
Veena Sharma
Department of Bioscience and Biotechnology, Banasthali University, Banasthali, M.A.I. Jaipur, India
Leena Kansal
Department of Bioscience and Biotechnology, Banasthali University, Banasthali, M.A.I. Jaipur, India
Arti Sharma
Department of Bioscience and Biotechnology, Banasthali University, Banasthali, M.A.I. Jaipur, India
Shweta Lodi
Department of Bioscience and Biotechnology, Banasthali University, Banasthali, M.A.I. Jaipur, India
S.H. Sharma
Department of Bioscience and Biotechnology, Banasthali University, Banasthali, M.A.I. Jaipur, India
Journal of Pharmacy and Allied Health Sciences
Year: 2011 | Volume: 1 | Issue: 1 | Page No.: 16-29







ABSTRACT
To study the efficacy of Coriandrum sativum as an antioxidant in lead intoxicated mice. Oxidative stress was induced in mice by a daily dose of lead nitrate (40 mg kg-1 body weight by oral gavage) for seven days. From day eight, experimental animals received an oral dose of coriander extracts (aqueous extract -300 and 600 mg kg-1 body weight; ethanolic extract -250 and 500 mg kg-1 body weight) along with lead nitrate daily for 40 days. The coriander supplementation to intoxicated mice has protected the weights of experimental animals as compared to lead nitrate exposed animals. Ingestion of Pb (NO3)2 was significantly decreased RBC count, WBC count, Hb level and serum total protein contents in the lead nitrate treated mice. But, serum alanine transaminase, aspartate aminotransferase, creatinine and cholesterol level were significantly increased after implication of this metal. However, oral administration of Coriandrum sativum to lead treated mice led to marked improvement in both hematological and serum biochemical changes. A decrease in viability of neutrophiles, phagocytic index, immunoglobulin level and plaque count were the salient features observed in lead exposed animals. Oral administration of coriander extracts to Pb (NO3)2 treated groups attenuated the deranged parameters to some extent. The study reveals that treatment with coriander significantly protect against lead induced toxicity and warrants the isolation and identification of active compounds responsible for its antioxidant effect.
PDF Abstract XML References Citation
Received: April 04, 2011;
Accepted: April 05, 2011;
Published: August 29, 2011
How to cite this article
Veena Sharma, Leena Kansal, Arti Sharma, Shweta Lodi and S.H. Sharma, 2011. Ameliorating Effect of Coriandrum sativum Extracts on Hematological and Immunological Variables in an Animal Model of Lead Intoxication. Journal of Pharmacy and Allied Health Sciences, 1: 16-29.
DOI: 10.3923/jpahs.2011.16.29
URL: https://scialert.net/abstract/?doi=jpahs.2011.16.29
DOI: 10.3923/jpahs.2011.16.29
URL: https://scialert.net/abstract/?doi=jpahs.2011.16.29
INTRODUCTION
The environmental contamination by lead generated from human activities has become an evident problem during the last decades (Anaes, 2003). Lead can penetrate to the human or animal organisms by inhalation, ingestion and by skin (El-Feki et al., 2000). After absorption into the blood, 95% of lead is bound to erythrocytes and the remaining 5 percentage stay in plasma to be carried to other tissues. Erythrocytes have a high affinity for lead and contain the majority of the lead found in the blood stream which makes them more vulnerable to oxidative damage than many other cells. Moreover, erythrocytes can spread lead to different organs of the body (Sivaprasad et al., 2003).
Lead exposure is known to provoke weight reduction (El-Feki et al., 2000). In consequence, the reduction in body weight could be as a result of lead intoxication, where losing the animal appetite is the principal cause (Cezard and Haguenoer, 1992). However, the circulatory system, liver, kidneys, gonads and bones are the target organs for the accumulation of lead and its derivatives (Gerber et al., 1978).
A number of studies have documented the potential toxic effects of lead on the immune system (Cohen et al., 1994). Lead can suppress immune functions resulting in increased incidence and severity of infectious diseases (Kishikawa et al., 1997).
A limited amount of data suggest that the biochemical and molecular mechanisms of Pb toxicity involve the induction of oxidative stress in target cells, partly via the activation of Reactive Oxygen Species (ROS), followed by DNA damage and apoptosis (Fracasso et al., 2002). Nevertheless, indirect support for the involvement of reactive oxygen radicals in Pb toxicity has come from studies demonstrating beneficial effects of antioxidants on Pb-induced toxicity in various tissues, including brain, liver, kidney, sperm and blood cells (Patra et al., 2001).
A considerable body of research has been accumulated on chemical therapy of lead poisoning (Varnai et al., 2004). There is, however, a paucity of data about treatment of lead intoxication by natural products (Ashour, 2002).
