepsy test): The grid test was measured according to Rahman et al.21 and used as index of catalepsy or sensorimotor deficit.
Assessment of locomotor and exploratory activity by the open field test: The open-field test was measure rearing and ambulation according to Looser22.
Tissue sampling: Tissue sampling were collected after 24 h of the end experiment and the rats were decapitated and both hippocampi were isolated one was kept in 10% formalin for histopathological examination while the other at -80°C for estimating the other biochemical parameters. For the determination of neurotransmitters, a 10% (w/v) homogenate was prepared in a 75% methanol for HPLC. Each homogenate was centrifuged at 10006×g (4°C) for 10 min. The resultant supernatant was divided into 2 halves, the first was dried using vacuum (70 millipore) at RT and its residues were derivatized for the determination of brain amino acids (HVA, GLU, GABA), whereas the second half was used for monoamines determination (5-HT, DA, NE). In another subset, hippocampi were homogenized in 10% (w/v) phosphate buffer (pH 7.6) for the assay of the other biochemical parameters23.
Determination of malondialdehyde (MDA), reduced glutathione (GSH), Nitric Oxide (NO) contents, superoxide dismutase (SOD) activity and Total Antioxidant Capacity (TAC) in hippocampal brain area: Hippocampal MDA, GSH, NO, SOD and TAC were measured according to the methods of Prins and Loose24, Miranda et al.25, Nishikimi et al.26 and Koracevic et al.27, respectively.
Determination of hippocampal Brain Derived Neurotrophic Factor (BDNF) content: Enzyme Linked Immunosorbent Assay (ELISA) was used to determine hippocampal BDNF by using a test reagent kit (CUSABIO Biotech co., USA), according to the manufacturer’s instructions.
Determination of hippocampal amino acids: Hippocampal free amino acids, monoamines and their metabolites were measured by HPLC UV detector28,29.
Determination of hippocampal acetylcholinesterase (AChE) activity: Hippocampal AChE was determined by using DTNB-phosphate reagent after 10 min incubation of the hippocampal homogenate with acetyl thiocholine iodide30.
Determination of hippocampal 8-OHDG content: Hippocampal 8OHdG was measured by HPLC UV detector31.
Determination of hippocampal adenosine tri-phosphate (ATP) content: Hippocampal ATP was measured by HPLC UV detector32.
Histopathological and immunohistochemical examinations of Bax and Caspase-3 (Casp-3) For histological study, hippocampal brain area were cut and processed for paraffin sections of 5 μm thickness to be stained with H&E, according to Bancroft et al.32. Immunohistochemical detection of caspase-3 was performed by using primary rabbit anti-rat caspase-3 antibody from Neo Markers Fremont CA, Lab Vision. The steps were applied instead of the primary antibodies33.
Statistical analysis: All values were presented as Mean±SEM. Statistical analysis was performed by using GraphPad Prism version 5 (Graph-Pad, San Diego, CA). A comparison between different groups was carried out using one-way analysis of variance (ANOVA) followed by Tukey–Kramer multiple comparisons test. A significant differences were considered when probability less than 0.05.
RESULTS
Behavioral tests for proofing metabolic syndromes associated with aging: The Morris water maze, grid and open field test were applied for determine neuronal metabolic syndrome affected by D-gal and obviate its effects concurrent with natural remedy.
Morris Water Maze (MWM) training course: There were no significant differences between groups in the mean escape latency on the first day of training. On the second, third and fourth days of investigation, D-gal increased escape latency time (p<0.05) compared to the control group (Fig. 1a). In contrast, treated group showed markedly amelioration and differ from D-gal group after 1st trail for 2nd, 3rd and 4th test, but didn’t differ from normal group except GB group at 3rd and 4th group. In addition, treated group with D-gal deteriorated memory test resembling in the decreasing of swimming time at the selective square of platform (Fig. 1b).
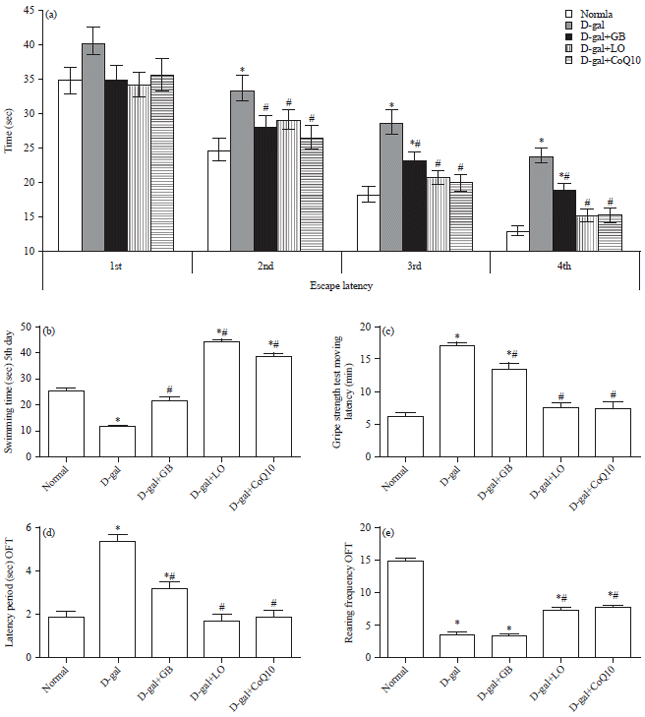 | |
| Fig. 1(a-e): | Behavioral effects of Linseed Oil (LO) or Coenzyme Q10 (CoQ10) and Ginko biloba (GB) extract on D-galactose (D-gal)-induced brain aging in rats on morris water maze test (a) Escape latency and (b) Swimming time, (c) Moving latency, (d) Open field test and latency time and (e) Rearing frequencies |
Each bar represents means (n = 10)±standard error. Statistical analysis was carried out by one way ANOVA followed by Tukey-Kramer multiple comparison test. *Significantly different from normal group at p<0.05, #Significantly different from control (D-gal) group at p<0.05 | |
On the other hand, treated groups with GB, LO and CoQ10 showed significant amelioration via increase swimming time in comparing with D-gal group, but LO and CoQ10 showed superior amelioration from control group. In the grid test, as illustrated in Fig. 1c, the administration of D-gal to rats induced significant increase in catalepsy score of grid test compared to the normal group. Administration of GB and D-gal decreased moving latency of grid test compared to D-gal-treated group. Concomitant administration of either LO plus D-gal or CoQ10 plus D-gal induced significant normalize catalepsy score, respectively as compared to D-gal treated group. In the OFT, as illustrated in Fig. 1d-e, administration of D-gal significantly increased latency time and decrease the rearing respectively in comparing with normal group.
