
Ayodele O. Kolawole
Department of Biochemistry, Federal University ofTechnology, Akure, Nigeria
Joshua O. Ajele
Department of Biochemistry, Federal University ofTechnology, Akure, Nigeria
Journal of Plant Sciences
Year: 2007 | Volume: 2 | Issue: 4 | Page No.: 425-432







ABSTRACT
The effect of 4-days malting on the thermo-activation thermodynamic data of β-amylase from African finger millet (Eluesine coracana) seed was determined and compared with that of the β-amylase purified from unmalted seed. The results showed that the relative importance of enthalpic (ΔH!) and entropic activation between malted and unmalted was different. The activation energy for catalysis of unmalted Eluesine coracana β-amylase was 2.00 kcal mol1. The free energy change (ΔG!), enthalpy change (ΔH!) and entropy change (ΔS!) for activation at 50°C (optimum temperature) were 15.60 kcal mol1, 1.36 kcal mol1 and -45.60 e.u. (entropy unit), respectively. The activation energy for catalysis of malted Eluesine coracana β-amylase was 7.539 kcal mol1. The free energy change (ΔG!), enthalpy change (ΔH!) and entropy change (ΔS!) for activation at 50°C were 15.60 kcal mol1, 6.90 kcal mol1 and -24.96 e.u., respectively. The purified β-amylase from malted seed at 90°C had an half life of 5.87 min with free energy of inactivation of 22.928 kcal mol1, enthalpy 14.640 kcal mol1 and entropy -22.83 e.u. The results indicated that malted seed β-amylase reaction is enthalpy driven and unmalted seed β-amylase reaction is entropy driven to activation complex. The thermoactiavtion data both for malted and unmalted shows that the reaction is bisubstrate with a unimolecular rate determining step. The thermodynamic data of inactivation suggest that the enzyme is less stable at higher temperatures.
PDF Abstract XML References
How to cite this article
Ayodele O. Kolawole and Joshua O. Ajele, 2007. Comparative Studies on Thermodynamic Activation Data Between Malted and unmalted African Finger Millet Seed(Eluesine coracana) β-Amylase. Journal of Plant Sciences, 2: 425-432.
DOI: 10.3923/jps.2007.425.432
URL: https://scialert.net/abstract/?doi=jps.2007.425.432
DOI: 10.3923/jps.2007.425.432
URL: https://scialert.net/abstract/?doi=jps.2007.425.432
INTRODUCTION
β-amylase (E.C. 3.2.1.2) plays an important role in the biogeochemical cycle of carbon and it also has a wider application in the biotechnological-based pharmaceutical, food and beverage industries in the conversion of starch into maltose solution (Mohapatra et al., 1998). β-amylase hydrolyses α-1,4-glucosidic linkages of the starch-type substrate in an exo-fashion from the non-reducing end to β-maltose (Visko-Neilsen et al., 1997). High value is placed on extreme thermostability and thermo activity of β-amylase used in the bioprocessing of starch because of the high temperature operating condition (Saha and Zeikus, 1989).
At higher temperature, thermostability of proteins generally and of enzymes in particular is as a result of resistance to formation of incorrect conformations due to processes such as hydrolysis of peptide bond, resistance to destruction of amino acids and aggregation (Tomb, 1985; Ahern and Manning, 1992). These reactions have pH and temperature dependencies as was reported for endo-1,4-glucanase in Thermomonospora curvata (Stutzenberger and Lupo, 1985) and for some amylases (Briggs et al., 1981).
Thermodynamic data of thermo-activation on enzyme-catalyzed reactions play an important role in the prediction of the extent and position of equilibrium for any process in which these reactions occur. Thermodynamic information of thermo-activation is also needed in biotechnology when one needs to optimize product yields and to calculate the energy requirements of a given reaction (Goldberg et al., 1993).
Recently, study showed that African finger millet has good malting and brewing characteristics (Nzelibe and Agbojo, 2005) and exhibited maximal activity at day 4 of germination. The essence of malting is for the mobilization of the enzymes of the grain especially β-amylase which is a diastic power indicator (Briggs et al., 1981). Reporting thermal thermodynamic data for β-amylase has not been customary. The significance of this parameter and its relationship to the β-amylase catalysis has not yet been analyzed in detail. The present study is, therefore, designed to examine the influence of malting on thermal activation thermodynamic data (enthalpy, entropy and free energy of activation) of the purified β-amylase from Eluesine coracana at the temperatures where the enzyme appeared to operate actively.
