
Rahul K. Keswani
Department of Chemical Engineering, Indian Institute of Technology,
Bombay, Powai, Murnbai-400 076, India
Jayesh R. Bellare
Department of Chemical Engineering, Indian Institute of Technology,
Bombay, Powai, Murnbai-400 076, India
Research Journal of Parasitology
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 31-41







ABSTRACT
Research on attraction of mosquitoes towards their hosts has recently received a boost with health organizations raising alarms on the spread of diseases through mosquito vectors. This report reviews the findings of current studies being conducted and presents useful analysis on the same. The present research shows that no particular attractant is the sole reason for the mosquitoes in locating their host. It is a blend of different odours and compounds like CO2, short chain carboxylic acids, ammonia and other sweat compounds that contribute to the olfaction and subsequent attraction of these pests. Various customized experimental models have been used throughout the world for carrying out research in this relatively new field. Experimental and technical aspects of mosquito research have been covered in great detail with particular importance on the use of new technology. Genomic studies have been briefed upon and certain useful aspects of mosquito research have been covered at the end of the report including the identification and location of odorant receptors in mosquitoes. Genomic results have only contributed to the further understanding of the exact mechanisms and bodily functions involved in location of potential hosts for blood feeding.
PDF Abstract XML References
How to cite this article
Rahul K. Keswani and Jayesh R. Bellare, 2006. A Review of Mosquito Attraction Studies: Important Parameters and Techniques. Research Journal of Parasitology, 1: 31-41.
URL: https://scialert.net/abstract/?doi=jp.2006.31.41
URL: https://scialert.net/abstract/?doi=jp.2006.31.41
INTRODUCTION
Many chemical and visual lures are used to attract insects thus playing a vital role in the monitoring and reduction of insect population particularly of those species that are a nuisance to the human population like mosquitoes, tsetse flies, locusts and others.
Mosquitoes are carriers of disease causing parasites like malaria, yellow fever, encephalitis, to name a few and the severity of these diseases in certain pockets of the world is a matter of huge concern for many health organizations. Controlling of these pests is largely limited to use of repellents and insecticides. Recently, steps have been taken to study the attraction of these insects to their hosts and thereby attempt to cut off the lethal action of these pests at the source itself (Rose and Geier, 2004).
This review aims to study the basic aspects of mosquito attraction towards humans. It should be noted that attraction refers to the whole process right from activation of neural senses to arrival at host. However, it has been noted that many studies relate their results only based on the end result of a catch. A study of different chemicals and their attraction as a blend towards mosquitoes, sampling procedures to determine the prevalence of certain mosquito species and the various experimental techniques used, data handling and the human touch to these experiments have been analyzed minutely. Genomic aspects, albeit in the nascent stage, is also a widespread area of research.
Olfaction, Attraction and Related Studies
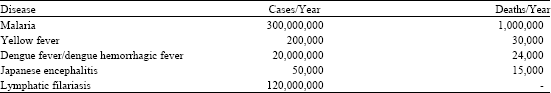
Having evolved themselves as blood-suckers from innocent plant juice feeders, mosquitoes have become the top reason for human and animal deaths in the present world (Table 1).
Differential Attractiveness
The primary cause of attraction of hosts towards mosquitoes is certain semiochemicals, essentially kairomones and 340 of which have already been identified by US agricultural chemist Ulrich R. Bernier (Barnard et al., 2000).