| Table 1: | Neuroprotective effect of linseed oil as compared with Ginko biloba on oxidative stress parameters in hippocampal homogenate in D-galactose |
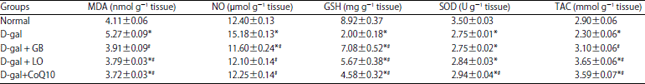 | |
Each value represents means (n = 10)±standard error. Statistical analysis was carried out by one way ANOVA followed by Tukey–Kramer multiple comparison test. *Significantly different from normal group at p<0.05. # Significantly different from control (D-gal) group at p<0.05, GB: Ginko biloba, LO: Linseed oil, CoQ10: Coenzyme Q10, MDA: Malondialdehyde, NO: Nitric oxide, GSH: Reduced glutathione, SOD: Superoxide dismutase, TAC: Total antioxidant capacity | |
In contrast, treated group with GB, LO and CoQ10 significantly decreased the latency and increase the rearing respectively compared to D-gal group except GB didn’t differ for rearing only. Finally, treated group with LO and CoQ10 showed significant amelioration resembling in latency period only, but the other parameter and groups showed markedly disrupt via increase the latency and decrease rearing in comparing with normal group.
Hippocampus MDA, NO, GSH, SOD and TAC contents: As shown in Table 1, D-gal induced a significant increase in hippocampus MDA and NO content to be 5.27±0.09 and 15.18±0.13, respectively with significant decrease in hippocampus GSH, TAC contents and SOD activity respectively as compared with normal rats. Administration of GB, LO and CoQ10 significantly decrease the MDA (3.91±0.09, 3.79±0.03 and 3.72±0.03) and NO contents (11.60±0.24, 12.10±0.13 and 12.25±0.14), respectively as compared with aging rats. While, GB, LO and COQ10 significantly increased GSH content (7.08±0.52, 5.66±0.38 and 4.58±0.32), respectively and TAC by means of 3.10±0.06, 3.65±0.06 and 3.59±0.07 respectively, however only CoQ10 nearly normalize SOD content as compared with aging animals..
Hippocampus BDNF content: As illustrated in Fig. 2, D-gal induced a significant decrease in hippocampus BDNF content (1.48±0.03) as compared with normal rats (10.28±0.31). Oral administration of GB, LO and CoQ10 significantly increased the content of BDNF (4.08±0.04, 5.13±0.13 and 7.68±0.35), respectively as compared with aging animals.
Hippocampus neurotransmitters contents: As illustrated in Fig. 3a-f, D-gal deteriorated hippocampus monoamines and amino acids contents were manifested by reduction in 5-HT content (0.38±0.01), DA (1.09±0.03) and NE (0.55±0.03) contents as compared with values of normal rats. This reduction accompanied by significant increment in GABA (5.50±0.11), GLU (1.12±0.02) and HVA (0.49±0.02) contents as compared with normal rats, respectively.
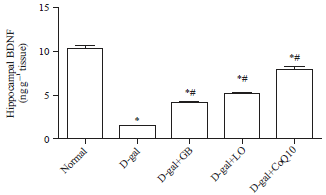 | |
| Fig. 2: | Neuroprotective effect of Linseed Oil (LO), Coenzyme Q10 (CoQ10), and Ginko biloba (GB) extract on hippocampal Brain Derived Neurotrophic Factor (BDNF) in D-galactose (D-gal) treated rats |
Each bar represents means (n = 10)±standard error. Statistical analysis was carried out by one way ANOVA followed by Tukey-Kramer multiple comparison test, *Significantly different from normal group at p<0.05, #Significantly different from control (D-gal) group at p<0.05 | |
Administration of CoQ10 markedly increased the reduced hippocampus contents of 5-HT (0.74±0.04), DA (1.47±0.06) and NE (0.92±0.04) accompanied with significant decrease in GABA (4.51±0.11), GLU (0.87±0.01) and HVA (0.23±0.01) contents as compared with aging animals. Moreover, treatment of aging animals with GB markedly increased the reduced hippocampus contents of DA (1.46±0.09) with significant decrease in GABA, GLU and HVA contents as compared with D-gal treated animals. In addition, treatment of aging rats with LO significant decrease in GABA, GLU and normalized HVA contents as compared with D-gal treated animals.
Hippocampus AChE content: As illustrated in Fig. 3g, D-gal markedly increased hippocampus AChE content as compared with that of normal rats. In contrast, administration of LO and CoQ10 similarly markedly decreased the elevated hippocampus AChE content to 1.76±0.02 and 1.76±0.03 as compared with D-gal treated rats.