MATERIALS AND METHODS
Materials
The study was conducted at the Department of Biochemistry, Obafemi Awolowo University, Ile-Ife, Nigeria in the last quarter of year 2006. 3,5-dinitrosalicylic acid, sodium potassium tartarate and potato soluble starch were products of BDH Chemical Limited, Poole, England. Epoxyl activated Sepharose 6B was from Pharmacia Fine Chemicals, Uppsala, Sweden. α-Cyclodextrin was from Sigma Chemical Company Limited. St Loius, Mo., USA. All other chemicals and reagents used were of analytical grade. African finger millet seed (Eluesine coracana) was obtained from a local market in Jos, Nigeria.
Methods
Preparation of α-Cyclodextrin Sepharose 6B (Affinity gel)
α-Cyclodextrin was immobilized on epoxyl activated Sepharose 6B according to the method of Vretbald (1974). Epoxyl activated Sepharose 6B (8.0 g) was swollen in distilled water and then transferred to 0.1 M NaOH. After removal of excess liquid by suction, the gel was reacted with 600 mg of α-cyclodextrin in 24 mL of 0.1 M NaOH with shaking at 45±3°C for 24 h. The immobilized α-cyclodextrin- was washed as described and then used.
Enzyme Extraction
Ground seed (100 g) was mixed and stirred in cold 300 mL 50 mM sodium acetate buffer pH 4.5 for 24 h. The homogenate was filtered using cheesecloth. The filtrate was centrifuged at 14000 g for 10 min with Automatic Superspeed centrifuge at 4°C. The supernatant was filtered through a loose plug of glass wool to remove the floating lipids. The filtrate served as the crude extract. Another 100 g of the seed was soaked in distilled water for 24 h. Excess water was drained and allowed to malt for 4 days on moist paper and crude extract was got through the same process as described above.
Purification
The method of purification adopted was that of Totsuka and Fukazawa (1993) was adopted. Crude extract (5.0 mL) was applied to a column (1.0x10 cm) packed with the affinity gel that had previously been equilibrated with 50 mM acetate buffer pH 4.5 containing 1.0 M ammonium sulphate. The concentrated extracts were earlier mixed with 1.0 M ammonium sulphate. The unadsorbed proteins were eluted. Fractions of 5.0 mL were collected at a flow rate of 10 mL h-1. The bound proteins were eluted with the same eluting buffer containing no ammonium sulphate. The fractions containing β-amylase activity were pooled and stored at -5°C. Unless otherwise stated all purifications were carried out at 4°C in the cold laboratory.
Enzyme Assay
β-Amylase activity was measured as described by Oboh and Ajele (1997) using 3,5-dinitrosalicylic acid (DNSA) colour reagent. The appropriately diluted enzyme solution (0.5 mL) of protein concentration of 0.07 mg mL-1 was added to 0.5 mL of 1% (w/v) soluble starch dissolved in the appropriate buffer. Control consisted of 0.5 mL buffer plus 0.5 mL soluble starch solution. Reaction tubes were incubated at the appropriate temperature for 3 min. DNSA reagent (1.0 mL) was added to each tube and the tube was placed in a boiling water bath for 5 min. The tubes were cooled and 10 mL of distilled water was added and mixed and the absorbance was taken at 540 nm. The unit of β-amylase activity was defined as that amount of enzyme, which released one micromole of maltose from starch per minute at 25°C and pH 6.0. Protein concentration was determined by the method of Lowry et al. (1951) using bovine serum albumin as standard.
Activation Energy for Catalysis (Ea)
African finger millet seed β-Amylase from 4 days malted seeds and unmalted seeds was assayed as earlier described (Kolawole et al., 2006) at various temperatures ranging from 5 to 50°C. The ln Kcat values were plotted against the Arrhenius temperature. Activation energy of catalysis of β-amylase was calculated from the slope of the plot and thermodynamic data were calculated by rearranging Erying’s absolute equation derived from the transition state theory as described by Eyring and Stearn (1939).
Activation Energy for Denaturation
The first order rate constants for denaturation (Kd) of the enzyme at different temperatures (50, 60, 70, 80 and 90°C) were determined. The first order rate constant of malted seed β-amylase were plotted as described by Sizer (1943) and Alrokayan (2002) and thermodynamic data were calculated by rearranging Erying’s absolute equation derived from the transition state theory as described by Eyring and Stearn (1939). The first order rate constants were used to calculate the activation energy of denaturation (Ea = Slope X R). R is the gas constant (1.9872 cal K-1 mol-1). The apparent activation energy of denaturation was calculated.