Coriandrum sativum (Common name: Coriander), belonging to family Umbelliferae, is a herb that is widely cultivated in India and is recognized for its carminative and cooling properties (Sairam, 1998). Both the leaves and seeds of the plant are used for medicinal purpose. In the field of alternative medicine, coriander has immense value for the treatment of abdominal problems, especially stomach ulcers. The hypotensive (Burdock and Carabin, 2008) and hypoglycaemic (Eidi et al., 2009) properties of coriander have been documented. The ability of coriander to increase levels o f antioxidant enzymes and to manipulate lipid metabolism also have been reported by Chithra and Leelamma (2000). Coriander seeds contain moisture (6.3%), protein (11-17%), volatile oil (0.3%), nonvolatile ether extract (22%), ether extract (19.6%), crude fiber (31.5%), carbohydrates (24%), ash (5.3%), calcium (0.08%), phosphorus (0.44%), sodium (0.02%), potassium (1.2%), Vitamin B1 (0.26 mg 100 g), Vitamin B2 (0.23 mg 100 mg), niacin (3.2 mg 100 g), Vitamin C (ascorbic acid; 12 mg 100 g), Vitamin A (175 IU 100 g), flavonoid glycosides (26%) (Budvari, 1996) and essential oil linalool (60-80%), alpha-pinene (0.2-8.5%), gamma-terpinene (1-8%), geranylacetate (0.1-4.7%), camphor (1.4%) and geraniol (1.2-4.6%) (Burdock and Carabin, 2008). In Ayurvedic literature, the regular use of the decotation of the seeds of coriander is considered to be effective in lowering blood lipid levels (Sairam, 1998). It has also been speculated that Chinese parsley may enhance the excretion of heavy metals in the urine of patients with various infections and augments the efficacy of antibiotics (Omura et al., 1996).
The objective of this study, therefore, is to evaluate the protective role of coriander supplemented against lead toxicity in male mice.
MATERIALS AND METHODS
Chemicals: Lead nitrate was purchased from Central Drug House (India). All other chemicals used in the study were of analytical reagent and obtained from Sisco Research Laboratories (India), Qualigens (India/ Germany), SD fine chemicals (India), HIMEDIA (India) and Central Drug House (India).
Animals: Male Swiss albino mice weighing approximately 15-30 g (2 to 2.5 months) were obtained from Haryana Agricultural University, Hissar, India for experimental purpose. The Animal Ethics Committee of Banasthali University, Banasthali, India has approved the experimental protocol. All experiments were conducted on adult male albino mice (Mus musculus L.) weighing 25-35 g (3-4 month old). They were housed in polypropylene cages in an air conditioned room with temperature maintained at 25±3°C, relative humidity of 50±5% and 12 h alternating light and dark cycles. The mice were provided with a nutritionally adequate chow diet (Hindustan lever Limited, India) and drinking water ad libitum throughout the study.
Experimental plant material: The plant Coriandrum sativum (seeds) was collected from Krishi Vigyan Kendra, Banasthali University, Rajasthan, India and was identified as a RCR 435 variety.
Preparation of aqueous extract of Coriandrum sativum: Dried coriander seeds were ground to a fine powder, of which 100 g were added to 500 mL distilled water. After 24 h maceration was done at room temperature (37°C), the mixture was then heated for 30 min in the water bath at 65°C. The extract was filtered, concentrated by heating over the water bath (65°C) and dried under vacuum (Gray and Flatt, 1999) with the yield of 5.9% (w/w). The extract was stored at 4°C and used to treat animals as needed.
Preparation of alcoholic (ethanolic) extraction of Coriandrum sativum: The dried and powered seeds (200 g) were extracted successively with ethanol (800 mL) in a soxhlet extractor for 48 h at 60°C. After extraction, the solvent was evaporated to dryness at 50-55°C by using a rotary evaporator and the extract left behind (yield was 9.8%) was stored at 4°C. It was dissolved in distilled water whenever needed for experiments.
Procedure and design of experiments: In the present study 144 adult male Swiss albino mice (Mus musculus L.) weighing 25-35 g (3 to 4 months old) were used for hematological, biochemical and immunological studies.
| • | For hematological and biochemical studies: 72 mice were divided into 6 groups of 12 mice each |
| • | For immunological parameters: 72 mice were divided into 6 groups of 12 mice each |
| • | For immunological studies, all mice were antigenically challenged twice with sheep RBC, first challenge was on day 14 and second was on 20 day of the experiment |
The groups for each parameter treated once, daily by oral gavage were as follows:
| Group I | : | Control (Normal, Untreated), received distilled water without any treatment |
| Group II | : | Lead nitrate treated group, received freshly dissolved Pb (NO3)2 in 1 mL distilled water at a dose of 40 mg kg-1 body weight/day for 40 days |
| Group III | : | After exposure to lead nitrate (40 mg kg-1 body weight/day for 7 days) the animals received aqueous coriander extract at a dose of 300 mg kg-1 body weight/day and lead nitrate simultaneously for 33 days |
| Group IV | : | After exposure to lead nitrate (40 mg kg-1 body weight/day for 7 days) the animals received aqueous coriander extract at a dose of 600 mg kg-1 body weight/day and lead nitrate simultaneously for 33 days |
| Group V | : | After exposure to lead nitrate (40 mg kg-1 body weight/day for 7 days) the animals received ethanolic coriander extract at a dose of 250 mg kg-1 body weight/day and lead nitrate simultaneously for 33 days |
| Group VI | : | After exposure to lead nitrate (40 mg kg-1 body weight/day for 7 days) the animals received ethanolic coriander extract at a dose of 500 mg kg-1 body weight/day and lead nitrate simultaneously for 33 days |
The dose for lead nitrate was decided on the basis of experiments conducted in the laboratory and the concentration of lead nitrate used in the experiment was 1/56 of LD50 (Plastunov and Zub, 2008). The plant doses were selected on the basis of experiments conducted in our own laboratory and on the basis of earlier published reports, suggesting that Coriandrum sativum is having prophylactic efficacy (Sushruta et al., 2006). After the administration of the last dose, the animals were given rest overnight and then on the next day, they were sacrificed under light chloroform anesthesia. Blood was collected by heart puncture in eppendroff tubes rinsed with and without anticoagulant for hematological, serum biochemical and for immunological parameters. The spleen was excised and used for PFC assay.