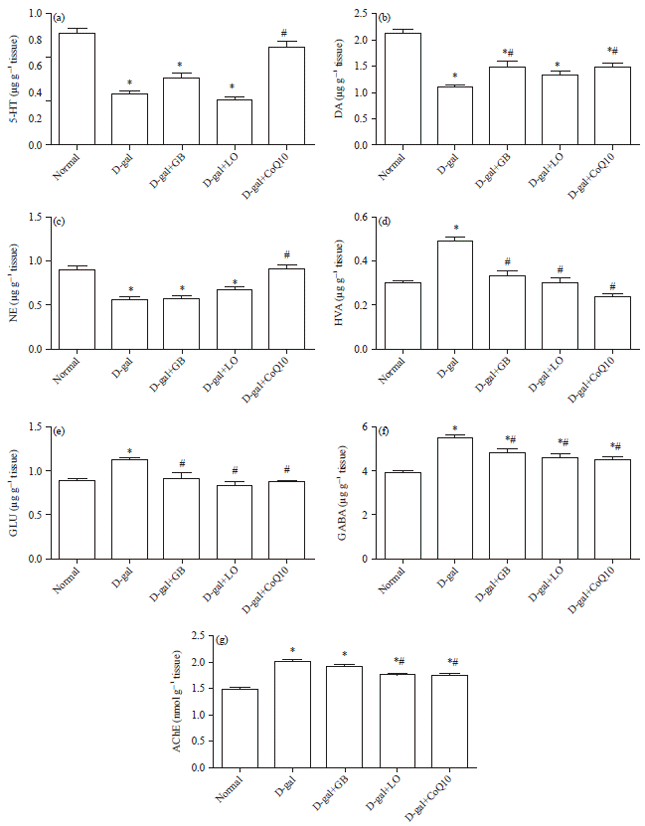 | |
| Fig. 3(a-g): | Neuroprotective effect of Linseed Oil (LO), Coenzyme Q10 (CoQ10), and Ginko biloba (GB) extract on (a) Hippocampal serotonin (5-HT), (b) Dopamine (DA), (c) Norepinephrine (NE), (d) Homovanillic acid (HVA), (e) Glutamate (GLU), (f) Gamma aminobutyric acid (GABA) and (g) Acetylcholinesterase (AChE) contents in D-galactose (D-gal) treated rats |
Each bar represents means (n = 10)±standard error. Statistical analysis was carried out by one way ANOVA followed by Tukey-Kramer multiple comparison test, *Significantly different from normal group at p<0.05. #Significantly different from control (D-gal) group at p<0.05 | |
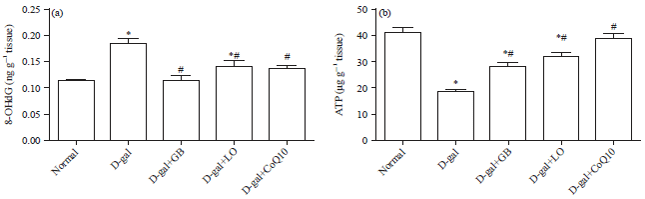 | |
| Fig. 4(a-b): | Please replace title to this :Neuroprotective effect of Linseed Oil (LO), Coenzyme Q10 (CoQ10) and Ginkgo biloba (GB) extract on (a) Hippocampus 8-Hydroxy-2-deoxyguanosine (8-OHdG) and (b) Adenosine tri-phosphate (ATP) contents in D-galactose (D-gal) treated rats |
Each bar represents means (n = 10)±standard error, Statistical analysis was carried out by one way ANOVA followed by Tukey-Kramer multiple comparison test. *Significantly different from normal group at p<0.05. #Significantly different from control (D-gal) group at p<0.05 | |
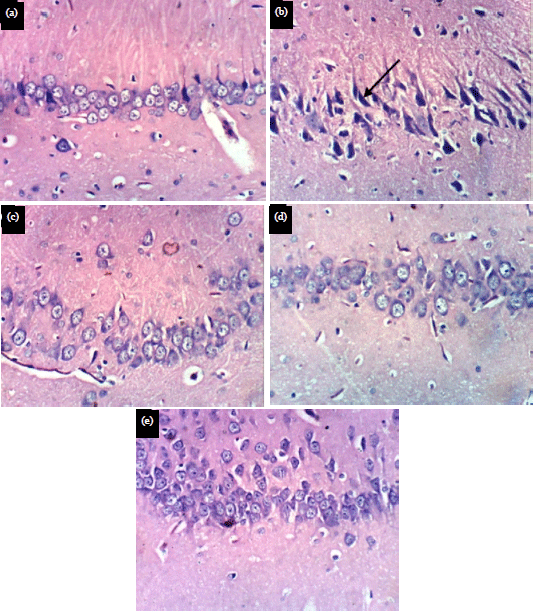 | |
| Fig. 5(a-e): | Photomicrographs of rat hippocampus sections of, (a) Normal control group, (b) D-gel treated and (c-e) GB, LO and CoQ10 treated, stained with H&E (X400) |
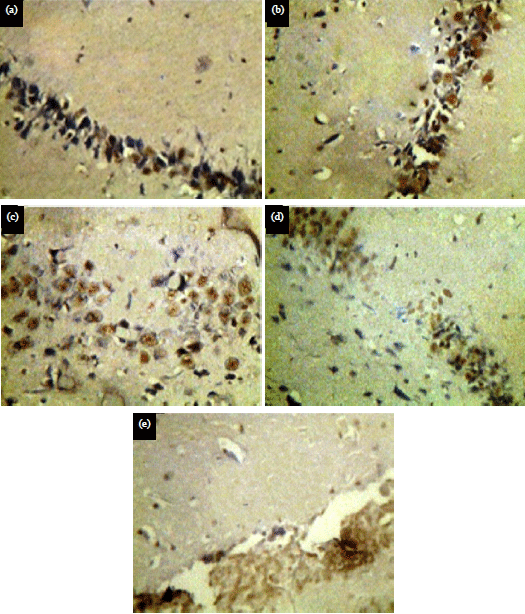 | |
| Fig. 6(a-e): | Photomicrographs staining of box in hippocampus sections of (a) Normal control group, (b) D-gel treated and (c-e) GB, LO and CoQ10 treated, stained with H&E (X200) |
Hippocampus 8-OHdG and ATP contents: As illustrated in Fig. 4a, b D-gal deteriorated hippocampal ATP and 8-OHDG contents were manifested by reduction in ATP accompanied by significant increment in 8-OHDG as compared with normal rat, respectively. Oral administration of GB, LO and CoQ10 induced a significant increase in ATP, respectively accompanied with significant reduction in 8-OHDG, respectively as compared with aging animals.
Histopathological and immunohistochemical examinations of Bax and Caspase-3 (Casp-3): Photomicrographs of rat hippocampus sections stained with hematoxylin and eosin (H&E) were illustrated in Fig. 5a-e. Normal control group showed no histopathological changes (Fig. 5a). Sections of rat
treated with D-galactose showed necrosis and pyknosis of neurons (Fig. 5b). Sections of rat treated with Ginkgo biloba extract, linseed oil and CoQ10, respectively showed normal histopathological appearance (Fig. 5c-e).