| Table 1: | Various mosquito-related diseases and deaths (Rose et al., 2004) |
 | |
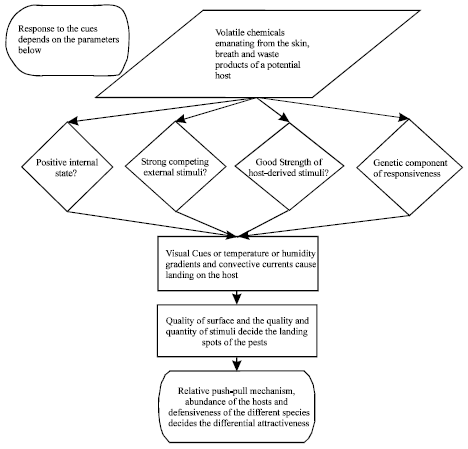 | |
| Fig. 1: | Flow-Chart for exact mosquito attraction response pattern |
However, all species are not equally attracted to humans (Mukabana et al., 2002) and hence show specific anthropophilic or zoophilic behaviour. The flowchart (Fig. 1) explains the basic behaviour of mosquitoes on locating a potential host. It has been reasoned out that a push-pull model comes into play involving attractive and repelling components, the relative abundance of the hosts and the defensiveness of different species of hosts (Costantini et al., 1998). A comparison of the feeding patterns of mosquitoes vis-a-vis the humans and animals has been studied (Gomes et al., 2003).
There is also documented evidence of the variability of attraction of mosquitoes to humans depending on age, size and other factors including pregnancy in females (Lindsay et al., 2000). The aforesaid evidence points to certain physiological changes like the ontogenic development of skin glands and other skin features that give variation in skin microflora and odour profile (Braks et al., 1999].
Odours
Rather than a single compound, it is a blend of different compounds and concentrations that stimulate the sensilla of the mosquitoes. Such blends contain a variety of compounds like lactic acid, ammonia and other sweat compounds. It is not surprising that CO2, which has been widely regarded as the primary cause for such attraction, does not figure in this blend (Barnard et al., 2000). Foot Odour plays a pivotal role in attraction and hence it does not come as a surprise that biting preferentially occurs on feet or ankles of a naked human host. The cause for the same has been identified as resident combinations of skin microflora that give out an odour similar to that of Limburger Cheese (Knols et al., 1996). A species of mosquitoes Anopheles atroparvus is known to go mainly for the face, which shows an affinity for human breath and sweat-related compounds (Vanden Broek et al., 1999). Convection currents along the body are also known to drive the mosquitoes to the lower parts of the body. Socks worn by humans also attract a fair share of these insects when combined with CO2 (Barnard et al., 2000). Synthetic host-odours like those of carboxylic acids and enols have also been shown to stimulate the mosquitoes using electroantennogram measurements (Vanden Broek et al., 1999). The fine scale structure of host odour plumes modulates capture of Anopheles gambiae and Aedes aegypti. Reasons behind the aforesaid observations include the variability in plume structure, rate of encountering pockets of kairomones, odour concentrations which affect the upwind velocities and flights of the mosquitoes (Dekker et al., 2001).
Microfluora and Parasites on Host Body
Skin-microfluora includes bacteria on the host body that produce a host of chemicals and odours. The microbial breakdown products are responsible for a distinctive human olfactory signature. Direct microbial involvement in the production of kairomones has been demonstrated by showing that incubated sweat enhances its attractiveness towards mosquitoes (Meijerink et al., 2001). Certain strains of bacteria are known to break down triglycerides on the skin to form short and long chain fatty acids that signal the mosquitoes. Pungent foot odour is also formed by similar strains of bacterium. The relative prevalence of these bacteria on the host has been identified as the key towards differential attractiveness of the hosts towards mosquitoes.
Parasite infections can change an individual's odour by changing the composition of residential skin microbes and by affecting the immunological and endocrine system, thus, causing a major change in the olfaction of mosquitoes. The transmission of the disease is in part due to the parasite Plasmodium falciparum life-cycle that takes place within the mosquito vector. It is also believed that the parasite might manipulate the mosquito's behaviour to enhance its transmission. The various development stages (oocysts to sporozites) within the parasite play an integral role in increasing or decreasing the biting rate and thus the rate of contact of the mosquito with the humans, effectively leading to changes in the rate of transmission. However, differential attractiveness makes the testing of such a parameter difficult. The background variation leads to uncertainties that any observed difference is due to infection rather than to a difference in intrinsic attractiveness. At the same time, this variability of intrinsic attractiveness to mosquitoes could be accounted for with a comparison of two measures of the attractiveness of individuals; once when they have the parasite and the next one when they have cleared it.
