
O.E. Okon
Department of Zoology and Environmental Biology, University of Calabar, Calabar, Nigeria
K.N. Opara
Department of Zoology, University of Uyo, Uyo, Nigeria
S.E. Etim
Department of Biological Sciences, Cross River University of Technology, Calabar, Nigeria
G.A. Arong
Department of Zoology and Environmental Biology, University of Calabar, Calabar, Nigeria
E.E. Oku
Department of Zoology and Environmental Biology, University of Calabar, Calabar, Nigeria
C.I. Iboh
Department of Biological Sciences, Cross River University of Technology, Calabar, Nigeria
Research Journal of Parasitology
Year: 2011 | Volume: 6 | Issue: 5 | Page No.: 168-175







ABSTRACT
Tick infestation is highly significant in production of livestock for meat, milk and other resources. Experimental transmission of B. bigemina between cattle and ticks (Boophilus decoloratus) were carried out in the laboratory. Body bags were applied on the bodies of two calves infected with B. bigemina. B. decoloratus larvae were then introduced into these body bags to make contact with the calves, Engorged female ticks were then collected several days later as the dropped from the calves. Haemolymph smears of the engorged females B. decoloratus derived from calves 28B (a two month old intact male Ayrshire calf) and 10B (a four month old splenectomized male Friesian calf) revealed 93 (35%) and 112 (34%) infection rates with kinetes of B. bigemina, respectively. Twelve percent mortality rate was reported for ticks derived from calf 28B while 5% was reported for ticks derived from calf 10B before the commencement of egg laying. The bloods of the two calves were found to show declining Packed Cell Volume (PCV) values and red blood cell counts. B. decoloratus was found to transmit B. bigemina in cattle with a high level of success.
PDF Abstract XML References Citation
Received: February 03, 2012;
Accepted: April 28, 2012;
Published: June 20, 2012
How to cite this article
O.E. Okon, K.N. Opara, S.E. Etim, G.A. Arong, E.E. Oku and C.I. Iboh, 2011. Experimental Transmission of Babesia bigemina by Boophilus decoloratus in Cattle. Research Journal of Parasitology, 6: 168-175.
URL: https://scialert.net/abstract/?doi=jp.2011.168.175
URL: https://scialert.net/abstract/?doi=jp.2011.168.175
INTRODUCTION
Arthropod vectors of Babesia are all hard ticks of the family Ixodidae (Koch 1844). In general, each species of Babesia is associated with a single vector species which may differ from region to region (Baranton et al., 2001). Hoyte (1979) observed that in Australia and South America, the vector of Babesia bovis is Boophilus microplus. Reports of the development of Babesia species in tick vectors are numerous, such as those of B. bigemina in B. microplus (Stewart et al., 1986; Higuchi et al., 1992). Babesia argentina in B. microplus (Guglielmone, 1992) and B. canis in Haemaphysalis leachi (Bashir et al., 2009).
Bock et al. (2004) in their study in Europe showed that B. bovis infect cattle in those areas where Rhipicephalus bursa were present, whereas Babesia divergens was the main cattle parasite where Ixodes ricinus were present. Morzaria (1975) described some stages in the development of Babesia major in H. punctata. Babesia microti is transmitted by ixodid ticks (Oliviera and Kreier, 1978).
B. bovis and B. bigemina are apicomplexan hemoparasites of cattle that are transmitted by one-host Boophilus ticks (Stiller et al., 2002; Ueti et al., 2005). Several tick species including one-host tick B. decoloratus have been reported to transmit B. bigemina in South Africa. Throughout East Africa, B. decoloratus and R. appendiculatus are reported as a vector of B. bigemina (Morzaria, 1975).
Purnell et al. (1970) attempted to transmit B. bigemina with R. appendiculatus without success. Morzaria et al. (1977) in their experiment established B. decoloratus as a natural vector of B. bigemina in Kenya. They found that female B. decoloratus ticks became infected during feeding on an animal with a patent B. bigemina parasitaemia.
The objective of the present study was to assess the transmission of B. bigemina between cattle and experimentally reared B. decoloratus. This study will enhance the planning of appropriate control strategies for bovine babesiosis due to B. bigemina.