Photomicrographs staining of Bax in hippocampus sections are illustrated in Fig. 6a-e, normal control group showed normal expression of Bax (Fig. 6a) and sections of rat treated with D-galactose showed salient raise in the number of positive Bax immunoreactive cells (Fig. 6b). Daily administration of Ginkgo biloba extract showed moderate highly positive expression of Bax which induced by D-galactose (Fig. 6c). Daily administration of linseed oil and CoQ10 showed weak positive expression of Bax, respectively as compared with D-galactose treated rats (Fig. 6d-e).
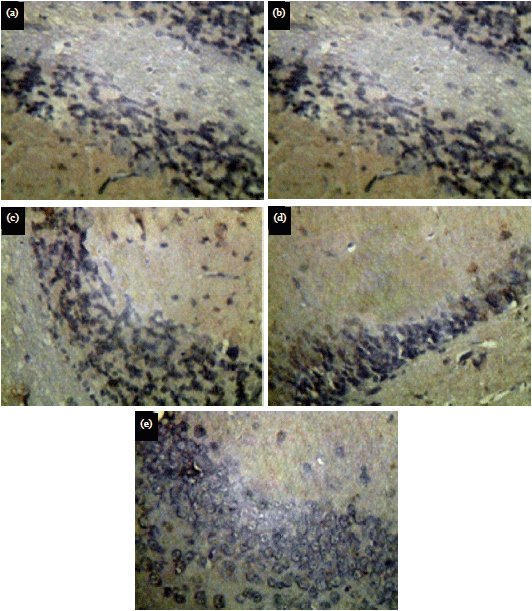 | |
| Fig. 7(a-e): | Photomicrographs staining of Casp-3 (x200) in hippocampus sections of (a) Normal control group, (b) D-gel treated and (c-e) GB, LO and CoQ10 treated, stained with H&E (X |
Photomicrographs staining of Casp-3 in hippocampus sections are illustrated in Fig. 7a-e, normal control group showed normal expression of Casp-3 (Fig. 7a), sections of rat treated with D-galactose showed increment in the number of positive Casp-3 immunoreactive cells (Fig. 7b). Daily administration of Ginkgo biloba extract showed moderate positive expression of Casp-3 which induced by D-galactose (Fig. 7c). Daily administration of linseed oil and CoQ10 showing almost normal expression of Casp-3, respectively as compared with D-galactose treated rats (Fig. 7d-e).
DISCUSSION
According to dietitians, dieting has a significant impact on the body's homeostasis and subsequent to brain and neurotransmitters stability. Therefore, proper nutrition can eliminate signs of aging and maintain public health and in particular nervous system. On the basis of these facts, many studies have shown that carbohydrate treatment, especially D- galactose has a negative effect and causes significant changes, including reduced neuromuscular activity, increased free radical production, reduced antioxidant activity and reduced immune responses and all these changes match the symptoms of natural aging34. D-galactose is a reducing sugar and has the ability to easily interact with free amines and peptides to form Advanced Glycation End-product (AGE). The AGE is increasingly formed during aging and has been associated with the emergence of many diseases such as; diabetes, arteriosclerosis, nephropathy and Alzheimer's disease35. The hypothesis suggested that accumulation of D-galactose may interact with proteins, peptides and increases the composition of AGEs and accelerates the onset of aging. In the present study long term administration of D-gal induced impairment in working and reference memories in senescence rats by spent a longer time in finding the hidden platform during the retrieval trial in the Morris Water Maze (MWM) in task with a decline in coordination skills has been reported as an increased falling rate and weak grip strength on the grid strength test that used to screen for neuromuscular function and remarkably decreased the number of squares passed and prolonged latency to move in the open field test suggested the anxiogenic effects of D-gal when compared with the normal rats36. These finding are in accordance with previous studies1,37-40. Memory and learning impairment is thought to be the result of increased neurodegeneration and decline in the neuronal function. The resulting cognitive decline is a multifactorial process caused by excessive levels of D-gal40 and involves oxidative stress, altered brain neurotransmitters and apoptosis41,35. In this study, obtained data demonstrated that D-galactose, treated rats had a significant decrease in brain monoamines (NE, DA and 5HT) and increase turnover of DA metabolites (HVA). The present data are conferment with previous study of Ho et al.2, who reported that D-gal induced changes that resemble natural aging in rodents, including neurodegeneration and cognitive dysfunction and thus becomes a routinely used method to induce aging in rodents. In addition, excitatory amino acid was markedly increase and in the same way inhibitory amino acid GABA was elevate in comparing with control group although its mechanism needs further research. D-galactose induced a lead to aging progressive deterioration of cell and DNA, accompanied with severity metabolites and subsequent increase 8-OHdG. worthily, 8-OHdG can be seen as results from the mimetic ageing. These finding are in consistent with Evans et al.42, who reported that D-Gal which occupied terminal positions in N-linked oligosaccharides of glycoproteins and play important roles in altering various biological functions which leads to emergence of pathologies in nucleus DNA structure and function. Increase of 8-OHdG formation in brain, may be due to large numbers of mitochondria that are the most reliant upon oxidative phosphorylation43. On the same manner results showed that D-galactose markedly decreases ATP which remarkable for cell energy and dysfunctional of cell carrying capacity in comparing with control group. However, glycation presses didn’t need ATP for lipid or protein deterioration only it the cell energy was depleted or declined. Because the accumulation of redox status progressively exhausting ATP for endogenous balancing reaction. Presented data found that D-gal induced a significant increase in hippocampal MDA and NO content with significant decrease in hippocampal rGSH, TAC contents and SOD activity these results compatible with previous studies1,7,44 and Anand et al.45, who explained that D-gal at the physiological level can metabolize into glucose. So, an over-supply of D-gal is converted into aldos, hydrogen peroxide and formation of advanced glycation end products (AGEs) resulting in production and accumulation of ROS, which is widely recognized as a primary mechanism of natural age-related changes. On the one hand, the brain is the largest organ for oxygen consumption and subjected to deficiency of antioxidant enzymes46. In addition, BDNF encourage the survival and differentiation of neurons, protected neurons against neurodegeneration47. Presented data found that BDNF is decreased in the hippocampus of aging rats and is in agreement with Woo et al.48, who found that repeated administration of D-gal decline BDNF production in the brain resulting in cognitive impairment. In normal aging and increased oxidative stress conditions, memory function declines due to decreases in BDNF48,49. In the present study, D-gal induced hippocampal neurotransmitters deterioration, which may be an important cause of memory decline that appeared with aging. This may be due