The half-life was determined from the relationship:
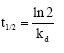 |
where kd is the first order deactivation rate constant that can be derived from:
where vo is the initial enzyme activity while v is the enzyme activity time at of incubation and -kd can be obtained from the plot of:
RESULTS
The Arrhenius plot is linear for unmalted African finger millet seed β-amylase at physiological temperature (5-50°C) and, malted African finger millet seed β-amylase displayed curved Arrhenius plot at physiological temperatures. The activation energy of catalysis for β-amylase from unmalted and malted African finger millet seed were 2.00 and 7.54 kcal mol-1, respectively.
| Table 1: | Thermodynamic parameters for thermal activation of β-amylase from malted and unmalted African finger millet seed |
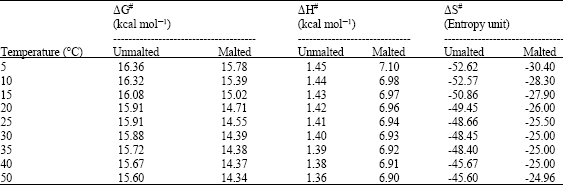 | |
| ΔEa of activation for β-amylase from malted seed was 2.00 kcal mol-1 and ΔEa of activation for β-amylase from unmalted seed was 7.539 kcal mol-1 | |
| Table 2: | Thermodynamic parameters for thermal inactivation of malted African finger millet seed β-amylase |
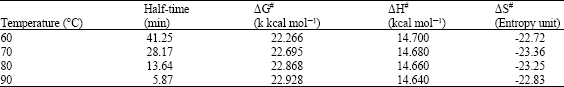 | |
| ΔEa (activation for denaturation) = 15.362 kcal mol-1 | |
The order of thermal activation appears to be first order (Figure not shown). The activation enthalpy (ΔH#) and entropy (ΔS#) were 1.40 kcal mol-1 and -48.45 e.u. for the β-amylase from unmalted and 6.93 kcal mol-1 and -25.0 e.u. for the β-amylase from malted at 30°C, respectively (Table 1 and 2). There was a major decrease in the enthalpy and entropy as the temperature increased both in malted and unmalted seed β-amylase.
The first order rate constant from Table 2 was used to calculate the activation energy of denaturation (Ea = - slope X R). The apparent activation energy was estimated to be 15.362 kcal mol-1. As shown in Table 2, the β-amylase from day 4 malted seed thermodynamic data for inactivation showed a positive ΔG#, ΔH# but negative ΔS#. The half life of the enzyme at 60°C and 90°C were 41.25 min and 5.87 min, respectively. The purified β-amylase from malted seed at 90°C has free energy of inactivation of 22.928 kcal mol-1, enthalpy 14.640 kcal mol-1 and entropy -22.83 e.u. The result of the thermodynamic data of inactivation showed that with increase in temperature there was a reduction in half-life, increase free energy of deactivation and reduction in the enthalpy of deactivation.
DISCUSSION
Analysis and calculation of activation thermodynamic parameters (free energy of activation, enthalpy and entropy of activation) describing the thermodynamic differences between the ground state and the transition state provide some vital information to the nature of the transition state, the energy requirements to the transition state and the mechanism involved in the reaction (Jencks, 1987; Piszkiewicz, 1977; Goldberg et al., 1993). Higher ΔG# values reflect decreased activity; high ΔH# values reflect the increased thermo-dependence and lower entropic contributions possibly reflect reduced conformational changes between the ground state ES complex and the activated transition state (D’Amico et al., 2003). The result of this study has shown that the free energy of activation of β-amylase in 4 days malted finger millet seed was lower than that of the unmalted finger millet seed β-amylase (Table 1). Free energy of activation is related to reaction rate (Piszkiewicz, 1977). The low free energy of activation data is quite instructive. It shows that the rate of reaction catalyzed by β-amylase is faster in malted than in unmalted seed under the activation temperature range. The probable reason for malting by maltster had been to mobilize diastic enzymes and better expression of the enzymes (Briggs et al., 1981). These thermodynamic data may provide another probable reason for malting. It is to increase the saccharification rate. The low free energy of activation of malted over unmalted offers low cost saving in terms of low energy requirements. The thermodynamic data might indicate the probable reason why traditional maltster prefers to use malted grain as an adjunct. Malted African millet has been used in the traditional non-alcoholic beverage, Togwa, in East Africa (Kitabatake et al., 2003) and Togwa contains many kinds and a high quantity of minerals derived from finger millet malt (Oi and Kitabatake, 2003).
