
Fernando Garci Tamayo
Departamento de Biolog�a, Facultad de Qu�mica,
Universidad Nacional Aut�noma de Mexico, Mexico D.F 0451 Mexico
Teresa Izquierdo
Departamento de Sistemas Biol�gicos,
Universidad Aut�noma Metropolitana-Xochimilco, Mexico
Guadalupe Reyes
Departamento de Biolog�a, Facultad de Qu�mica,
Universidad Nacional Aut�noma de Mexico, Mexico D.F 0451 Mexico
Patricia Elizalde
Departamento de Sistemas Biol�gicos,
Universidad Aut�noma Metropolitana-Xochimilco, Mexico
Elba Rojas
Departamento de Sistemas Biol�gicos,
Universidad Aut�noma Metropolitana-Xochimilco, Mexico
Emilio Rojas del Castillo
Departamento de Genetica y Toxicolog�a Ambiental,
Instituto de Investigaciones Biomedicas, UNAM, Mexico, D.F 0451 Mexico
Luis I. Terrazas
Uniclad de Biomedicina, Facultad de Estudios Supenores-lztacala, t_XAM,
Asian Network for Scientific InformatonTlalnepantla, Edo. de Mexico, 54090 Mexico
Journal of Medical Sciences
Year: 2005 | Volume: 5 | Issue: 3 | Page No.: 169-176







ABSTRACT
A water-soluble Polysaccharide Extract (PE) isolated from the pulp of the fruit bodies of a Mexican species of Tamarindus indica L. was characterized by thin-layer chromatography and HPLC. Fructose, glucose, arabinose and xylose were the major components of the hydrolyzed PE samples. Normal lymphocytes and several lineages of malignant cells were cultured in presence of different amounts of PE solution and their effects on cellular proliferation were assessed by tritiated thymidine uptake. After incubation for 48 h, a significant reduction on the thymidine uptake was observed when Con-A-stimulated mice’s splenocytes were incubated with tamarind PE. Furthermore, a similar reduction in proliferative activity was detected also in CTLL-2 and HeLa cell lines. A comparative control assay showed a more intense inhibition when similar amounts (5-100 μg mL-1) of cis-platinum was added to HeLa cells. The morphological examination of PE-treated HeLa cells revealed their transformation into multinucleated giant cells. The results suggest that polysaccharide extracts obtained from tamarind pulp may contain active inhibitors of in vitro cellular multiplication.
PDF Abstract XML References Citation
How to cite this article
Fernando Garci Tamayo, Teresa Izquierdo, Guadalupe Reyes, Patricia Elizalde, Elba Rojas, Emilio Rojas del Castillo and Luis I. Terrazas, 2005. Altered Proliferative Response of Normal and Malignant Cells Exposed to a
Polysaccharide Extract from Tamarindus indica Pulp. Journal of Medical Sciences, 5: 169-176.
DOI: 10.3923/jms.2005.169.176
URL: https://scialert.net/abstract/?doi=jms.2005.169.176
DOI: 10.3923/jms.2005.169.176
URL: https://scialert.net/abstract/?doi=jms.2005.169.176
INTRODUCTION
Although Tamarindus indica L. is a tree indigenous to India and South East Asia, it now grows naturally in other tropical and subtropical regions of the world where their fruit is popular. Tamarind seeds are frequently used as a soup seasoner and the pulp of the fruit is widely used for foods and beverages[1]. Tamarind seeds contain high levels of certain essential amino acids, thus making them a good candidate to supplement animal and human diets[2]. Moreover, diverse medicinal liquors made of the tamarind pulp are recommended in developing countries for their laxative, antiseptic, diuretic and anti-inflammatory effects[3]; they have also been showed to be beneficial in controlling fever[4].
The frequent use of tamarind fruit as food[5] and the traditional application of their crude extracts for medicinal purposes[6] have stimulated diverse studies concerning to its chemical composition. Studies of the biological activities of their extracts have shown diverse and fairly interesting activities. For example, the pulp extracts of T. indica can influence intestinal permeability by increasing the bioavailability of aspirin in healthy human volunteers[7] and extracts obtained from seeds was found to possess anti-fungal[8] and anti-oxidative activities[9]. Recently, a crude polysaccharide obtained from tamarind seeds had an in vitro inhibitory effect on both chemotaxis and proliferation of human peripheral blood cells[10]. The present study reported that a similar crude polysaccharide, obtained from the pulp fruit of a Mexican species of Tamarindus indica, inhibits the in vitro proliferative response of both normal and transformed cells.