Body weight: The body weights of animals recorded at the end of the experiment was nearest to gram. The % change in body weight of animals in different group was calculated by using the following formula:
Hematological variables: Total white blood cell counts, red blood cell count and hemoglobin content of each sample were determined by using standard laboratory procedures.
Hematochemical variables: The activities of Serum Glutamate Pyruvate Transaminase (SGPT) and Serum Glutamate Oxaloacetate Transaminase (SGOT) were assessed by the method of Reitman and Frankel (1957). Total protein content was determined by using Bovine serum albumin as a standard by the method of Lowery et al. (1951). The cholesterol level was determined by using Cholesterol as a standard by the method of Zak (1977). The concentration of creatinine in serum was determined by using QuantiChromTM Creatinine Assay Kit (DICT-500).
Immunological assays
Determination of antibody titer level (ELISA): The concentration of antibody in serum was determined by using GeNeiTM Antibody Capture ELISA Kit (Cat No. KT51).
Plaque forming cells (PFC) assay: PFC assay was done according to the method described by Jerne and Nordin (1963).
Preparation of a spleen cell suspension: The spleen from each animal was collected and all fat and extraneous tissue was removed. Spleen cell suspension from each spleen was prepared by grinding the spleen in 15 mL Balanced Salt Solution (BSS). The suspension was centrifuged at 500xg for 5 min and supernatant was decanted. Red blood cells were lysed by resuspending the pellet in 3 mL of ammonium tris chloride buffer (0.144 M NH4Cl, 0.017 M tris, pH 7.2) and incubated at 37°C for 3 min. The cell suspension was again washed with 15 mL BSS, centrifuged as above, and pellet was resuspended in 5 mL BSS.
Agar plate preparation: Agar (0.7 g) was combined with 50 mL HBSS at 80°C and gently stirred until completely dissolved. This solution (2 mL) was transferred to each petri dish with a warmed 5 mL pipette and allowed to solidify. These plates were stored at 4°C.
PFC assay and enumeration: Spleen suspension (250 μL) was combined with 0.15 mL of 20% SRBC and 0.80 mL of 0.7% agar solution (45°C), then poured into the previously prepared petri dishes and allowed to solidify. Plates were then incubated for 1 h at 37°C. To this, 1 mL of 10% complement source serum was added and the plates were further incubated for 30 min. Plaques were enumerated and evaluated by using a dissecting microscope with a 10X ocular strength.
Phagocytosis: Phagocytosis was performed according to the method of Boyum (1968). Ice-cold normal saline (5 mL) was injected forcefully with 20-gauze needle to ensure adequate mixing. Fluid was then harvested in plastic tubes using needle and kept in ice. Cell suspension was centrifuged at 1000 rpm for 10 min at 4°C. The supernatant was discarded and pellet was washed with 5 mL of ice-cold saline, mixed and recentrifuged at same speed and temperature for 5 min. Pelleted cells were re-suspended in 0.5 mL of ice-cold saline.
Viability of neutrophils by using trypan blue: For testing the viability of neutrophils, 1 mL of 0.1% trypan blue was added to an equal volume of neutrophil suspension. Number of unstained cell (viable) and stained cell (dead) were counted in Neubauer’s chamber. The percentage of viable cells was then calculated.
Phagocytic index: For the evaluation of phagocytic index, neutrophils were incubated with the prepared dead microbial culture as mentioned above. The eppendroff tubes were marked as 5, 10 and 15 min, respectively. To each of the tube, 200 μL of neutrophils suspension, 6.6 μL of 10% pooled serum (for opsonization) and 200 μL of E. coli suspension was added, mixed well and incubated for a period of 5, 10 and 15 min, respectively, at room temperature. Ice-cold HBSS (100 μL) was used for stopping the reaction. Mixture was then centrifuged at 1000 rpm for 3 min. Supernatant was discarded and pellet was resuspended thoroughly in left droplet. Four slides of mixed suspension from each of the tube were made by forming a thin smear. The smears were air dried, fixed in 100% methanol for 5 min and stained with Geimsa stain for 10 min. Stained smear was thoroughly washed with ice-cold PBS and then mounted in DPX. Numbers of E. coli associated with neutrophils were then counted at 100X of the microscope using oil immersion at different locus:
Statistical analysis: Data are expressed as the Mean±SEM. The data was analyzed using the Statistical Package for Social Science program (SPSS 11). Statistical analysis was done using analysis of variance (ANOVA) followed by Tukey test and the level of significance was set at p<0.05.