The distinction between different activation parameters (free energy of activation, enthalpy and entropy of activation) may reflect different conformation changes of β-amylase in the malted and unmalted seeds towards activation. From the results, the rate of enhancement of the enzyme reaction in the malted is enthapic driven while the reaction in the unmalted seed is entropic in its origin. The major decrease in entropy with increased temperature indicates a drastic change in the conformation of the enzyme and this appears to favour catalysis. The enthalpies of activation, not entropy of activation tend to govern the variation of spontaneous reactions (Wolfenden et al., 1999). The high enthalpy of activation in malted seed shows the enzymatic reaction is more spontaneous, compared, to enzymic reaction in unmalted seed β-amylase governed by entropy of activation. It has earlier been proposed that sequence changes outside the active site can alter the enthalpy and entropy of conformational changes of importance in binding and catalysis; these effects should play a critical role in evolutionary adaptation of kcat and Km and in establishing the responses of these kinetic parameters to acute temperature changes (Fields and Somero, 1998).
Spontaneous enzymatic reaction is second order, probably bimolecular with a unimolecular rate determining step (Leininger and Westley, 1968). This might indicate that β-amylase action in both malted and unmalted is bi-substrate and water molecule serving as a second substrate. The order of addition of the two substrates is still not understood. In the protein enzyme field, curved Arrhenius plots can often be ascribed to temperature-induced changes in the conformation of the catalyst (Dixon and Webbs, 1979). The curved Arrhenius plot derived for the enzymic reaction in the case of malted seed β-amylase is an indication that the change in conformation of β-amylase that favour catalysis was temperature induced (Fig. 1) while unmalted seed β-amylase remains temperature uninduced, hence possibly adapted to cold induction (Fig. 2). The unmalted β-amylase might probably be entropy driven as shown by high negative entropy change (Table 1). This opens a new way into further investigation on the structural techniques of X-ray crystallography and NMR to unravel the molecular basis of this phenomenon.
The decrease of ΔH# is the major adaptive characteristic of cold-adapted enzymes is function effectively at low temperature and is achieved by decreasing the number of enthalpy driven interactions that have to be broken during formation of the transition state (D’Amico et al., 2002).
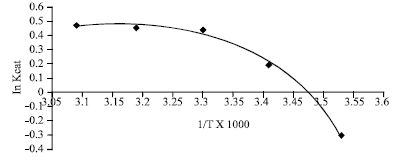 | |
| Fig. 1: | Arrhenius plot for the determination of activation energy (Ea) for catalysis of 4 day malted African finger millet seed β-amylase |
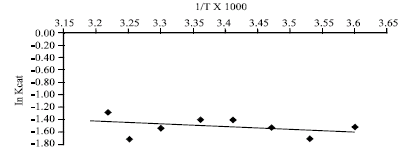 | |
| Fig. 2: | Arrhenius plot for the determination of activation energy (Ea) for catalysis of β-amylase from unmalted African finger millet seed |
This renders the reaction less temperature dependent (Lonhienne et al., 2001). The present thermodynamic data of activation might give some further information on the possible roles of the beta amylase in unmalted cereal. It shows that in situ β-amylase in unmalted seed is adapted to convert soluble starch at low temperature, albeit, at slower rate compared to malted seed. It therefore shows that in situ β-amylase is a regulatory enzyme especially under cold condition as earlier posited by Kaplan and Guy (2004). That means the beta amylase is involved in maltose accumulation without an increase in beta amylase activity. A study by Kaplan and Guy (2004) demonstrated that the induction of β-amylase under abiotic stress was to protect proteins, membranes and photosynthetic electrons transports in response to acute temperature stress.
Negative entropy of activation suggests the need for the reactants (starch and water) to properly orient themselves in the transition state (Piszkiewicz, 1977). The high entropy of activation in the unmalted seed β-amylase might contribute to the stability of the folded conformation and as a result, the region of the enzyme containing the active site should be less stable. This might likely explain the activity-stability trade off in cold adapted enzymes (D’Amico et al., 2002).This high entropy ordering as posited by the high entropy of activation might be involved in regulatory role of the enzyme to unnecessary maltose accumulation. The β-amylase from malted seed is more ordered and require less entropy of activation to imposing its orientation and steric requirements on its substrates. The result has indicated that during malting, there was favourable conformational change in the finger millet seed β-amylase. The conformational change might be optimized at day 4 of malting. The negative entropy of activation has, also, shown that the reaction is bisubstrate (Piszkiewicz, 1977).