MATERIALS AND METHODS
Isolation and characterization of the polysaccharide extract: The fruits were obtained in Cuautla, a village near 150 km to México City. In the Etnopharmacology Laboratory of the Universidad Autónoma Metropolitana, a Crude Polysaccharide (CP) from dried pulp (50 g) of the fruit of T. indica was extracted with water at 4°C according to methodology Sreelekha et al.[10]. The CP was filtrated, precipitated and collected by centrifugation. The pellet was dissolved in distilled water and dialyzed with agitation for 48 h against distilled water at 4°C. After centrifugation, the supernatant was collected and mixed with chloroform. Denatured proteins were removed from the water-chloroform interphase. The procedure was repeated until the interphase was clear. The aqueous layer containing the Polysaccharide Extract (PE) was collected, dialyzed against distilled water, lyophylized and redissolved (5 mg mL-1) in acetate buffer pH 7.0. Before its use, the solution was clarified by vacuum filtration through activated charcoal (Merck). The protein content of the PE was assayed by the Bradford method[11] and the component sugars of hydrolyzed samples were analyzed by chromatography[4].
Chromatography: Preliminary thin layer chromatography studies were performed with silica gel plates (Merck) impregnated in 0.02 M sodium acetate. Separation of the water soluble saccharides from the hydrolized extract of the tamarind pulp was also obtained in the Organic Chemistry Department of the Facultad de Química, UNAM, by HPLC, using a modular model (Waters Associate Inc., Milford, MA) with a high pressure pump model S10 and a Rheodyne injector connected to a 300x3.9 mm carbohydrate analysis steel column (Perkin-Elmer Corp, Norwalk, CO), with a 410 refraction index detector model. A 90-10% v/v acetonitrile-water mobile phase with a 1 mL min-1 flux was used. Standard solutions of xylose, arabinose, fructose, glucose and galactose were used as reference carbohydrates.
Cell sources: Cells used to test the anti-proliferative effect of PE were obtained from different sources. (a) Splenic lymphocytes were obtained from two-month old BALB/c healthy female mice, which were killed by cervical dislocation, having previously been anesthetized with ether. The cells from the spleen were dissociated in a cold Phosphate Buffered Solution (PBS) under sterile conditions; (b) CTLL-2 (an IL-2-dependent lymphoid cell line) was originally obtained from the American Type Culture Collection (Rockville, MD); (c) YAC-1 cell line derived from a virus-induced murine lymphoma was kindly provided by MC Nancy Mora (Immunology Dept, IIBM, UNAM) and (d) HeLa cell line was derived from an adenocarcinoma of human uterine cervix. All cells were washed three times before their use and then they were adjusted in RPMI 1640 medium (Hy Clone Lab, USA) supplemented with previously inactivated (56°C for 30 min) 10% Fetal Calf Serum (Flow Lab, USA), 10 mM L-glutamine (Microlab, Mexico, DF), 25 mM HEPES (Microlab, Mexico, DF), 100 U mL-1 penicillin (Microlab, Mexico, DF) and 100 μg mL-1 streptomycin (Microlab, Mexico, DF). All cultures and the inhibition of the proliferative assays were performed in the Biology Department of the Facultad de Química, UNAM.
Cell viability: Initial cell viability was measured by the addition of one volume of 0.4% trypan blue (Gibco BRL, USA) in PBS to 5 volumes of cells suspended in the culture medium. After incubation at 23°C for 5 min, the cells were examined by light microscopy for evidence of dye exclusion. In other experiments, final viability in HeLa cells was studied after incubating for 48 h with two different amounts (50 and 100 μg mL-1) of tamarindus polysaccharide extract.
Cell cultures: (a) Single cell suspensions of normal lymphocytes were obtained by disaggregating of the splenic tissue. The erythrocytes were eliminated by osmotic shock with NH4Cl solution and the remaining cells were washed three times in RPMI medium and adjusted to 3x106 cells mL-1. One hundred microlitres of splenic mononuclear cells were plated in triplicate, twice, in 96-well flat-bottomed plates (Costar, Cambridge, Mass.) and mixed with 100 μL of culture medium containing progressive 2x dilutions of an initial solution (100 μg mL-1) of tamarind PS. (b) CTLL-2 cell line was grown in RPMI-1640 and supplemented as mentioned above by adding 1 U mL-1 of recombinant murine IL-2 (rmIL-2) (Pharmingen, San Diego, CA). After three days of culturing, the CTLL-2 cells were washed with RPMI-1640, adjusted to 200x103 cells mL-1 and suspended in a culture medium containing 8 U mL-1 of IL-2. Aliquots of 100 μL were added in triplicate to the wells of a microculture plate which had previously received 100 μL of serial dilutions of tamarind PE. (c) YAC-1 and HeLa cell lines were also adjusted at 200x103 cells mL-1 in RPMI-1640 medium, without IL-2 and mixed with equal volumes of the PS dilutions. All cells were plated in a similar fashion and the cultures were performed in triplicate, with or without the polysaccharide extract. A control inhibitory assay was also performed by adding progressive 2x dilutions of a 100 μM cis-platinum solution to HeLa cell cultures.