MATERIALS AND METHODS
Rearing of babesia-free batch of Boophilus decoloratus: Several engorged uninfected B. decoloratus were brought from the South Africa, Bureau of Standard Field Station (SABS), South Africa and reared in the Babesia Research Laboratory, Imperial College, London. The engorged ticks were placed in Petri dishes and maintained in an incubator from the date of arrival at 26-28°C. Humidity was kept at 85% using saturated potassium chloride (KCl). Dates of egg laying and larval emergence were monitored.
After complete larval emergence, the larvae were transferred from the incubator at 26-28°C to another incubator at 15-17°C under the same humidified condition. The essence of this sharp reduction in the temperature of incubation was to reduce the activity of the larvae under this non-feeding stage of its life history. The larvae were then sent at appropriate time to the Centre for Tropical and Veterinary Medicine (CTVM), Edinburgh to establish infection with B. bigemina.
Infection of calves with B. bigemina: Two calves maintained at the centre for tropical and veterinary medicine, Edinburgh were used for the experiment. The inoculations of B. bigemina, application of ticks, collection of ticks and monitoring of the calves were done by researchers at CTVM.
The two calves were:
| • | A 4 month old splenectomised male Friesian calf number 10B previously inoculated with B. bigemina piroplasms from the in vitro culture of the Babesia laboratory |
| • | A 2 month old intact male Ayrshire calf number 28B infected with 1 mL of B. bigemina Muguga blood stabilate. The 1 mL of blood stabilate was prepared from calf 634 (at 2.2% parasitaemia in 5% Dimethyl Sulfoxide) and 1 mL of RBC in autologous serum at 50% PCV. The blood stabilates were injected into calf 28B 17 days following larval application |
Monitoring of calves and application of ticks: Daily rectal temperatures of both calves were monitored. Blood smear, PCV and RBC counts of calf 28B were examined three times weekly before inoculation of parasites. After inoculation of parasites, they were examined daily. Blood smear, PCV and RBC counts of calf 10B were examined three times weekly.
Body bags were applied to the bodies of the calves by methods previously described by Thompson (1976) and Hodgson et al. (1989) with modifications. 14, 000 B. decoloratus larvae were applied using the same procedure on calf 28B but in this case the B. bigemina stabilates were injected after 17 days of its application. Engorged female ticks were collected as they dropped from the calf and dispatched in sample bottles plugged with wet cotton wool to the Babesia laboratory.
Screening of the engorged females for the kinetes of B. bigemina: Giemsa stained haemolymph smears of the engorged ticks were examined for the presence of kinetes. This was done by draining haemolymph from the third or fourth pair of legs onto a glass slide within the first 10 days of dropping from the host. They were fixed in absolute ethanol for 1 min, stained with 10% Giemsa and examined under the oil immersion objective (Mahoney and Mirre, 1971).
The degrees of infectivity of the ticks were ranked as 1+ for those with one kinete per microscopic field, 2+ for those with two to three kinetes per field and 3+ for those with four kinetes and above per microscopic field.
RESULTS
Experimental transmission of Babesia bigemina in the laboratory
Rearing of Babesia free Boophilus decoloratus: The Babesia free engorged Boophilus decoloratus ticks brought from SABS commenced egg laying a day after arrival and continued for 12 days. The round, brownish eggs were laid in masses. Each tick was estimated to lay 1000-3000 eggs. Larvae emergence commenced 2 weeks after egg laying and lasted for about 12 days. The larvae were minute, very mobile organisms bearing 3 pairs of legs as against the 4 pairs found in the adult.
Infection of the calves: Calf 28B which was infected 17th day following application of larvae showed signs of B. bigemina in the blood smear from the 19th day till the end of the experiment. From the first to the 19th day of the experiment, the PCV and the RBC counts ranged between 31-35% and 8.55-9.97x106 μL-1, respectively. The PCV and RBC counts dropped after the 19th day to the end of the experiment ranging from 23-27% and 6.91-7.71x106 μL-1, respectively (Table 1). The rectal temperature of calf 28B ranged between 38.2-40.2°C throughout the duration of the experiment (Table 1).