RESULTS AND DISCUSSION
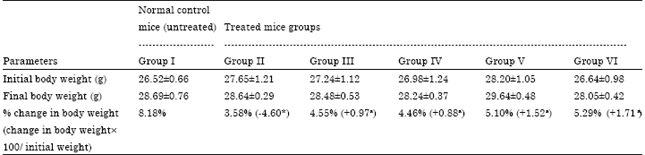
Effect on body weight: The changes in the body weight of animals were detected at the end of the experiment in different groups and are presented in Table 1. It is evident from the Table 1 that there is gradual increase in body weight with age of control animals.
| Table 1: | Ameliorating effect of Coriandrum sativum extracts on body weight of mice exposed to lead nitrate. |
 | |
| Values are Mean±SEM; n = 12. *p<0.001 compared to normal animals. ap<0.001 compared to lead nitrate exposed animals | |
The initial average body weight of mice was 26.52±0.66 (control); 27.65±1.21 (lead induced group II); 27.24±1.12 (group III); 26.98±1.24 (group IV); 28.20±1.05 (group V) and 26.64±0.98 g (group VI) in comparison with body weight (28.69±0.76, 28.64±0.29, 28.48±0.53, 28.24±0.37, 29.64±0.48 and 28.05±0.42 g in all experimental groups, respectively) after 40 days experimental period. At the termination of experiment, lead nitrate treated animals showed significant decrease in % body weight (4.60%, p<0.001) as compared to control group (8.18%), indicating a condition of weight loss. It has been demonstrated that inorganic lead selectively bound to proteins in some tissues (Valle and Ulmer, 1972), disrupts protein function and results in weight loss. Further, Kaltreider et al. (2001) also demonstrated that exposure to metal altars hormonal function in the glucocorticoid system. The glucocorticoid hormones play an important role in glucose regulation, as well as carbohydrate, lipid and protein metabolism. Dysfunction in the glucocorticoid system has been linked to weight gain/ loss. The glucocorticoid hormonal system plays an important role in protein metabolism.
But then, treatment with coriander extracts (group III, IV, V and VI) along with lead nitrate exhibited 0.97, 0.88, 1.52 and 1.71% increase in body weight, respectively, compared to relevant lead nitrate treated group. However, data regarding protection of lead induced loss in body weight gain by both the aqueous and ethanolic extract of coriander is scarcely known.
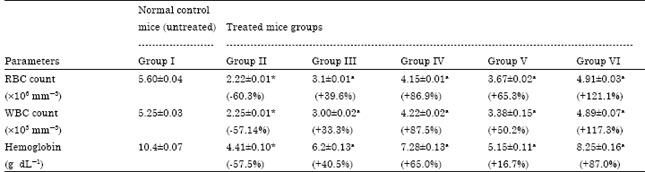
Hematological variables: Effect of lead nitrate alone and ameliorating effect of Coriandrum sativum extracts individually during lead nitrate exposure on some selected blood profiles in different groups were checked and are presented in Table 2.
In comparison to normal control mice, a significant (p<0.001) decline in the RBC count, WBC count and Hemoglobin concentration were recorded in lead nitrate exposed mice. The RBC count became lower with lead treatment, due to hemolysis (Darmono, 1995). The decrease in RBC count observed here is also in agreement with other previous reports of Lavicoli et al. (2003). The effects of erythrocyte membranes in particular, are analyzed because erythrocytes have a high affinity for lead, and are more susceptible to oxidative damage than other cells (Gurer et al., 1998) . Osmotic susceptibilities of erythrocyte were reported to be increased in lead toxicity accompanied by decreased deformability and a shortened life span (Levander et al., 1977). Decrease in WBCs by lead in the present study is in favor with earlier report of McLeay and Gorden (1977). Some researchers have observed decrease in hemoglobin concentration in lead treated animals (Bersenyi et al., 2003). Lead may inhibit the body’s ability to make hemoglobin by interfering with several enzymatic steps in the heme pathway. Studies have shown that lead inhibits the enzymes δ-aminolevulinic acid dehydratase (ALAD) and ferrochelatse of the heme synthetic pathway thus preventing conversion of ALA to porphobilinogen and inhibits incorporation of iron into the protoporphyrin ring, respectively.
| Table 2: | Ameliorating effect of Coriandrum sativum extracts on hematological variables of lead nitrate exposed mice |
 | |
| Values are Mean± SEM.; n = 12. *p<0.001 compared to normal animals. ap<0.001 compared to lead nitrate exposed animals | |
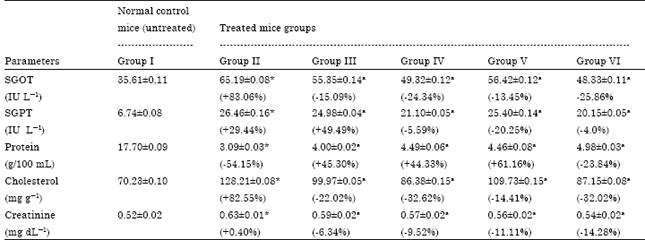
| Table 3: | Ameliorating effect of Coriandrum sativum extracts on heametochemical variables of lead nitrate exposed mice |
 | |
| Values are Mean±SEM.; n = 12. *p<0.001 compared to normal animals. ap<0.001 compared to lead nitrate exposed animals | |
This results in reduced heme synthesis and elevated levels of the precursor δ-aminolevulinic acid (ALA) which is a weak gamma-aminobutyric acid (GABA) agonist that decreases GABA release by presynaptic inhibition (Baranowska-Bosiacka et al., 2000).