Inhibition of the proliferative assay: The cell cultures with splenic lymphocytes were tested by mitogen stimulation using Concanavalin A (Con A) (Sigma, St. Louis, Mo.) according to the procedure previously described[12]. Con-A (2 μg mL-1) was solubilized and added (0.1 mL) to wells containing the culture medium and PE serial dilutions. The cells were incubated in a humidified atmosphere with 5% CO2 at 37°C for 72 h. Eighteen hours before culture ending, 0.5 μCi of [3H]-TdR (6.7 Ci/mM, New England Nuclear Corporation, Boston, Mass.) was added (in RPMI 1640 medium) to each well. In other experiments, the cell lines YAC-1, CTTL-2 and HeLa were cultured in the same medium without mitogen stimulation. These cells were mixed with serial dilutions of tamarind PE and incubated for only 48 h. In a control experimental assay, serial dilutions of 100 μM cis-platinum were added to HeLa cell samples only. Then, 0.5 μCi of [3H]-TdR was added to each well 8 hours before the culture ending. All cells were harvested onto fiberglass strips with a multiple cell harvester by using copious amounts of a saline solution. [3H]-TdR incorporation was measured by liquid scintillation spectroscopy. The results are expressed as a percentage of the inhibition of the cell proliferation as follows: 100/(cpm from control cells/cpm from tamarind-treated cells).
Morphology of PE-treated HeLa cells: The HeLa cells were seeded over glass coverslips and incubated for 48 h at 37°C in RPMI-1640 medium supplemented as mentioned above. Tamarind PE (50 or 100 μg mL-1) was added to cultures. After 48 h, coverslips with cells were fixed (80% ethanol, 10% sucrose). To study their gross morphology and chromatin structure, cells were stained with 2,4-diamino-2-phenyl-indole (DAPI) and visualized on a fluorescence microscope. This morphologic studies and the quantification of multinucleated cells were performed into Departamento de Genética y Toxicología Ambiental del Instituto de Investigaciones Biomédicas, UNAM.
Quantification of multinucleated and giant cells: Giant multinucleated DAPI-stained cells were identified by using a Nikon Labophot-2 microscope (Nikon, Garden City, NY). At least 10 areas were randomly selected for analysis under 400 x magnifications. An average of 1500 cells was examined per coverslip and the percentage of positive cells was calculated. The data presented are representative of three separate experiments and the cells were counted by two different technically trained persons.
RESULTS
Sugars identified on the polysaccharide extract from tamarind pulp: Chromatographic analysis of hydrolyzed samples from water soluble PE showed a constant mixture of nonvolatile sugars. Different kinds of sugars were detected after a thin layer chromatography. Hydrolyzed samples were analyzed by HPLC (Fig. 1), percentages of the major components obtained were as follows, fructose (36.15%), glucose-galactose (35.40%), arabinose (9.23%), xylose (3.32%) and non-identified sugars (15.88%). Protein content was only 13% according to the Bradford method (data not shown).
Effect of tamarind PE on Concanavalin A-stimulated lymphocytes: Spleen cells from healthy mice exposed to different doses of tamarind PE showed a significant reduction in 3H-thymidine incorporation compared with unexposed cells.