Maximizing the output of industrial enzyme, like finger millet seed β-amylase require less balance of the effect of temperature on enzyme stability and activation. Thermodynamic data for thermal activation provides a basis for this. The thermodynamic data for inactivation for the malted seed has shown the rate of denaturation is very slow at lower temperature but increased rapidly at higher temperature. The thermal denaturation brought about the increase in the enthalpy and entropy of activation of the malted finger millet seed β-amylase. Thermal denaturation has been proposed to proceed in two steps (N � U → 1) where N is the native enzyme, U is the unfolded enzyme that can reversibly be restored back to its active form upon cooling and I is the inactivated enzyme that cannot be reversibly refolded upon cooling. The thermal inactivation of β-amylase is accompanied by the disruption of the non-covalent bond with concomitant increase in the enthalpy and opening of the enzyme structure by an increase in the disruption or entropy of activation. This indicates a drastic change in the conformation as the temperature increases. The overall thermodynamic data of inactivation suggest that the enzyme is less stable at higher temperatures. Increasing the stability of the enzyme at higher temperature merit further investigation, maybe, through chemical modification studies.
REFERENCES
- Alrokayan, S., 2002. Purification and characterization of adenosine deaminase from camel skeletal muscle. Int. J. Biochem. Cell Biol., 34: 1608-1618.
Direct Link - D'Amico, S., P. Claverie, T. Collins, D. Georlette and E. Gratia et al., 2002. Molecular basis of cold adaptation. Phil. Trans. R. Soc. Lond. B, Biol. Sci., 357: 917-925.
Direct Link - D'Amico, S., C. Gerday and G. Feller, 2003. Temperature adaptation of proteins: Engineering mesophilic-like activity and stability in a cold-adapted α-amylase. J. Mol. Biol., 332: 981-988.
Direct Link - Field, P.A.P. and G.N. Somero, 1998. Hot spots in cold adaptation: Localized increases in conformational flexibility in lactate dehydrogenase A orthologs of Antarctic notothenioid fishes. Proc. Nat. Acad. Sci., 95: 11476-11481.
Direct Link - Goldberg, R.N., Y.B. Tewari, D. Bell, K. Fazio and E. Anderson, 1993. Thermodynamics of enzyme-catalyzed reactions: Part 1. Oxidoreductases. J. Phys. Chem. Ref. Data, 22: 515-582.
Direct Link - Kaplan, F. and C.L. Guy, 2004. β-amylase induction and the protective role of maltose during temperature shock. Plant Physiol., 135: 1674-1684.
Direct Link - Kitabatake, N., D.M. Gimbi and Y. Oi, 2003. Traditional non-alcoholic beverage, Togwa, in East Africa, produced from maize flour and germinated finger millet. Int. J. Food Sci. Nutr., 54: 447-455.
Direct Link - Kolawole, A.O., D.M. Sanni and J.O. Ajele, 2006. Catalytic and thermodynamic properties of digitaria exilis β-amylase. J. Plant Sci., 1: 273-285.
CrossRefDirect Link - Lonhienne, T., E. Baise, G. Feller, V. Bouriotis and C. Gerday, 2001. Enzyme activity determination on macromolecular substrates by isothermal titration calorimetry: Application to mesophilic and psychrophilic chitinases. Biochim. Biophys. Acta, 1545: 349-356.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mohapatra, M.A., U.C. Banerjee and M. Bapuji, 1998. Characterization of a fungal amylase from Mucor sp. associated with the marine sponge Spirastrella sp. J. Biotechnol., 60: 113-117.
CrossRefDirect Link - Nzelibe, H.C. and D.A. Agbojo, 2005. Malting characteristics of different varieties of finger millet (Eluesine coracana) compared with Fonio millet (Digitaria exilis). Ife J. Sci., 7: 183-187.
Direct Link - Oi, Y. and N. Kitabatake, 2003. Chemical composition of an East African traditional beverage, Togwa. J. Agric. Food Chem., 51: 7024-7028.
Direct Link - Wolfenden, R., M. Snidder, C. Ridgway and B. Miller, 1999. The temperature dependence of enzyme rate enhancement. J. Am. Chem. Soc., 121: 7419-7420.
Direct Link