It has been shown (Lacroix et al., 2005) that increased attractiveness is not due to an intrinsic attractiveness of gametocyte carriers but to the infection status associated with the presence of gametocytes. The exact reasons for this mechanism remains unexplained, but it is likely that the parasites change the infected individual’s breath or body odour, as these are involved in attracting mosquitoes at the distances involved in the experiment. Former gametocyte carriers also seemed to repel the mosquitoes in these tests. An explanation for this result could be based on the slight anaemia in previously infected children. The sense of this condition could have led them to children with a higher concentration of red blood cells. At the same time, however, it could be interpreted that the children previously infected with the asexual stage should also repel mosquitoes but this was not observed.
Compounds of Interest
Inspite of earlier studies (de Jong and Knol, 1995) showing CO2 as a potential attractant, it has been recently proved otherwise in independent studies in Wageningen Agricultural University and Kenya (Mukabana et al., 2004). Homogenous plumes of CO2 also attracted fewer mosquitoes as compared with skin odour (Dekker et al., 2001). CO2 has been reported as an attractant, repellent and neutral in studies carried out in the 60 and the 70s. Hence, the precise mechanisms involved in flight in CO2 plumes of differing structures needs further study at both sensory and behavioural levels.
Carboxylic acids have been identified in human sweat as well as in Limburger cheese, from which they are thought to be released as a result of the microbial activity of related bacterial strains. Sensory cells associated with the grooved peg sensillum of Aedes aegypti are known to be sensitive to lactic acid and short chain carboxylic acids. In a recent study conducted using tungsten electrodes inserted into the sensillum of the mosquitoes, high responses were noticed for 1-octen-3-ol and to short chain carboxylic acids at a load of 2500 μg with a stimulus duration of 2 s. It has also been observed that a part of the sensilla trichodea contains cells that respond with either excitation or inhibition and certain others show mixed behaviour. Dosages of the acids present play an important role, which can be justified by the fact that below a certain minimum amount of these compounds, the cells showing excitatory responses were inhibited. Higher concentrations lead to increased responses of these cells with respect to time (Meijerink et al., 1999).
Ammonia is emitted from the human skin as a microbial breakdown of excreted sweat products like urea and nitrogen-rich compounds. Preliminary analysis involving wind tunnels revealed a receptor neuron in the antennal grooved-peg sensilla sensitive to the odour of incubated sweat and unresponsive to fresh sweat, which indicated a strong attraction towards ammonia (Meijerink et al., 2001). Recent studies have shown strong electroantennogram responses by incubated sweat. Ammonia, being one of the major components of human sweat is thought to be one of the primary attractant for Anopheles gambiae. Ammonia sensitive receptor neurons have been discovered through EAG responses. However, their population in the grooved-peg sensilla is not widespread and only certain neurons have been specifically identified. Experiments with the sensilla trichodea have not yielded any significant results. This is in part due to the fact that carboxylic acids inhibit the sensilla trichodea associated receptor neurons. Field-testing is yet to yield results.
Other sweat components include indole, 3-Methyl-1-butanol, 6-methyl-5-hepten-2-one, geranyl acetone, 1-dodecanol, hexanedioic acid and certain amines among others. In general, the receptor neurons responded positively to these sweat borne components (Meijerink et al., 2001). However, it has been proved that although, sweat compounds do illicit responses from mosquitoes, all the sensilla are not equally attracted. Different dosages of each of these compounds also bring about varied responses (Meijerink et al., 2001). Incubated sweat might contain certain inhibitory compounds like carboxylic acids that might hamper the sensitization process of the sensilla trichodea. The fluid nature of the incubated sweat implies that it may have retained odours that are readily volatized from the surface of the human skin. These factors lead to anomalies in experimental results, for e.g., no responses were shown by the sweat sample, whereas, individual components recording strong responses. Overall, 3 sweat borne components indole, geranyl ketone and 6-methyl-5-hepten-2-one gave significant EAG responses, indicating that they might serve as kairomones.