Treatment with low and high dose of aqueous coriander extract significantly (p<0.001) increase lead nitrate induced decrease of RBC count, WBC count and Hemoglobin concentration as compared to lead nitrate treated animals (group II). This protective effect was more pronounced in mice supplemented with ethanolic extract of plant in group V and VI. This change was significant at p<0.001 for both low and high dose of aqueous and ethanolic extract of coriander compared to lead nitrate treated group. The protective action of coriander against lead toxicity could be attributed to the antioxidant action of its component.
Hematochemical variables: Effect of lead nitrate alone and ameliorating effect of Coriandrum sativum extracts individually during lead nitrate exposure on some selected hematopoietic biochemical variables in various groups were assessed and are presented in Table 3.
It is clear from the results that treatment with lead nitrate showed a significant elevation in some serum biochemical parameters which include SGOT, SGPT, total cholesterol and creatinine level as compared to control group animals (p<0.001). The mean value of total serum protein was significantly decreased by lead nitrate intake when compared with control (p<0.001). Lead is known to bind to the sulfhydryl groups of enzymes containing cysteine and found to form complexes with amino acids and protein. Since aminotransferases (SGOT and SGPT) are an important class of enzymes linking carbohydrate and amino acid metabolism, the relationship between the intermediates of the citric acid cycle is well established. These enzymes are regarded as markers of liver injury (Talmage et al., 1978). Increasing the activities of AST and ALT in the blood sera was most likely a consequence of the hepatotoxic effect of lead, i.e., the occurrence of toxic hepatitis. The lead entering the body by oral gavages is delivered to the liver through the portal blood circulation where the greatest part of it remains stored. Only a smaller part of this toxic metal “breaks the liver barrier” and enters the body circulation. The accumulated lead in the liver can act by directly damaging the hepatocytes, primarily by destroying the permeability of the cell membrane which results in the increased release of cytosolic enzymes AST and ALT into the circulation. It has been reported that ALT elevated significantly more than AST on lead exposure, which indicates liver damage (Shalan et al., 2005).
Cholesterol in reasonable quantities is critical to good health as it is a component of cell membranes and is found in all body tissues and is converted into various hormones. Present finding of elevated total cholesterol level supports previous reports (Nomiyama et al., 2002).
The association between lead exposure and high serum lipid levels is biologically plausible and could be due to either increased synthesis or decreased removal of lipoproteins. Decreased removal may occur as a result of the alteration of cell surface receptors for lipoproteins (Tarugi et al., 1982) or as a result of the inhibition of hepatic lipoprotein lipase activity (Chajet-Shaul et al., 1989). Furthermore, lead has been shown to depress the activity of cytochrome P-450 (Meredith et al., 1977). This can limit the biosynthesis of bile acids which is the only significant route for elimination of cholesterol from the body. Increased synthesis may be due to a lead induced increase in hepatic enzymes at important control points for de novo cholesterol synthesis, as has been found in Wistar rats (Dessi et al., 1984) or it may be due to impaired feedback inhibition. The positive association between serum lead level and serum cholesterol among exposed subjects may have important clinical implications. Other finding shows that the increase concentration of cholesterol could result in relative molecular ordering of residual phospholipids resulting in a decrease in membrane fluidity (Sushama Kumari et al., 1990).
Blood serum protein is a fairly labile biochemical system, precisely reflecting the condition of the organism and the changes happening to it under influence of internal and external factors. In the present study, decrease in protein level was noticed. Lead binds to plasmatic protein, where it causes alterations in high number of enzymes. It can also perturb protein synthesis in hepatocytes (Goering, 1993). The observed decrease in protein content of mice treated with Pb may be due to decreased hepatic DNA and RNA (Shalan et al., 2005). Hassanin (1994) and El-Zayat et al. (1996) also reported decrease in hepatic total protein content in response to lead intoxication. They attributed that to a decreased utilization of free amino acids for protein synthesis.
Serum creatinine is a function of muscle mass and diet, as well as the glomerular filtration rate (Cameron, 1992). Increase in creatinine concentration is in agreement with previous reports (Bechara, 2004). Creatinine is a waste product derived from creatine and the kidneys excrete it. Creatinine values are used as indicators of renal functions; usually increased creatinine levels do not appear unless significant renal impairment exist (Teitz, 1982). Thus, the increase serum creatinine level observed for the lead nitrate treated group seems to corroborate the noticeable damage to cortex and glomerulus.
The SGOT, SGPT, total cholesterol and creatinine levels were significantly reduced by administration of aqueous coriander extract at a dose of 300 and 600 mg kg-1 body weight (p<0.001 vs. lead nitrate intoxicated mice). In comparison to lead nitrate exposed animals (group II), total protein increased insignificantly in groups III and IV. Compared with the lead nitrate control (group II), administration of ethanolic Coriandrum sativum extract at a dose of 250 and 500 mg kg-1 body weight resulted in significant decrease (p<0.001) of SGOT, SGPT, total cholesterol and creatinine levels. The total protein content in groups V and VI significantly (p<0.001) increased in serum, when compared with lead control values (group II). The reduced concentrations of SGOT and SGPT as a result of plant extracts administration observed during the present study might probably due to presence of flavonoids in the extract. It is well documented that flavonoids and glycosides are strong antioxidants (Kavithalakshmi et al., 2006). About the cholesterol lowering effect of coriander, the increased activity of plasma LCAT, enhanced hepatic bile acid synthesis and the increased degradation of cholesterol to fecal bile acids and neutral sterols appeared to account for its hypocholesterolemic effect (Chithra and Leelamma, 1997). Further, research is necessary to identify the mode of the action of coriander seeds.