to increase degradation of monoamine neurotransmitters. These results are compatible with Kou et al.50, who found that AChE increased so decrease cholinergic functions associated with cognitive decline with normal aging. In addition, D-gal raised GLU as Zhou et al.51 who found that GLU increment mediated neurotoxicity and oxidative stress due to reduction of the SOD level that decreased glutamate transporter 1 which removed GLU from synapse. Oxidative damage to DNA has a strong association with the molecular process of aging52,53. Valavanidis et al.54, study showed that 8-OHdG is the general forms of free radical-induced DNA oxidative injury. Obtained results are in accordance with many lines of evidence that emphasize that expression of 8-OHdG was observed centrally in the cytoplasm of cells suggested that D-gal induced DNA oxidative injury in the hippocampus54-57. Apoptosis is one of the mechanisms involved in D-galactose-induced neurodegeneration, these results are compatible with Yu et al.58. Apoptosis is modified by many proteins, so the Bax/Bcl2 ratio is a crucial indicator of apoptosis59, also caspase-3 is known as an apoptosis stimulus60. Administration of D-gal increased the Bax and caspase-3 in hippocampus tissues and apoptosis was developed in these tissues61,62. In the present study, treatments with linseed oil and CoQ10 improved working and reference memories in MWM, muscle coordination as well as locomotor function as compared with D-gal treated rats. At first, linseed oil could improve the D-gal- induced neurotoxic effects may be due to the free radical scavenging activity and its Omega-3 fatty acid content. Taken together presented results are compatible with study of Freemantle et al.63, linseed oil containing ALA is a good substrate for ketogenesis as well as β-oxidation. It has been hypothesized that ALA improved cognitive function by providing the brain with a fast available source of energy and facilitate astrocyte development64. A higher level of antioxidant reserve in linseed oil combined with D-gal treated rats as compared to rats of D-gal group only could be due to reduced oxidative stress because of antioxidant activity of flax lignan complex, so that no need for induction of antioxidant reserve9. The decrease in MDA, NO and 8-OHdG content correlated well with the increase of glutathione. Obtained study, linseed oil increased catecholamine content in brain of rats due to its Omeg-3 fatty acids content that may modify brain function by affecting production and function of neurotransmitters such as 5-HT and DA65. In addition, decreasing in Omega-3 fatty acids predicted decreasing 5-hydroxyindoleacetic acid, the major metabolite of 5-HT, which is known as anti-depressive and Omega-3 fatty acid deficiency is associated with decrements in 5-HT, DA and acetylcholine release66. In the present study, chronic treatment with CoQ10 in D-gal-treated rats significantly attenuated an impairment of spatial learning and memory task performance, CoQ10 is an essential cofactor of the electron transport chain, the neuroprotective effects of CoQ10 lie in the blocking of neuronal lesions produced by the mitochondrial toxin malonate and blocks activation of the mitochondrial permeability transition67. The study observed significant improvement in CoQ10 group which recover the low neurotransmission and energy metabolic concentration in the brain tissue after D-galactose exposure. CoQ10 has been shown to enhance neurotransmitters (monoamines) and reduce their turn-over. Schiefer et al.68 observed increased neurotransmission by activating serotonin receptors in chronic unpredictable mild stress rat model. In addition, Schiefer et al.68 reported that the CoQ10 treatment increased the concentration of 5-HT and NE in diabetic rats model. On the other hand, the use of CoQ10 reduced the activity of high tyrosine hydroxylase lead to increase of NE concentration. In addition, the concentration of the increase of monoamines in paraventricular nucleus in the hypertension model is due to the high activity in the transfer of electrons in the electron transport chain during oxidative phosphorylation and ATP production in the mitochondria69. Swomley et al.70 has linked between an exaggerated production of ROS/RONS and development of hippocampal degeneration. Thus, CoQ10 protected against oxidation of DNA damage and increased lifespan in rats. In addition, other studies have showed a direct association between longevity and mitochondrial levels of CoQ10 in the model of aging-accelerated animals71. Supplementation of CoQ10 increases brain endogenous Q10 content and affords as an antioxidant against free radical generation and has been reported to improve cognitive function, ATP synthesis and provides significant improvement in patients with neurological disorders72. Fischer et al.73 confirmed that a high CoQ10 is accompanied by an increase in muscle strength that in line with results of present study.
CONCLUSION
Present study concluded that, oxidative stress is a major cause in the initiation of neurobehavioral dysfunction in aging. The findings suggested that linseed oil or CoQ10 improved these changes and attenuated the effect of aging on learning and memory in D-gal treated rats. They exhibited beneficial effects on the cognition of aging rats by enhancing antioxidant capacity and anti-apoptotic, promoting the survival and neurogenesis, normalized hippocampal neurotransmitters. Therefore, they may be considered as healthcare anti-aging products to decrease incidence of age related brain diseases such as; Alzheimer which may be due to their free radical scavenger affinity.
This study discover the possible role of linseed oil and coenzyme Q10 as compared with standard neuroprophylactic Ginkgo biloba extract to protect hippocampal cells against toxic effects induced by D-galactose where the dietary intake of natural substances may be beneficial in common aging troubles.
SIGNIFICANCE STATEMENT
This study discover the comparison between the most common neuroprotective herb and diet rich with linseed oil or CoQ10 on D-galactose induced metabolic syndrome and neuronal destruction, that can be beneficial for attention to medicinal herbs and nutritional supplements with ageing. This study will help the researcher to uncover the critical areas of the neuronal monoamines turn-over and subsequently apoptotic stimulation after aging combined with D-gal that many researchers were not able to explore. Thus, a new theory on neuronal apoptosis may be arrived at.