Moisture is reported to influence mosquito behaviour since long, further substantiated by the fact that mosquitoes are frequently spotted in huge swarms near large reservoirs of water. In recent studies, grooved-peg receptor neurons in Aedes aegypti have been found to respond to water vapour. Some receptor neurons were even found to show a decreased response to stimuli lacking in water like ether and ethanol. Hygro-sensitive sensilla have 3 receptor neurons, a cold receptor, a moist receptor and a dry receptor. Moist receptors respond with excitation to sudden increases in humidity, while dry receptors respond in similar fashion to sudden decreases in humidity (Meijerink et al., 2001). It has also been shown that different transduction mechanisms are used for water vapour than for volatiles. Behavioural experiments conducted reveal that moist air is attractive to Anopheles gambiae. In a recent study (Dekker et al., 2001), Anopheles gambiae was caught in traps with only clean humidified air. The high sensitivity of Anopheles gambiae to humidity gradients might have induced trap entry in this species. However, similar information on Aedes aegypti is not known. In fact, since moisture has a huge significance in host selection, tests were incorporated to test the credibility of attraction of Limburger cheese (It contains 44% water), which eventually proved the contrary (Knols et al., 1996).
Inhibition
Inhibition of the intrinsic behaviour of the mosquitoes with respect to the location of a host sometimes takes place in its life cycle. Blood feeding is an important factor whose role has been effectively studied (Takken et al., 2001). Mated females too showed a markedly different feeding pattern whose primary reason has been attributed to the nutritional state of the male mate (Fernandez et al., 1995). Allomones in the human breath have been experimentally documented to be an effective inhibitor (Mukabana et al., 2004).
Dosage Responses
Olfactory mechanisms work only if they are provided with the right concentration of the chemicals, i.e., mosquitoes are attracted only if the concerned stimulus is present in large enough concentrations. Sometimes nano-grams of a sample are enough for significant excitation. Different species respond with varied excitation to different chemicals as was verified in a recent study (Vanden Broek et al., 1999). Hence, it becomes increasingly important to realize that the relative quantities of the various substances in the odour blends emanated by men and animals may determine host recognition.
Modes of Measurement
Experimental procedures involved in carrying out subtle observations require high end equipment that is capital intensive and not the most convenient to maintain. However, in the study of mosquitoes, certain cheap albeit highly effective instruments have been used and sometimes developed by the researchers themselves to conduct their experiments. The following section summarizes the major aspects of some of these instruments and analysis of the data measured by them.
Olfactometers are used to test how compelling single compounds or mixtures are towards attracting these insects. Construction aspects of these instruments have been described in an earlier study (Enserink, 2002). These studies for the attraction towards blends are generally carried out for a carefully bred sample of mosquitoes. The breeding of mosquitoes is carried out in a very controlled manner as far as atmospheric conditions is concerned and depend on the experiments that are to be done. An example of the type of conditions required can also be found in a recent work (Takken et al., 2001) among the many similar studies carried out in the past..
In certain attractant baited traps, chemical attractants are usually impregnated or encased in a rubber or plastic lure that slowly releases the active component(s) over a period of several days or weeks in these traps to monitor pest populations or for simple data handling. Basic advantages include high sensitivity of these traps, ability to capture insects at very low densities and extreme specificity. These can be effectively used to indicate the first emergence or peak flight activity of a pest species in a given area. However, there are certain external problems like ambient temperature, wind speed, direction and the time of the day during which it is used that cause discrepancies in the data collected (Weinzierl et al., 1995). Human beings are also being used as bait to collect mosquitoes for e.g., the Mbita Trap wherein they are used as bait but are not bitten. A recent study (Langanier et al., 2003) evaluated the effectiveness of these often-used traps and it was found that the Mbita trap collection was poor and unreliable as compared to human landing collections. However, this study being local in nature cannot be used for extrapolation to other areas since basic nature of mosquitoes (anthropophilic or zoophilic) was not tested.