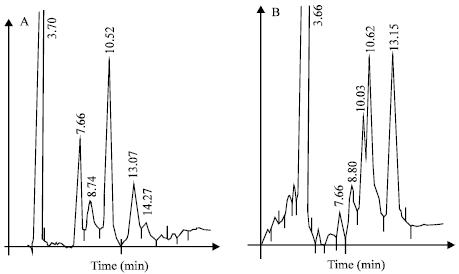 | |
| Fig. 1: | HPLC analysis of a hydrolyzed polysaccharide extract from the pulp fruit of Mexican species of Tamarindus indica. The first image (A) shows the successive elution of the components in the sugar mixture of the control sample, which were collected from a Carbohydrate Analysis column (300 x 3.9 mm) by using water: acetonitrile (10:90 v/v) as a mobile phase and a flow rate : 1.0 mL min-1; detector: RI. Graph shows first the void volume and after the successive resolution of xylose, arabinose, fructose, glucose and galactose. Image B shows the elution of the components in the hydrolyzed polysaccharide extract from tamarindus pulp fruit, corresponding to the void volume, xylose, arabinose, unidentified sugars, fructose and glucose-galactose |
A low dose of 25 μg mL-1 reduced until 70% the proliferative response of these cells (p<0.05), a higher concentration of PE showed a more intense inhibitory effect, whereas the lower dose (1.56 μg mL-1) produced no significant inhibition in the proliferation rate (Fig. 2). Thus, PE appears to act in a dose-dependent manner (r = 0.89).
Effect of tamarind PE on the multiplication of malignant cell lines: Since Con-A binds to carbohydrate structures expressed on the membranes of lymphocytes, we next attempt to rule out a possible interference of tamarind PE with the molecular target of Con-A. We use three different cell lines in the absence of the mitogenic stimulation by Con-A. When the tamarind PE was added to the culture medium containing CTLL-2 cells plus 8 μ mL-1 of rmIL-2, a dose-dependent inhibition in the cell proliferation was also observed. Tamarind PE at 25 μg mL-1 inhibited 60% of the 3H-thymidine uptake (Fig. 2), in this cell line. A similar effect was observed with HeLa cell cultures exposed to tamarind PE, where a progressive inhibition on the multiplication of this non-lymphoid tumor cell line was also detected after 48 h of incubation with increasing doses of tamarind PE (Fig. 2). PE concentration at 25 μg mL-1 inhibited 40% of the 3H-thymidine uptake by HeLa cells, whereas in the comparative control test, the [3H]-TdR uptake by these cells was inhibited 90% when 25 μg mL-1 of cis-platinum were added to the culture medium (Fig. 2).
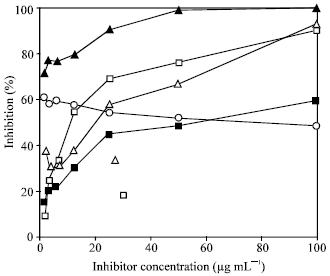 | |
| Fig. 2: | In vitro inhibition of nuclear uptake of 3H-thymidine in Con A-stimulated splenic T murine lymphocytes (□), CTLL-2 cells in presence of rmIL-2 (Δ), YAC-1 (○) and HeLa (■) cell lines, when they were exposed to progressive 2x dilutions (>100 μg mL-1) of the extract of tamarindus polysaccharide (PE). HeLa cells were also incubated with progressive dilutions of a 100 μM cis-platinum solution (▲). The inhibition of proliferation is expressed as the percent of reduction on the thymidine uptake in control cells without the PE. Results show the average of three separate experiments, each in triplicate |
Interestingly, non dose-dependent inhibitory effect was observed in cultures of the YAC-1 murine lymphoid cells when diverse amounts of tamarind PE were added to the culture, however, this cell line displayed a consistent susceptibility to tamarind PE exposure, since their proliferation was inhibited around 50-60 % in all doses tested (Fig. 2).
Effect of tamarind PE on the appearance of HeLa cells: The microscopic examination of the HeLa cells seeded on coverslips revealed morphological differences between HeLa cells exposed to tamarind PE and without PE. In the absence of PE, HeLa cells were abundant in the culture medium, having a uniform appearance and a relatively small size (Fig. 3A). In contrast, HeLa cells cultures exposed to tamarind PE exhibited an irregular size and shape (Fig. 3B). While some PE-treated HeLa cells were of comparable appearance to those cultured without PE, other cells displayed a cytoplasmic enlargement and an increase in the number of nuclei that each cell possess, sometimes more than 10 per cell (Fig. 3C and D), which were often grouped in the central area of the cells while at the periphery the cytoplasm was broadly spread out in thin lamellas with developed pseudopods. Between 200 and 250 out of 1500 HeLa cells were recorded as multinucleated-giant cells when 50 μg mL-1 of PE were added to the cultures versus 20-25 multinucleated-giant cells observed in control cultures without PE (Fig. 4). A significant proportion (20%) of the HeLa cells appeared dead after their incubation for 24 h with 50 μg mL-1 of tamarind PE, by contrast only 5% of cells dead when they were incubated in the culture medium without PE. However, the number of multinucleated cells was significantly elevated after they were exposed to 50 and 100 μg mL-1 of tamarind PE (Fig. 4).