The observed decrease in these enzymes shows that aqueous and alcoholic coriander extract preserve the structural integrity of the animal from the toxic effect of Lead.
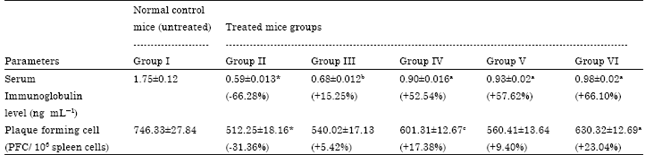
Immunological studies: Table 4 exhibits serum immunoglobulin level and plaque forming cell assay in control and experimental group of animals. The level of serum antibody and PFC showed significant decrease (p<0.001) in lead-treated animals (group II) than that of normal untreated mice. The mechanism responsible for lead induced immunosuppression is uncertain. However, any compound-induced alteration in antigen processing, presentation, cell proliferation and/or differentiation or related synthesis and/or release of interleukins, could modify this response (Luster et al., 1992). A limited amount of data suggest that the biochemical and molecular mechanisms of Pb toxicity involve the induction of oxidative stress in target cells, partly via the activation of Reactive Oxygen Species (ROS), followed by DNA damage and apoptosis (Ercal et al., 2000). Generation of ROS appears to be central for the immunotoxic effects of Pb (Fracasso et al., 2002).
Administration of aqueous coriander extract at a dose of 300 and 600 mg kg-1 body weight, significantly elevated immunoglobulin level (p<0.01 and p<0.001, respectively), compared to lead nitrate treated animal (group II). In comparison to lead nitrate exposed animals (group II), PFC increased insignificantly in group III and IV treated animals. Supplementation of ethanolic coriander extract in animals registered a significant increase (p<0.001 for both low and high dose) in both immunoglobulin level, in both plant treated group, compared with lead treated group. In comparison to lead nitrate exposed animals (group II), PFC value increased significantly in group VI (p<0.001) but insignificantly in group V treated animals. To the best of literature survey no reports are available in support for these observations.
Effect of lead nitrate alone and ameliorating effect of Coriandrum sativum extracts individually during lead nitrate exposure on viability and phagocytic index of Neutrophils of various groups were assessed and are presented in Table 5.
Lead nitrate at a dose of 40 mg kg-1 body weight caused significant (p<0.001) decrease in the viability of neutrophils and insignificant decrease in phagocytosis of neutrophils. Number of non-viable cells was higher (p<0.001) in lead exposed group II than in normal untreated group. However, treatment with aqueous coriander extract along with lead nitrate caused a significant elevation (p<0.001 for both low and high dose) in viability of neutrophiles when compared with lead induced group.
| Table 4: | Ameliorating effect of Coriandrum sativum extracts on serum immunoglobulin level and on PFC in lead nitrate exposed mice |
 | |
| Values are Mean±SEM.; n = 12. *p<0.001 compared to normal animals. ap<0.001 compared to lead nitrate exposed animals. bp<0.01 compared to lead nitrate exposed animals. cp<0.02 compared to lead nitrate exposed animals | |
| Table 5: | Ameliorating effect of Coriandrum sativum extracts on viability and phagocytic index of neutrophils in lead nitrate exposed mice |
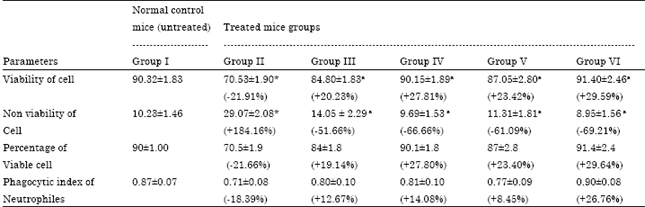 | |
| Values are Mean±SEM.; n = 12. *p<0.001 compared to normal animals. ap<0.001 compared to lead nitrate exposed animals | |
Lead treatment caused marked depletion in viable cells and percent phagocytosis. Present result is compatible with the research of several workers, who noted that lead has been shown to inhibit neutrophil phagocytosis in occupationally exposed workers in comparison to normal healthy humans (Zelikoff et al., 1993). These results suggest that lead may interfere with antigen specific interaction between macrophages/neutrophils and T cells.
An insignificant increase in phagocytosis was observed after the treatment with aqueous coriander extract, in comparison to lead nitrate exposed group II. Supplementation of ethanolic extract of coriander offered significant increase in viability (p<0.001 for both low and high dose) and insignificant increase in phagocytosis of neutrophiles in both groups, compared to group II. While administration of the same dose significantly decrease the nonviability of neutrophiles in group V and VI, compared to lead group (p<0.001 for both low and high dose).