Before the commencement of the experiment, calf 10B (the 4 month old splenectomized male Friesian calf) was earlier challenged with B. bigemina piroplasm from in vitro tissue culture maintained in the laboratory. The rectal temperature, RBC count and Packed Cell Volume (PCV) of the calf monitored throughout the course of the experiment ranged between 38.4-39.8°C, 6.33-7.85x106 μL-1 and 24-28%, respectively (Table 2).
Infection of the ticks: Haemolymph smears of the 329 engorged female ticks derived from calf 10B revealed 112 (34%) infection rate with the kinete stage of B. bigemina. The degree of infectivity ranked as 1+, 2+ and 3+ were 19, 9 and 7%, respectively (Table 3). Figure 1 illustrates kinetes at 3+ degree of infectivity derived from the haemolymph of B. decoloratus.
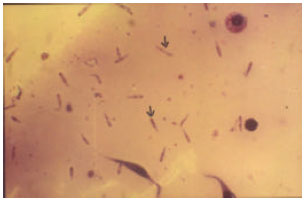 | |
| Fig. 1: | Kinete stage of B. bigemina at 3+ degree of infectivity derived from haemolymph smear of engorged B. decoloratus, magnification x1000 |
| Table 1: | Experimental infection of calf 28B with B. bigemina (African) blood stabilate and subsequent application of Boophilus decoloratus larvae |
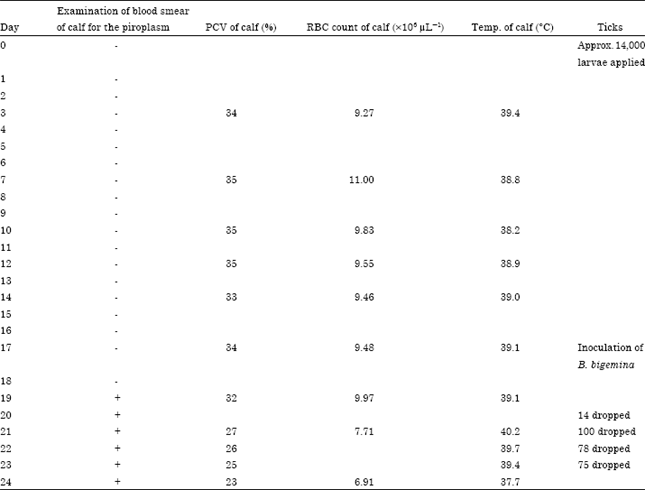 | |
| -: Negative , +: Positive, for B. bigemina | |
| Table 2: | Experimental infection of calf 10B with in vitro cultured B. bigemina (African) and simultaneous application of Boophilus decoloratus larvae |
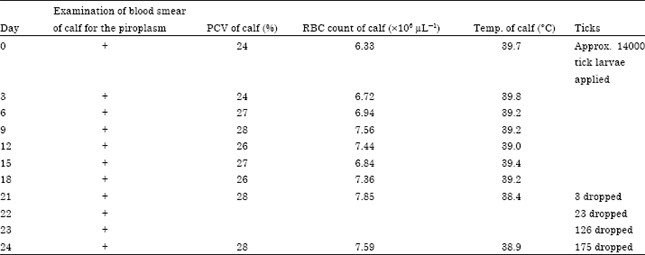 | |
| +: Positive for B. bigemina | |
| Table 3: | Summary of the degree of infectivity of the ticks applied on the two strains of calves |
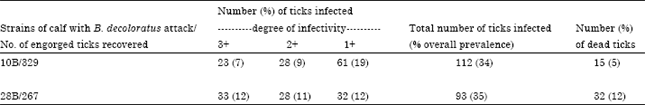 | |
Haemolymph smears of the 267 engorged female ticks derived from calf 28B revealed 93 (35%) infection rate with the kinete stages of B. bigemina. The degree of infectivity ranked as 1+, 2+ and 3+ were 12, 11 and 12%, respectively (Table 3). The number of dead ticks recovered from calf 10B and 28B after dropping were 15 (5%) and 31 (12%), respectively.