Measurement of certain human skin emanations has assumed considerable importance in this field and many innovative techniques have sprung up. Electronic measurements are carried out through specially designed electronic noses that are exposed to the headspace of the compounds. The variation in steady state frequencies is used for comparison between different concentrations. The construction and working of these electronic noses is described in a recent study (Di Natale et al., 2000). Measurement of compounds emanated through glass beads has been improvised by Ulrich R. Bernier (Bernier et al., 1999). The main advantage of this technique is that the measurements correspond to compounds in their actual state that specifically cause the activation or host-seeking behaviour rather than those that are dissolved in solution. The collected compounds are then analysed in a chromatograph. An interesting procedure developed in Japan (Shirai et al., 2000), prevents human bites in spite of using human bait by cutting of the proboscis of the female mosquitoes. The positive results indicate that experiments with amputated mosquitoes show the same results as compared to normal mosquitoes. Electronic measurements of the sensilla are also carried out for obtaining information on nerve endings excited by compounds. These electrodes are generally inserted into the thorax or the eye of the mosquito and then moved around to catch any electrophysiological activity. These tests are generally carried out for certain doses of compounds (Meijerink et al., 1999).
Small-scale spatial variation and temporal heterogeneity in mosquito densities can have important consequences for disease transmission, but the extreme variation which is observed in populations of malaria vectors makes it difficult to obtain good predictions of densities for short time periods over limited areas. Preliminary procedures involve a search for the location where the sampling can be done. Once, this is done, a rough sketch of the place is made. GPS or other navigational systems are frequently used for this purpose. A population-check of the selected location is carried out including those of cattle and the general work practices of the human population are noted down. Sampling is done in each house, preferably and continuously for a fixed number of nights. Models are then used to simulate the factors affecting mosquito densities (Smith et al., 1995). The crucial factor in these studies is the model fitting, i.e., how close the models are in estimating the actual density of the vectors. Construction aspects of houses prove to be an important determinant in the sampling procedure of certain species. Houses without windows would have higher densities of mosquitoes; however, results have been reported to be in contradiction of the same and can be attributed to geographical factors. Raised houses also reduce mosquito bites (Charlwood et al., 2003). Distance of human populations from larval habitats also has been identified as a critical factor for mosquito densities (Minakawa et al., 2002). Biting of Anopheles gambiae took place primarily outside at ground level, on the verandahs and inside houses built on stilts. Houses built on stilts had half the number of mosquitoes than the ones built at ground level. However, not all species of mosquitoes could be differentiated in this manner. These studies can only help us in a local control of mosquito vectors. Future studies on such construction aspects can give us further insight into the flight of mosquitoes.
Genomic and Molecular Studies
Apart from physiological experiments, the main thrust in insect research revolves around genomic studies and measurement, where by, receptors and certain proteins are identified and studied. Understanding the molecular basis for human host recognition by mosquitoes can allow us to devise new rational approaches to interfere with this process. A systematic description of the exact process of olfaction at the molecular level is provided in Xu et al. (2003). Mosquitoes detect volatile chemical signals using olfactory neurons located on their antenna and maxillary palps, and tastants are detected by contact chemoreceptors distributed broadly over their body surface. These chemosensory neurons reside within segregated compartments called sensilla. Each sensillum is hair-like, hollow, fluid-filled structure containing the dendrites of olfactory or gustatory neurons bathed in sensillum lymph. Odorants enter the sensilla through pores or grooves in the wall, dissolve in the sensillum lymph and activate olfactory neurons. Sensillum lymph is secreted by non-neuronal support cells and contains a variety of secreted proteins. One of these proteins are the Odorant Binding Proteins (OBPs) secreted by the support cells into the sensillum lymph. A detailed description of the exact molecular process involving all the proteins is provided in a recent study (Zwiebel and Takken, 2004).