The antioxidative property of coriander seed is related to the large amounts of tocopherols, carotenoids and phospholipids (Ramadan and Morsel, 2003) which act through different mechanisms. Carotenoids act as primary antioxidants by trapping free radicals and as secondary antioxidants by quenching singlet oxygen (Reische et al., 2002). Tocopherols and sterols interact with oil surfaces and release hydrogen, inhibiting the propagation step of radical reactions (Reische et al., 2002). Synergetic effects were evidenced with combinations of carotenoids and tocopherols (Reische et al., 2002). Although the exact mechanism of antioxidative action of phospholipids is not still fully established, these substances would synergistically act with tocopherols, would form barrier for O2 between air/oil interfaces would favor formation of Mallard like compounds with oxidation products or would chelate pro-oxidant metals with phosphate groups (Haila et al., 1996). Thus, Administration of Coriandrum sativum caused a significant increase in serum antibody level and PFC assay compared to lead induced group supporting the antioxidant effect of plant.
CONCLUSION
It could be concluded from the present results that supplementation of Coriandrum sativum possessed a significant protective value as a potent antioxidant activity against a few lead sensitive biochemical variables, hematological and immunological parameters in blood and serum of mice. Further work need to be done to isolate and purify the active principle involved in the antioxidant activity of this plant.
ACKNOWLEDGMENTS
The authors are thankful to the authorities of Banasthali University for providing support to the study.
REFERENCES
- Sivaprasad, R., M. Nagraj and P. Varalakshmi, 2003. Combined efficacies of lipoic acid and meso-2, 3-dimercaptosuccinic acid on lead induced erythrocyte membrane lipid peroxidation and antioxidant status in rats. Hum. Exp. Toxicol., 22: 183-192.
CrossRefDirect Link - Gerber, G.B., J. Maes, N. Gilliavod and G. Casale, 1978. Brain biochemistry of infant mice and rats exposed to lead. Toxicol. Lett., 2: 51-63.
CrossRef - Cohen, M.D., Z. Yang and J.T. Zelikoff, 1994. Immunotoxicity of particulate lead: In vitro exposure alters pulmonary macrophage tumor necrosis factor production and activity. J. Toxicol. Environ. Health., 42: 377-392.
PubMedDirect Link - Kishikawa, H., R. Song and D.A. Lawrence, 1997. Interleukin-12 promotes enhanced resistance to Listeria monocytogenes infection of lead exposed mice. Toxicol. Applied Phormacol., 147: 180-189.
CrossRef - Fracasso, M.E., L. Perbellini, S. Solda, G. Talamini and P. Franceschetti, 2002. Lead induced DNA strand breaks in lymphocytes of exposed workers: Role of reactive oxygen species and protien kinase C. Mutat. Res./Genet. Toxicol. Environ. Mutagenesis, 515: 159-169.
CrossRefDirect Link - Patra, R.C., D. Swarup and S.K. Dwivedi, 2001. Antioxidant effects of a-tocopherol, ascorbic acid and L-methionine on lead induced oxidative stress to the liver, kidney and brain in rats. Toxicology, 162: 81-88.
PubMedDirect Link - Varnai, V.M., M. Piasek, M. Blanusa, M.M. Saric, D. Juresa and K. Kostial, 2004. Succimer treatment and calcium supplementation reduce tissue lead in suckling rats. J. Applied Toxicol., 24: 123-128.
CrossRefDirect Link - Burdock, G.A. and I.G. Carabin, 2009. Safety assessment of coriander (Coriandrum sativum L.) essential oil as a food ingredient. Food Chem. Toxicol., 47: 22-34.
CrossRefPubMedDirect Link - Eidi, M., A. Eidi, A. Saeidi, S. Molanaei, A. Sadeghipour, M. Bahar and K. Bahar, 2009. Effect of coriander seed (Coriandrum sativum L.) ethanol extract on insulin release from pancreatic β cells in streptozotocin-induced diabetic rats. Phytother. Res., 23: 404-406.
CrossRefDirect Link - Chithra, V. and S. Leelamma, 2000. Coriandrum sativum effect on lipid metabolism in 1, 2-dimethyl hydrazine induced colon cancer. J. Ethnopharmacol., 71: 457-463.
CrossRef - Omura, Y., Y. Shimotsuura, A. Fukuoka, H. Fukuoka and T. Nomoto, 1996. Significant mercury deposits in internal organs following the removal of dental amalgam and development of pre-cancer on the gingiva and the sides of the tongue and their represented organs as a result of inadvertent exposure to strong curing light (used to solidify synthetic dental filling material) and effective treatment: A clinical case report, along with organ representation areas for each tooth. Acupuncture Electro-Ther. Res., 21: 133-160.
PubMedDirect Link - Gray, A.M. and P.R. Flatt, 1999. Insulin-releasing and insulin-like activity of the traditional anti-diabetic plant Coriandrum sativum (coriander). Br. J. Nutr., 81: 203-209.
CrossRefPubMedDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Jerne, N.K. and A.A. Nordin, 1963. Plaque formation in agar by single antibody producing cells. Science, 140: 405-406.
CrossRefDirect Link - Boyum, A., 1968. Isolation of mononuclear cells and granulocytes from human blood. Isolation of monuclear cells by one centrifugation and of granulocytes by combining centrifugation and sedimentation at 1g. Scand. J. Clin. Lab. Invest., 97: 77-89.
PubMedDirect Link - Vallee, B.L. and D.D. Ulmer, 1972. Biochemical effects of mercury, cadmium and lead. Annu. Rev. Biochem., 41: 91-128.