REFERENCES
- Kumar, A., A. Prakash and S. Dogra, 2011. Centella asiatica attenuates D-galactose-induced cognitive impairment, oxidative and mitochondrial dysfunction in mice. Int. J. Alzheimers Dis., Vol. 2011.
CrossRefDirect Link - Ho, S.C., J.H. Liu and R.Y. Wu, 2003. Establishment of the mimetic aging effect in mice caused by D-galactose. Biogerontology, 4: 15-18.
CrossRefDirect Link - Ames, B.N., M.K. Shigenaga and T.M. Hagen, 1993. Oxidants, antioxidants and the degenerative diseases of aging. Proc. Nat. Acad. Sci. USA, 90: 7915-7922.
CrossRefDirect Link - Lu, J.M., P.H. Lin, Q. Yao and C. Chen, 2010. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell. Mol. Med., 14: 840-860.
CrossRefDirect Link - Ullah, F., T. Ali, N. Ullah and M.O. Kim, 2015. Caffeine prevents D-galactose-induced cognitive deficits, oxidative stress, neuroinflammation and neurodegeneration in the adult rat brain. Neurochem. Int., 90: 114-124.
CrossRefDirect Link - Lee, K.Y., H.Y. Jung, D.Y. Yoo, W. Kim and J.W. Kim et al., 2017. Dendropanax morbifera Léveille extract ameliorates D-galactose-induced memory deficits by decreasing inflammatory responses in the hippocampus. Lab. Anim. Res., 33: 283-290.
CrossRefDirect Link - Zhu, J., X. Mu, J. Zeng, C. Xu and J. Liu et al., 2014. Ginsenoside Rg1 prevents cognitive impairment and hippocampus senescence in a rat model of D-galactose-induced aging. PloS One, Vol. 9, No. 6.
CrossRefDirect Link - Edeas, M., D. Attaf, A.S. Mailfert, M. Nasu and R. Joubet, 2010. Maillard reaction, mitochondria and oxidative stress: Potential role of antioxidants. Pathol. Biol., 58: 220-225.
CrossRefDirect Link - Abdel-Moneim, A.E., M.A. Dkhil and S. Al-Quraishy, 2011. The redox status in rats treated with flaxseed oil and lead-induced hepatotoxicity. Biol. Trace Element Res., 143: 457-467.
CrossRefDirect Link - Devarshi, P.P., N.M. Jangale, A.E. Ghule, S.L. Bodhankar and A.M. Harsulkar, 2013. Beneficial effects of flaxseed oil and fish oil diet are through modulation of different hepatic genes involved in lipid metabolism in streptozotocin-nicotinamide induced diabetic rats. Genes Nutr., 8: 329-342.
CrossRefDirect Link - Barcelo-Coblijn, G. and E.J. Murphy, 2009. Α-linolenic acid and its conversion to longer chain n-3 fatty acids: Benefits for human health and a role in maintaining tissue n-3 fatty acid levels. Prog. Lipid Res., 48: 355-374.
CrossRefDirect Link - Laredj, L.N., F. Licitra and H.M. Puccio, 2014. The molecular genetics of coenzyme Q biosynthesis in health and disease. Biochimie, 100: 78-87.
CrossRefDirect Link - Lopez-Lluch, G., J.C. Rodriguez-Aguilera, C. Santos-Ocana and P. Navas, 2010. Is coenzyme Q a key factor in aging? Mech. Ageing. Dev., 131: 225-235.
CrossRefDirect Link - Martin-Montalvo, A., Y. Sun, A. Diaz-Ruiz, A. Ali and V. Gutierrez et al., 2016. Cytochrome b 5 reductase and the control of lipid metabolism and healthspan. NPJ Aging Mech. Dis., Vol. 2, No. 1.
CrossRefDirect Link - Crane, F.L., 2001. Biochemical functions of coenzyme Q10. J. Am. Coll. Nutr., 20: 591-598.
CrossRefDirect Link - Blecharz-Klin, K., A. Piechal, I. Joniec, J. Pyrzanowska and E. Widy-Tyszkiewicz, 2009. Pharmacological and biochemical effects of Ginkgo biloba extract on learning, memory consolidation and motor activity in old rats. Acta Neurobiol. Exp., 69: 217-231.
Direct Link - Rajesha, J., K.N.C. Murthy, M.K. Kumar, B. Madhusudhan and G.A. Ravishankar, 2006. Antioxidant potentials of flaxseed by in vivo model. J. Agric. Food Chem., 54: 3794-3799.
CrossRefDirect Link - Emam, A.M., G.S. Georgy, O.G. Shaker, H.M. Fawzy and H.F. Zaki, 2016. Protective effects of alpha-lipoic acid and coenzyme Q10 on lipopolysaccharide-induced liver injury in rats. Der Pharmacia Lett., 8: 176-182.
Direct Link - Rahman, M.N.A. and A.A. Elsheikh, 2012. Study of the possible modulatory effect of resveratrol and coenzyme Q10 on MPTP-induced parkinsonism in mice. J. Am. Sci., 8: 958-967.
Direct Link - Erdinçler, D.S., A. Seven, F. Inci, T. Beǧer and G. Candan, 1997. Lipid peroxidation and antioxidant status in experimental animals: Effects of aging and hypercholesterolemic diet. Clin. Chim. Acta, 265: 77-84.
CrossRefDirect Link - Miranda, K.M., M.G. Espey and D.A. Wink, 2001. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide, 5: 62-71.
CrossRefDirect Link - Nishikimi, M., N.A. Rao and K. Yagi, 1972. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun., 46: 849-854.
CrossRefPubMedDirect Link - Koracevic, D., G. Koracevic, V. Djordjevic, S. Andrejevic and V. Cosic, 2001. Method for the measurement of antioxidant activity in human fluids. J. Clin. Pathol., 54: 356-361.
CrossRefPubMedDirect Link - Salah, A.S., O.A. Ahmed-Farid and M.S. El-Tarabany, 2019. Carcass yields, muscle amino acid and fatty acid profiles and antioxidant indices of broilers supplemented with synbiotic and/or organic acids. J. Anim. Physiol. Anim. Nutr., 103: 41-52.