Identification of Receptors
A recent research (Hill et al., 2002) used bioinformatic approaches to identify a total of 276 G protein-coupled receptors (GPCRs) from the Anopheles gambiae genome. GPCRs comprise the largest category of proteins in animal genomes and are involved in chemo- and photoreception and a variety of processes. These receptors link ligands and downstream effectors as well as integrate and amplify other cellular signals. Seventy nine of these were characterized as odorant receptors (GPRors). The identification of these gene clusters and their likely role in mediating host preference and other processes central to vectorial capacity suggests that these studies will likely contribute to the development of a new generation of mosquito attractants. In an independent study (Xu et al., 2003), 57 candidate odorant binding protein (OBP) genes were identified in the Anopheles gambiae genome. 29 classical OBP genes were identified that were structurally similar to OBP genes identified in other species. Similar studies have been carried out by other research groups (Fox et al., 2001; 2002) for further analysis of already identified genes. Receptor AgOr1, a protein, which responds to the human sweat, has also been identified (Hallem et al., 2004). This receptor responds strongly to 4-methylphenol. Another protein AgOr2 has been shown to respond to 2-methylphenol. Both these compounds have been identified as components of human sweat.
Location of Receptors
The majority of GPRors (64 of 79) show expression only in olfactory tissues whereas 4 others show additional expression in legs. No candidate GPR or-specific products were observed with the head/proboscis. In another study (Melo et al., 2004), candidate odorant recptors were mostly found in the sensilla trichodea of the Aedes aegypti. A detailed location analysis of the specific receptors can be found in the same study.
Impact on Attraction
After a blood meal, a down-regulation of several broad classes of Anopheles gambiae genes was observed which resulted in a dramatic slowdown on host hunting. The AgOr1 (one of the Anopheles gambiae odorant receptors) has been largely held responsible for the same, according to a study (Fox et al., 2001). These studies have led to believe that odour-specific components of the mosquito chemosensory system that are tuned to host-specific molecules, such as those described above are potential targets for novel inhibitors that could be applied to the skin to disguise human hosts.
Consumer Applications
The aim of any research has to be the application of the same and to make a better, safer world. The search for the perfect attractant blend for mosquitoes can lead towards an alternative to the insecticides and repellants that we use. A US company American Biophysics Corporation has already jumped into the bandwagon by manufacturing and selling the Mosquito MagnetTM. It attracts mosquitoes by emitting a compound called 1-octen-3-ol, as well as heat, CO2 and water vapour. It costs anywhere from US $200 to US $2000. Two university spin-off companies were set up at the beginning of 2003 to update, refine and market the new capturing devices efficiently. But, it is the high prices that are not attracting it to the common man. The key, however, is not to build such expensive gadgets. Willem Takken of Wageningen Agricultural University, The Netherlands, is of the opinion that a simple $1-2 trap that people in developing countries could affix to their doorposts to keep out the mosquitoes, thus preventing the spread of deadly diseases, could do the trick. An interesting gadget could be something like a blend that attracts mosquitoes attached to an electric net. Although, theoretically, it could work like a charm, a preliminary analysis has concluded that it attracts a messy mixture of insects, killing certain beneficial ones as well.
Summary, Conclusion and Future Research
Attraction studies began first in the 1950s. The use of insecticides brought a temporary halt to these studies in the 70 and 80s. However, when the harmful effects of these chemicals on humans were documented, research on the former received an impetus and has only gained in force and magnitude. The key would be to capture unwanted mosquitoes, without damaging the environment, when and where it is needed. Future research could involve a more detailed field analysis and the merging of various results from different parts of the world giving a more unified result that in turn, could prove more beneficial rather than having a local view of the study.
The search for the perfect attractant has hit a roadblock of sorts, considering the fact that not all species are equally attracted to all humans. Differential attractiveness plays a major role in such situations. Several compounds have been short-listed and tested but none have given positive results for all the species and it has become equally important to quantify the dosage responses seen in mosquito attraction. There is also a need to setup a good standard for experimentation that could aid in comparison and application of results worldwide. A blend has been conjured (Bernier et al., 1999) that attracts these pests in a good number. They are waiting for patents before they release the information.