DISCUSSION
Experimental evidence indicate that the haemolymph smears of engorged Boophilus decoloratus derived from calves 10B and 28B revealed 34 and 35% infection rates with B. bigemina, respectively. Bock et al. (1999) found 36% infection rate of ticks infected with B. bigemina through a similar haemolymph examination. This result is in consonance with our present studies on both calves 10B and 28B examined. In earlier studies although exact values were not given, the prevalence of the bovine Babesia within the progeny of infected female ticks was found to be very low (Mahoney and Mirre, 1971; Riek, 1964, 1966). Although, all the ticks applied on the calves were subjected to the same sets of condition on the different calves, not all were infected, probably due to physiological or genetic variations in the strains of B. decoloratus. Another likely reason for the low percentage of infected ticks was attributed to the fact that some of the female ticks completed engorgement when the parasite density in the bovine host blood was below a particular critical level (Friedhoff and Smith, 1981). Thus, the proportion of ticks infected with B. bigemina reflects the parasite rate of their bovine host. In contrast, in strains of B. bovis that was highly adapted to ticks, low cattle parasitaemia (0.1%) produced infection in 100% of the ticks (Friedhoff and Smith, 1981; Friedhoff, 1988). This contradicts the result of the present studies where 34 and 35% tick infection rates were obtained from calves 10B and 28B, respectively.
Twelve percent mortality rate was reported for ticks derived from calf 28B while only five percent was reported for ticks derived from calf 10B before the commencement of egg laying. Thus a higher death rate was found in ticks derived from calf 28B than ticks from calf 10B. This was attributed to the fact that the larvae of ticks applied on calf 10B were exposed to B. bigemina from the onset of the experiment; that is calf 10B was infected prior to application of larvae. Hence the engorged ticks were more adapted to the B. bigemina in calf 10B, thus facilitating a higher survival rate. On the contrary, calf 28B picked up the B. bigemina on the 19th day of the experiment when the ticks were almost dropping from the calf. Hence they could not cope with the sudden infection of the calf with B. bigemina, therefore, their higher mortality rate. Friedhoff and Smith, 1981 found that one strain of B. decoloratus had mortality rates of more than 80% of the engorged female ticks after alimentary infection with B. bigemina. This result is at variance with the present study which reported 12 and 5% mortality rates from calves 28B and 10B, respectively. Gray (1982) reported that the ingestion of large numbers of Babesia may have a negative effect upon female tick survival and reproduction. However, others found little effect of B. bigemina and B. bovis on B. microplus (Ouhelli et al., 1987; Guglielmone et al., 1989).
After the inoculation of the calves with the B. bigemina, there was a drastic fall in the PCV of the two calves. This was sequelled from the report collected from the centre for Tropical and Veterinary Medicine, Edinburgh. Calf 28B which was Babesia free for the first 19 days of the application of B. decoloratus larvae, had a PCV of 31-35%, After acquiring the infection the PCV dropped to 23-27%. Also, since calf 10B was previously infected before the commencement of the experiment, the PCV dropped to 24-28% throughout the duration. Similarly, calf 28B had RBC count of 8.55-11.0x106 μL-1 from day 1 to day 19 of the experiment. The RBC count dropped after day 19 to 6.91-7.71x106 μL-1. Also RBC count of calf 10B ranged from 6.33-7.85x106 μL-1 throughout the duration of the experiment. Hence there appears to be a relationship between bovine babesiosis, PCV and RBC count. It was observed that the destruction of the red blood cells by the parasites has an adverse effect on the RBC count and PCV because they both showed marked decline. Animals vaccinated with crude soluble parasite antigens essentially showed patent Babesia infections of shorter duration and a less marked fall in PCV values (Schetter and Montenegro-James, 1995).
Wright (1973) carried out a comparative study of the haematology of experimentally induced B. bovis and B. bigemina infections in cows. Six animals were each infected with B. bigemina by the inoculation of 500 mL of blood from a carrier animal. There was rapid fall in the total erythrocyte count from 7.6x106 mm-3 on day 4 to a minimum of 2.1x106 mm-3 on day 11. The value then persisted at about this level until all the animals had died. Their results were in agreement with those of Wright (1973) and also with that of the present study.
Calf 10B in which the uninfected B. decoloratus larvae were applied was earlier splenectomized before being inoculated with B. bigemina from the in vitro culture. The essence of the splenectomy was to ensure that the calf losses resistance immediately and then acquire the B. bigemina infection. Zwart and Brocklesby (1979) found that the spleen is important in non-specific and immunological defence of the host against Babesia species; reflected in the histopathological changes found in the spleen during Babesia infection. Hence, it was necessary to ensure that the calf adequately acquired the parasite by reducing their immune resistance.