CrossRefDirect Link - Ahmed-Farid, O.A.H., M. Nasr, R.F. Ahmed and R.M. Bakeer, 2017. Beneficial effects of Curcumin nano-emulsion on spermatogenesis and reproductive performance in male rats under protein deficient diet model: Enhancement of sperm motility, conservancy of testicular tissue integrity, cell energy and seminal plasma amino acids content. J. Biomed. Sci., Vol. 24, No. 1.
CrossRefDirect Link - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - Ali, E.H.A., O.A. Ahmed-Farid and A.A.E. Osman, 2017. Bone marrow-derived mesenchymal stem cells ameliorate sodium nitrite-induced hypoxic brain injury in a rat model. Neural Regen. Res., 12: 1990-1999.
CrossRefPubMedDirect Link - Bancroft, J., A. Stevens and D. Turner, 1996. Theory and Practice of Histological Techniques. 4th Edn., Churchill, Livingston, New York, London, San Francisco, Tokyo Pages: 740.
CrossRefDirect Link - Spiteller, G., 2010. Is lipid peroxidation of polyunsaturated acids the only source of free radicals that induce aging and age-related diseases? Rejuvenation Res., 13: 91-103.
CrossRefDirect Link - Kim, S.E., I.G. Ko, B.K. Kim, M.S. Shin and S. Cho et al., 2010. Treadmill exercise prevents aging-induced failure of memory through an increase in neurogenesis and suppression of apoptosis in rat hippocampus. Exp. Gerontol., 45: 357-365.
CrossRefDirect Link - Maurissen, J.P., B.R. Marable, A.K. Andrus and K.E. Stebbins, 2003. Factors affecting grip strength testing. Neurotoxicol. Teratol., 25: 543-553.
CrossRefDirect Link - Ashour, F.A., H. Abdel-Razek, G.S. Youssef, S.F. Ewida and M.M. Adel, 2016. Effect of exercise and/or melatonin on spatial learning and memory of D-galactose-treated rats. Menoufia Med. J., 29: 944-953.
CrossRefDirect Link - Pourmemar, E., A. Majdi, M. Haramshahi, M. Talebi, P. Karimi and S. Sadigh-Eteghad, 2017. Intranasal cerebrolysin attenuates learning and memory impairments in D-galactose-induced senescence in mice. Exp. Gerontol., 87: 16-22.
CrossRefDirect Link - Majdi, A., S. Sadigh-Eteghad, M. Talebi, F. Farajdokht, M. Erfani, J. Mahmoudi and A. Gjedde, 2018. Nicotine modulates cognitive function in D-galactose-induced senescence in mice. Front. Aging Neurosci., Vol. 10.
CrossRefDirect Link - Kumar, A., S. Dogra and A. Prakash, 2009. Effect of carvedilol on behavioral, mitochondrial dysfunction and oxidative damage against D-galactose induced senescence in mice. Naunyn-Schmiedeberg's Arch. Pharmacol., 380: 431-441.
CrossRefDirect Link - Haider, S., S. Saleem, T. Perveen, S. Tabassum and Z. Batool et al., 2014. Age-related learning and memory deficits in rats: role of altered brain neurotransmitters, acetylcholinesterase activity and changes in antioxidant defense system. Age, Vol. 36, No. 3.
CrossRefDirect Link - Evans, M.D., M. Dizdaroglu and M.S. Cooke, 2004. Oxidative DNA damage and disease: Induction, repair and significance. Mutat. Res./Rev. Mutat. Res., 567: 1-61.
CrossRefDirect Link - Brand, M.D., 2010. The sites and topology of mitochondrial superoxide production. Exp. Gerontol., 45: 466-472.
CrossRefDirect Link - Xu, L.Q., Y.L. Xie, S.H. Gui, X. Zhang and Z.Z. Mo et al., 2016. Polydatin attenuates D-galactose-induced liver and brain damage through its anti-oxidative, anti-inflammatory and anti-apoptotic effects in mice. Food Funct., 7: 4545-4555.
CrossRefDirect Link - Anand, K.V., M.S.M. Jaabir, P.A. Thomas and P. Geraldine, 2012. Protective role of chrysin against oxidative stress in D‐galactose‐induced aging in an experimental rat model. Geriatr. Gerontol. Int., 12: 741-750.
CrossRefDirect Link - Dkhar, P. and R. Sharma, 2010. Effect of dimethylsulphoxide and curcumin on protein carbonyls and reactive oxygen species of cerebral hemispheres of mice as a function of age. Int. J. Dev. Neurosci., 28: 351-357.
CrossRefDirect Link - Bekinschtein, P., M. Cammarota, C. Katche, L. Slipczuk and J.I. Rossato et al., 2008. BDNF is essential to promote persistence of long-term memory storage. Proc. Natl. Acad. Sci. USA., 105: 2711-2716.
CrossRefDirect Link - Woo, J.Y., W. Gu, K.A. Kim, S.E. Jang, M.J. Han and D.H. Kim, 2014. Lactobacillus pentosus var. plantarum C29 ameliorates memory impairment and inflammaging in a D-galactose-induced accelerated aging mouse model. Anaerobe, 27: 22-26.
CrossRefDirect Link - Erickson, K.I., R.S. Prakash, M.W. Voss, L. Chaddock and S. Heo et al., 2010. Brain-derived neurotrophic factor is associated with age-related decline in hippocampal volume. J. Neurosci., 30: 5368-5375.
CrossRefDirect Link - Kou, J., D. Zhu and Y. Yan, 2005. Neuroprotective effects of the aqueous extract of the Chinese medicine Danggui-Shaoyao-san on aged mice. J. Ethnopharmacol., 97: 313-318.
CrossRefDirect Link - Zhou, X.Q., Z.W. Yao, Y. Peng, S.S. Mao, D. Xu, X.F. Qin and R.J. Zhang, 2018. PQQ ameliorates D-galactose induced cognitive impairments by reducing glutamate neurotoxicity via the GSK-3β/Akt signaling pathway in mouse. Scient. Rep., Vol. 8, No. 1.