Genomic aspects have a large role to play in this area with a more evolutionary aspect coming into play, provided ethical aspects are borne in mind. The need of the hour is the combination of the behavioural, molecular and physiological studies being conducted currently in this research field.
ACKNOWLEDGMENTS
Sincere gratitude goes out to Dr. Takken, Department of Entomology, Wageningen University, The Netherlands; Dr. Ulrich Bernier, United States Department of Agriculture - Agricultural Research Service, Gainesville, Florida, USA and Dr. Zwiebel, Department of Biological Sciences, Vanderbilt University, Nashville, Tennessee, USA for their prompt sharing of relevant research papers.
REFERENCES
- Bernier, U.R., M.M. Booth and R.A. Yost, 1999. Analysis of human skin emanations by gas chromatography/mass spectrometry. 1. Thermal desorption of attractants for the yellow fever mosquito (Aedes aegypti) from handled glass beads. Anal. Chem., 71: 1-7.
CrossRefDirect Link - Braks, M.A.H., R.A. Anderson and B.G.J. Knols, 1999. Infochemicals in mosquito host selection: Human skin microflora and Plasmodium parasites. Parasitol. Today, 15: 409-413.
Direct Link - Charlwood, J.D., J. Pinto, P.R. Ferrara, C.A. Sousa, C. Ferreira, V. Gil and V.E. de Rosario, 2003. Raised houses reduce mosquito bites. Malaria J., 2: 45-45.
Direct Link - Costantini, C., N. Sagnon, A.D. Torre, M. Diallo, J. Brady, G. Gibson and M. Coluzzi, 1998. Odor-Mediated host preferences of West African mosquitoes with particular reference to Malaria vectors. Am. J. Trop. Med. Hyg., 58: 56-63.
Direct Link - De Jong, R. and B.G.J. Knols, 1995. Olfactory responses of host-seeking Anopheles gambiaes Giles (Diptera: Culicidae). Acta Trop., 59: 333-335.
CrossRefDirect Link - Di Natale, C., A. Macagnano, R. Paolesse, E. Tarizzo, A. Mantini and A. DAmico, 2000. Human skin odour analysis by means of an electronic nose. Sensors Actuators, B: Chemical, 65: 216-219.
CrossRefDirect Link - Enserink, M., 2002. What mosquitoes want? Secrets of host attraction. Science, 298: 90-92.
PubMedDirect Link - Fernandez, N.M. and M.J. Klowden, 1995. Male Accessory Gland Substances Modify the Host-seeking Behaviour of Gravid Aedes aegypti Mosquitoes. J. Insect Physiol., 41: 965-970.
CrossRefDirect Link - Fox, A.N., R.J. Pitts, H.M. Robertson, J.R. Carlson and L.J. Zwiebel, 2001. Candidate odorant receptors from the malaria vector mosquito Anopheles gambiae and evidence of down-regulation in response to blood-feeding. PNAS, 98: 14693-14697.
PubMedDirect Link - Fox, A.N., R.J. Pitts and L.J. Zwiebel, 2002. A Cluster of Candidate Odorant Receptors from the Malaria Vector Mosquito, Anopheles gambiae. Chem. Senses, 27: 453-459.
Direct Link - Gomes, A.C., N.N. Silva, G.R.A.M. Marques and M. Brito, 2003. Host-feeding patterns of potential human disease vectors in the Paraiba Valley Region. State of Sao Paulo, Brazil. J. Vector Ecol., 28: 74-78.
Direct Link - Hallem, E.A., A.N. Fox, L.J. Zwiebel and J.R. Carlson, 2004. Mosquito receptor for human-sweat odorant. Nature, 427: 212-213.
CrossRefDirect Link - Hill, C.A., A.N. Fox, R.J. Pitts, L.B. Kent and P.L. Tan et al., 2002. G Protein-coupled Receptors in Anopheles gambiae. Sci., 298: 176-178.