The present investigation showed that bovine babesiosis has a negative impact on the physiological functions of infected animals. This is evident in the reduction of the PCV values and the RBC counts of infected calves. Hence, the control of this infection should be given adequate attention in the livestock industry. Moreover, B. decoloratus has able proven to be the vector of B. bigemina. Even though, vectors of B. bigemina vary from region to region, in the control of bovine babesiosis, emphasis should be placed on the elimination of B. decoloratus and all other tick species in any field studies. The present studies will however stimulate field studies in various regions where the vector species have not yet been established.
ACKNOWLEDGMENT
We wish to express our sincere gratitude to the researchers at the centre for Tropical Veterinary Medicine, Edinburgh for inoculating B. bigemina to their calves, for applying the ticks on the calves and for monitoring the calves.
REFERENCES
- Baranton, G., G. Seinost, G. Theodore, D. Postic and D. Dykhuizen, 2001. Distinct levels of genetic diversity of Borrelia burgdorferi are associated with different aspects of pathogenicity. Res. Microbiol., 152: 149-156.
CrossRefPubMedDirect Link - Bashir, I.N., Z.I. Chaudhry, S. Ahmed and M.A. Saeed, 2009. Epidemiological and vector identification studies on canine babesiosis. Pak. Vet. J., 29: 51-54.
Direct Link - Bock, R., L. Jackson, A. de Vos and W. Jorgensen, 2004. Babesiosis of cattle. Parasitology, 129: S247-S269.
CrossRefPubMedDirect Link - Bock, R.E., T.G. Kingston and A.J. de Vos, 1999. Effect of breed of cattle on transmission rate and innate resistance to infection with Babesia bovis and B. bigemina transmitted by Boophilus microplus. Aust. Vet. J., 77: 461-464.
CrossRefDirect Link - Gray, J.S., 1982. The effects of the piroplasm Babesia bigemina on the survival and reproduction of the blue, tick, Boophilus decoloratus. J. Invertebrate Pathol., 39: 413-415.
PubMedDirect Link - Guglielmone, A.A., 1992. The level of infestation with the vector of cattle babesiosis in Argentina. Memorias Inst. Oswaldo Cruz, 87: 133-137.
CrossRefPubMedDirect Link - Guglielmone, A.A., A.J. Mangold, D.H. Aguirre, A.B. Gaido and A.A. De Olsen, 1989. The effect of infection by Babesia sp. on some biological parameters of engorged females of Boophilus microplus. Folia Parasitol., 36: 1-6.
PubMedDirect Link - Mahoney, D.F. and G.B. Mirre, 1971. Bovine babesiasis: Estimation of infection rates in the tick vector Boophilus microplus (Canestrini). Ann. Trop. Med. Parasitol., 65: 309-317.
PubMedDirect Link - Ueti, M.W., G.H. Palmer, L.S. Kappmeyer, M. Statdfield, G.A. Scoles and D.P. Knowles, 2005. Ability of the vector tick Boophilus microplus to acquire and transmit Babesia equi following feeding on chronically infected horses with low-level parasitemia. J. Clin. Microbiol., 43: 3755-3759.
CrossRefDirect Link - Ouhelli, H., V.S. Pandey and A. Aboughal, 1987. Effect of infection by Babesia species on the development and survival of free living stages of Boophilus annulatus. Vet. Parasitol., 23: 147-154.
PubMed - Purnell, R.E., D. Branagan and C.G.D. Brown, 1970. Attempted transmission of some piroplasms by Rhipicephalid ticks. Trop. Anim. Health Prod., 2: 146-150.
Direct Link - Thompson, K.C., 1976. A technique to establish a laboratory colony of Boophilus microplus infected with Babesia bigemina. Vet. Parasitol., 2: 223-238.
CrossRef - Zwart, D. and D.W. Brocklesby, 1979. Babesiosis: Non-specific resistance, immunological factors and pathogenesis. Adv. Parasitol., 17: 49-113.
PubMed