DISCUSSION
Diverse polysaccharides isolated from both vegetal and bacterial sources have shown antiproliferative and antitumor activities[13,14]. Most of them indeed exhibited these effects through the activation of the host’s immune system. For example, macrophages and cytotoxic T lymphocytes increase their anti-tumoral activities when fruit polysaccharides are orally administered in C3H mice previously implanted with carcinoma tumor cells[15]. The in vivo and in vitro murine phagocytic function have been stimulated by polysaccharide fractions obtained from the marine alga Porphyra yezoensis[16], the polysaccharide fractions from other Japanese plant increase the anti-Sheep Red Blood Cells (SRBC) antibody response in normal BALB/c mice[17] and lectins from tubers of plants belonging to the Araceae family are potent mitogens to human peripheral blood lymphocytes[18]. Moreover, polysaccharides isolated from Angelica gigas Nakai stimulate the response of B lymphocytes by increasing the expression of IL-2, IL-4, IL-6 and IFNγ[19] and some protein-bound mushroom polysaccharides increase the production of TH1-type cytokines[20].
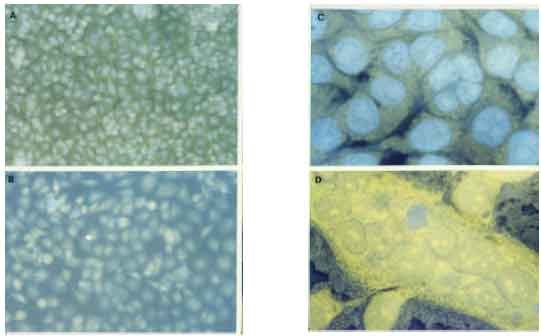 | |
| Fig. 3: | Diverse morphological abnormalities in HeLa cells were observed after their incubation for 48 h in culture medium with 50 or 100 μg mL-1 of the tamarind polysaccharide extract. (A) HeLa cells culture without PE (x200). (B) HeLa cells exposed to 100 μg mL-1 of PE after 48 h of culture (x200). (C) Several nuclei can be observed in HeLa cells incubated with PE (50 μg mL-1) during 48 h (x400). (D) Amplification of one HeLa cell with more than 10 nuclei after 48 h exposure to 50 μg mL-1 of tamarind PE (x1000) |
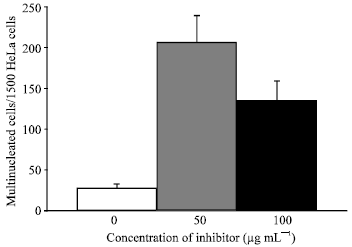 | |
| Fig. 4: | A high number of HeLa cells were identified as multinucleated cells after they were exposed during 48 h to different doses of tamarind PE (μg mL-1). Bars express the mean number±standard deviation of multinucleated cells, 1500 HeLa cells were counted per slide |
In other experiments, polysaccharide fractions derived from fungal samples[21] and dried safflower petals[22] can also exert an immunostimulatory effect by increasing the synthesis of pro-inflammatory cytokines.
However, not all vegetal, fungal or bacterial polysaccharides have the immuno-stimulant actions described above[13-22]. The arabinomannan exocellular polysaccharide from Mycobacterium tuberculosis inhibits the functions of human lymphocytes[23] and a cytotoxic principle isolated from Tamarindus indica fruits inhibits the development of sea urchin embryo cells[24]. Extracts from Tamarindus seeds exert an in vitro antiproliferative effect on human lymphocytes[10] and some polysaccharides isolated from fungal samples have antiproliferative activity on experimental transplanted tumors[25].
Here evidences indicating that a crude polysaccharide extracts from the pulp fruit from a Mexican species of T. indica also induces a significant inhibition of the cellular division of murine lymphocytes and also of tumor cell lines. In a previous study on the in vitro inhibition of the proliferative response of human lymphocytes[10] a blocking effect on the mitotic activity of phytohemagglutinin was proposed to explain the inhibitory activity of some polysaccharides from seeds extracts of T. indica. In contrast, our results have shown that the polysaccharides from the pulp of tamarindus fruit have a similar inhibitory effect on a cell line derived from a murine lymphoma (CTLL-2), whose in vitro multiplication is not induced by mitogenic stimulation. This discrepancy can be afforded if we considered that Sreelekha et al.[10] used the polysaccharide extracts from the seeds of tamarindus fruit while we utilized polysaccharides from the pulp of the fruit bodies of T. indica, which suggest that the composition of sugars could be different between the two extracts, besides a possible different target for their biological activity could also explain these apparent discrepancies. On the other hand, they assessed the antiproliferative response in normal human lymphocytes while we did it in four different types of cells.