CrossRefDirect Link - Knols, B.G.J. and R. De Jong, 1996. Limburger cheese as an attractant for the malaria mosquito Anopheles gambiae s.s. Parasitol. Today, 12: 159-161.
CrossRefDirect Link - Langanier, R., F.M. Randimby, V. Rajaonarivelo and V. Robert, 2003. Is the mbita trap a reliable tool for evaluating the density of anopheline vectors in the highlands of madagascar. Malaria J., 2: 42-42.
CrossRefDirect Link - Lindsay, S., J. Ansell, C. Selman, V. Cox, K. Hamilton and G. Walraven, 2000. Effect of pregnancy on exposure to malaria mosquitoes. Lancet, 355: 1972-1972.
Direct Link - Meijerink, J., M.A.H. Braks and Van J.J.A. Loon, 2001. Olfactory receptors on the antennae of the malaria mosquito Anopheles gambiae are sensitive to ammonia and other sweat-borne components. J. Insect Physiol., 47: 455-464.
PubMedDirect Link - Meijerink, J. and J.J.A. Van Loon, 1999. Sensitivities of antennal neurons of the malaria mosquito Anopheles gambiae to carboxylic acids. J. Insect Physiol., 45: 365-373.
CrossRefDirect Link - Melo, A.C.A., M. Rutzler, R.J. Pitts and L.J. Zwiebel, 2004. Identification of a Chemosensory Receptor from the Yellow Fever Mosquito, Aedes aegypti, that is highly conserved and expressed in olfactory and gustatory organs. Chem. Senses, 29: 403-410.
Direct Link - Minakawa, N., P. Seda and G. Yan, 2002. Influence of host and larval habitat distribution on the abundance of African malaria vectors in western Kenya. Am. J. Trop. Med. Hyg., 67: 32-38.
Direct Link - Mukabana, W.R., W. Takken, R. Coe and B.G.J. Knols, 2002. Host-specific cues cause differential attractiveness of Kenyan men to the malaria vector Anopheles gambiae. Malaria J., 1: 17-17.
Direct Link - Mukabana, W.R., W. Takken, G. Killeen and B.G.J. Knols, 2004. Allomonal effect of breath contributes to differential attractiveness of humans to the African malaria vector Anopheles gambiae. Malaria J., 3: 1-1.
Direct Link - Shirai, Y., K. Kamimura, T. Seki and M. Morohashi, 2000. Proboscis amputation facilitates the study of mosquito (Diptera: Culicidae) Attractants. Repellents and Host Preference. J. Med. Entomol., 37: 637-639.
Direct Link - Smith, T., J.D. Charlwood, W. Takken, M. Tanner and D.J. Spiegelhalter, 1995. Mapping the densities of malaria vectors within a single village. Acta Trop., 59: 1-18.
CrossRefDirect Link - Takken, W., J.J.A. van Loon and W. Adam, 2001. Inhibition of host-seeking response and olfactory responsiveness in Anopheles gambiae following blood feeding. J. Insect Physiol., 47: 303-310.
PubMedDirect Link - Van den Broek, I.V.F. and C.J. den Otter, 1999. Olfactory sensitivities of mosquitoes with different host preferences (Anopheles gambiae s.s., Anopheles arabiensis, Anopheles quadriannulatus, Anopheles m. atroparvus) to synthetic host odours. J. Insect Physiol., 45: 1001-1010.
Direct Link - Xu, P.X., L.J. Zwiebel and D.P. Smith, 2003. Identification of a distinct family of genes encoding atypical odorant-binding proteins in the malaria vector mosquito, Anopheles gambiae. Insect Mol. Biol., 12: 549-560.
PubMedDirect Link - Zwiebel, L.J. and W. Takken, 2004. Olfactory regulation of mosquito-host interactions. Insect Bio. Chem. Mol. Biol., 34: 645-652.
Direct Link