CrossRefDirect Link - Liang, L.P., S. Waldbaum, S. Rowley, T.T. Huang, B.J. Day and M. Patel, 2012. Mitochondrial oxidative stress and epilepsy in SOD2 deficient mice: attenuation by a lipophilic metalloporphyrin. Neurobiol. Dis., 45: 1068-1076.
CrossRefDirect Link - Wang, C.H., S.B. Wu, Y.T. Wu and Y.H. Wei, 2013. Oxidative stress response elicited by mitochondrial dysfunction: implication in the pathophysiology of aging. Exp. Biol. Med., 238: 450-460.
CrossRefDirect Link - Valavanidis, A., T. Vlachogianni and C. Fiotakis, 2009. 8-hydroxy-2'-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev., 27: 120-139.
CrossRefPubMedDirect Link - Kumar, A., A. Prakash and S. Dogra, 2010. Naringin alleviates cognitive impairment, mitochondrial dysfunction and oxidative stress induced by d-galactose in mice. Food Chem. Toxicol., 48: 626-632.
CrossRefPubMedDirect Link - Aydın, S., K. Yanar, P. Atukeren, E. Dalo and M.E. Sitar et al., 2012. Comparison of oxidative stress biomarkers in renal tissues of D-galactose induced, naturally aged and young rats. Biogerontology, 13: 251-260.
CrossRefDirect Link - Du, Z., Q. Yang, T. Zhou, L. Liu, S. Li, S. Chen and C. Gao, 2014. D-galactose-induced mitochondrial DNA oxidative damage in the auditory cortex of rats. Mol. Med. Rep., 10: 2861-2867.
CrossRefDirect Link - Yu, W., M. Bonnet, M. Farso, K. Ma and J.G. Chabot et al., 2014. The expression of Apoptosis Inducing Factor (AIF) is associated with aging-related cell death in the cortex but not in the hippocampus in the TgCRND8 mouse model of Alzheimer’s disease. BMC Neurosci., Vol. 15.
CrossRef - Scorrano, L. and S.J. Korsmeyer, 2003. Mechanisms of cytochrome C release by proapoptotic BCL-2 family members. Biochem. Biophys. Res. Commun., 304: 437-444.
CrossRefDirect Link - Kurokawa, M. and S. Kornbluth, 2009. Caspases and kinases in a death grip. Cell, 138: 838-854.
CrossRefPubMedDirect Link - Su, K.Y., C.Y. Yu, Y.P. Chen, K.F. Hua and Y.L.S. Chen, 2014. 3, 4-Dihydroxytoluene, a metabolite of rutin, inhibits inflammatory responses in lipopolysaccharide-activated macrophages by reducing the activation of NF-κB signaling. BMC Complement. Altern. Med., Vol. 14, No. 1.
CrossRefDirect Link - Shahroudi, M.J., S. Mehri and H. Hosseinzadeh, 2017. Anti-aging effect of Nigella sativa fixed oil on D-galactose-induced aging in mice. J. Pharmacopuncture, 20: 29-35.
CrossRefPubMedDirect Link - Freemantle, E., M. Vandal, J. Tremblay-Mercier, S. Tremblay and J.C. Blachère et al., 2006. Omega-3 fatty acids, energy substrates and brain function during aging. Prostaglandins, Leukotrienes Essential Fatty Acids, 75: 213-220.
CrossRefDirect Link - Joardar, A. and S. Das, 2007. Effect of fatty acids isolated from edible oils like mustard, linseed or coconut on astrocytes maturation. Cell. Mol. Neurobiol., 27: 973-983.
CrossRefDirect Link - Du Bois, T.M., C. Deng, W. Bell and X.F. Huang, 2006. Fatty acids differentially affect serotonin receptor and transporter binding in the rat brain. Neuroscience, 139: 1397-1403.
CrossRefDirect Link - Yehuda, S., S. Rabinovitz and D.I. Mostofsky, 2006. Nutritional deficiencies in learning and cognition. J. Pediatr. Gastroenterol. Nutr. Des., 43: S22-S25.
CrossRefDirect Link - Beal, M.F., D.R. Henshaw, B.G. Jenkins, B.R. Rosen and J.B. Schulz, 1994. Coenzyme Q10 and nicotinamide block striatal lesions produced by the mitochondrial toxin malonate. Ann. Neurol., 36: 882-888.
CrossRefDirect Link - Schiefer, J., G.B. Landwehrmeyer, H.G. Lüesse, A. Sprünken and C. Puls et al., 2002. Riluzole prolongs survival time and alters nuclear inclusion formation in a transgenic mouse model of Huntington's disease. Movement Disord., 17: 748-757.
CrossRefDirect Link - Gao, H.L., X.J. Yu, J. Qi, Q.Y. Yi and W.H. Jing et al., 2016. Oral CoQ10 attenuates high salt-induced hypertension by restoring neurotransmitters and cytokines in the hypothalamic paraventricular nucleus. Scient. Rep., Vol. 6.
CrossRefDirect Link - Swomley, A.M., S. Förster, J.T. Keeney, J. Triplett, Z. Zhang, R. Sultana and D.A. Butterfield, 2014. Abeta, oxidative stress in Alzheimer disease: Evidence based on proteomics studies. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis., 1842: 1248-1257.
CrossRefDirect Link - Tian, G., J. Sawashita, H. Kubo, S.Y. Nishio and S. Hashimoto et al., 2014. Ubiquinol-10 supplementation activates mitochondria functions to decelerate senescence in senescence-accelerated mice. Antioxidants Redox Signal., 20: 2606-2620.
CrossRefDirect Link - Hargreaves, I.P., 2014. Coenzyme Q10 as a therapy for mitochondrial disease. Int. J. Biochem. Cell Biol., 49: 105-111.
CrossRefDirect Link - Fischer, A., S. Onur, P. Niklowitz, T. Menke, M. Laudes, G. Rimbach and F. Döring, 2016. Coenzyme Q10 status as a determinant of muscular strength in two independent cohorts. PloS One, Vol. 11, No. 12.
CrossRefDirect Link