It is also observed that the tamarindus polysaccharide extract was able to induce the transformation of HeLa cells into giant multinucleated cells. Although the formation of giant cells occurs spontaneously in cultures of HeLa cells, here it seems to be enhanced by the exposure to tamarindus PE. The biological significance of these tamarind PE-induced transformations remains unknown. However, several anti-neoplastic agents have been reported to induce the formation of cells with two or more nuclei both in vivo and in vitro assay[26]. Sodhi[27] has claimed that cis-platinun induces the transformation of Sarcoma-180 cells into binucleated and multinucleated giant cells by inhibiting cytokinesis. Another drug such as bleomycin also induces the formation of giant polynucleated cells by cell fusion or by mitosis without cell division[28]. Giant HeLa cells have been also observed after their incubation with an etoposide[29]. Different mechanisms such as the disruption of both tubulin polymerization and actin/myosin polymerization[30] or abnormal expression of H-Ras[31] could be involved in the generation of multinucleated cells and their catastrophic death by arresting the cytokinesis or the mitotic process. Although HeLa cells incubated in the presence of the tamarindus polysaccharide also showed the morphological changes similar to those observed in the induced mitotic death, whether tamarindus PE induces similar mechanisms such as described above is unknown and were out of scope in the present study.
It is concluded that water-soluble polysaccharide extracts obtained from pulp fruit of a Mexican species of T. indica can induce a dose-dependent inhibition of the in vitro proliferation of both Con A-stimulated lymphocytes from normal mice and also from tumor cell lines. Furthermore, when the same tamarindus polysaccharide extract was added to HeLa cells cultures, an elevated number of giant multinucleated cells were observed. Further experiments are needed, however, to identify and characterize the specific sugars derived from the tamarindus fruit that may be associated with this antiproliferative effect, as well as to decipher mechanism involved in inhibiting the cell proliferation.
ACKNOWLEDGEMENTS
The authors wish to acknowledge Alma Ibarra and Joaquín Velázquez (Laboratorio de Etnofarmacología, UAM-Xochimilco) for their technical assistance, Joseph Doshner (Laboratorio de Idiomas, FQ, UNAM) for revising the English text and John W. Hadden (Cold Spring Harbor, NY) for the critical reading and comments on the manuscript.
REFERENCES
- Ishola, M.M., E.B. Agbaji and A.S. Agbaji, 1990. A chemical study of Tamarindus indica (Tsamiya) fruits grown in Nigeria. J. Sci. Food Agric., 51: 141-143.
CrossRefDirect Link - Rimbau, V., C. Cerdan, R. Vila and J. Iglesias, 1999. Antiinflammatory activity of some extracts from plants used in the traditional medicine of North-African countries. Phytother. Res., 13: 128-132.
PubMedDirect Link - Tsuda, T., M. Watanabe, K. Ohshima, A. Yamamoto, S. Kawakishi and T. Osawa, 1994. Antioxidative components isolated from the seed of tamarind (Tamarindus indica L.). J. Agric. Food Chem., 42: 2671-2674.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Yoshizawa, Y., A. Ametani, J. Tsunehiro, K. Nomura, M. Itoh, F. Fukui and S. Kaminogawa, 1995. Macrophage stimulation activity of the polysaccharide fraction from a marine alga (Prophyra yezoensis): Structure-function relationships and improved solubility. Biosci. Biotechnol. Biochem., 59: 1933-1937.
- Han, S.B., Y.H. Kim, C.W. Lee, S.M. Park and H.Y. Lee et al., 1998. Characteristic immunostimulation by angelan isolated from Angelica gigas Nakai. Immunopharmacology, 40: 39-48.
CrossRefPubMedDirect Link - Ng, T.B., 1998. A review of research on the protein-bound polysaccharide (polysaccharopeptide, PSP) from mushroom Coriolus versicolor (Basidiomycetes: Polyporaceae). Gen. Pharmacol., 30: 1-4.
Direct Link - Zhuang, C., H. Itoh, T. Mizuno and H. Ito, 1995. Antitumor active fucoidan from the brown seaweed, Umitoranoo (Sargassum thunbergii). Biosci. Biotechnol. Biochem., 59: 563-567.
CrossRefPubMedDirect Link