CrossRefPubMedDirect Link - Kaltreider, R.C., A.M. Davis, J.P. Lariviere and J.W. Hamilton, 2001. Arsenic alters the function of the glucocorticoid receptor as a transcription factor. Environ. Health Persp., 109: 245-251.
PubMedDirect Link - Iavicoli, I., G. Carelli, E.J. Stanek, N. Castellino and E.J. Calabrese, 2003. Effects of low doses of dietary lead on red blood cell production in male and female mice. Toxicol. Lett., 137: 193-199.
CrossRef - Gurer, H., H. Ozgunes, R. Neal, D.R. Spitz and N. Ercal, 1998. Antioxidant effects of N-acetylcysteine and succimer in red blood cells from lead-exposed rats. Toxicology, 128: 181-189.
CrossRefDirect Link - Levander, O.A., V.C. Morris and R.J. Ferretti, 1977. Filterability of erythrocytes from vitamin E deficient lead-poisoned rats. J. Nutr., 107: 363-372.
PubMedDirect Link - Bersenyi, A., S.G. Fekete, Z. Szocs and E. Berta, 2003. Effect of ingested heavy metals (Cd, Pb and Hg) on haematology and serum biochemistry in rabbits. Acta Veterinaria Hungarica, 51: 297-304.
CrossRefPubMedDirect Link - Baranowska-Bosiacka, I., A.J. Hlynczak and B. Machalinski, 2000. The impact of lead ions on metabolism of erythrocytes. Med. Pr., 51: 59-65.
PubMedDirect Link - Talmage, R.V., C.J.V. Wiel and H. Norimatsu, 1978. A reevaluation of the cause of acute hypercalcemia following intravenous administration of lead acetate. Calcified Tissue Res., 26: 149-153.
CrossRef - Shalan, M.G., M.S. Mostafa, M.M. Hassouna, S.E.H. El-Nabi and A. El-Refaie, 2005. Amelioration of lead toxicity on rat liver with vitamin C and silymarin supplements. Toxicology, 206: 1-15.
CrossRefPubMedDirect Link - Nomiyama, K., H. Nomiyama, S.J. Liu, Y.X. Tao, T. Nomiyama and K. Omae, 2002. Lead induced increase of blood pressure in female lead workers. Occup. Environ. Med., 59: 734-738.
PubMedDirect Link - Tarugi, P., S. Calandra, P. Borella, G.F. Vivoli, 1982. Heavy metals and experimental atherosclerosis: Effect of lead intoxication on rabbit plasma lipoproleins. Atherosclerosis, 45: 221-234.
Direct Link - Chajet-Shaul, T., G. Friedman, O. Stein, F. Shiloni, J. Etienne and Y. Stein, 1989. Mechanism of the hypertriglyceridemia induced by tumor necrosis fac-tor administration to rats. Biochim. Biophys. Acta, 1001: 316-324.
CrossRef - Meredith, P.A., B.C. Campbell and A. Goldberg, 1977. The effect of industrial lead poisoning on cytochrome P-450 mediated phenazone hydroxylation. Eur. J. Clin. Pharmacol., 12: 235-239.
CrossRef - Dessi, S., B. Batcna, E. Laconi, C. Enms and P. Pani, 1984. Hepatic cholesterol in lead nitrate induced liver hyperplasia. Chem. Biol. Interact., 48: 271-279.
CrossRef - Kumari, S.S., A. Verghese, D. Muraleedharan and V.P. Menon, 1990. Protective action of aspirin in experimental myocardial infarction induced by isoproterenol in rats and its effect on lipid peroxidation. Indian J. Exp. Biol., 28: 480-485.
PubMedDirect Link - Goering, P.L., 1993. Lead-protein interactions as a basis for lead toxicity. Neurotoxicology, 14: 45-60.
PubMed - Chithra, V. and S. Leelamma, 1997. Hypolipidemic effect of coriander seeds (Coriandrum sativum): Mechanism of action. Plant Foods Hum. Nutr., 51: 167-172.
CrossRefDirect Link - Luster, M.I., C. Portier, D.G. Pait, K.L. White Jr., C. Gennings, A.E. Munson and G.J. Rosenthal, 1992. Risk assessment in immunotoxicology: I. Sensitivity and predictability of immune tests. Fundam. Applied Toxicol., 108: 200-210.
CrossRef - Ercal, N., R. Neal, P. Treeratphan, P.M. Lutz, T.C. Hammond, P.A. Dennery and D.R. Spitz, 2000. A role for oxidative stress in suppressing serum immunoglobulin levels in lead exposed Fisher 344 rats. Arch. Environ. Contam. Toxicol., 39: 251-256.
PubMedDirect Link - Zelikoff, J.T., E. Parsons and R.B. Schlesinger, 1993. Inhalation of particulate lead oxide disrupts pulmonary macrophage-mediated functions important for host defense and tumor surveillance in the lung. Environ. Res., 62: 207-222.
CrossRef - Haila, K.M., S.M. Lievonon and M.I. Heinonen, 1996. Effects of lutein, lycopene, annatto and γ-tocopherol on autoxidation of triglycerides. J. Agric. Food Chem., 44: 2096-2100.
CrossRefDirect Link