
M.W. Harding
Crop Diversification Centre South, Alberta Agriculture and Rural Development, 301 Horticultural Station Road East, Brooks, AB, T1R 1E6, Canada
N. Butler
Innovotech Inc. 101-2011 94 Street, Edmonton, AB, T6N 1H1, Canada
W. Dmytriw
Alberta Innovates Technology Futures, P.O. Box 4000, Vegreville, AB, T9C 1T4, Canadaz
S. Rajput
Alberta Innovates Technology Futures, P.O. Box 4000, Vegreville, AB, T9C 1T4, Canadaz
LiveDNA: 91.10748
D.A. Burke
Crop Diversification Centre South, Alberta Agriculture and Rural Development, 301 Horticultural Station Road East, Brooks, AB, T1R 1E6, Canada
R.J. Howard
Crop Diversification Centre South, Alberta Agriculture and Rural Development, 301 Horticultural Station Road East, Brooks, AB, T1R 1E6, Canada
RJH Ag Research Solutions Ltd., P.O. Box 1456, Brooks, AB, T1R 1C3, Canada
Research Journal of Microbiology
Year: 2017 | Volume: 12 | Issue: 1 | Page No.: 20-32







ABSTRACT
Background and Objective: Microorganisms can colonize or contaminate fruits and vegetables at virtually any point during production, distribution, processing, packaging or food preparation. Microbial contamination of fruits and vegetables can cause food spoilage and human infections. This study was undertaken to collect, isolate and identify commonly occurring food spoilage microorganisms from a wide variety of fresh, direct-market fruits and vegetable in Southern Alberta and to test for the presence of the food-borne pathogen Listeria monocytogenes on a few representative farm operation surfaces. Methodology: The naturally-occurring microorganisms present on produce were amplified by incubation in humid chambers. Microorganism cultures were obtained by aseptically isolating from produce surfaces and sub-culturing on agar petri dishes. Results: Approximately 950 microbial isolates, both bacteria and fungi were collected from fresh produce. Identities of 80 selected isolates were confirmed using molecular analysis. A number of well-known plant pathogenic taxa were identified along with a few species that have not previously been reported to act as spoilage organisms, namely Isaria farinosus, Stenotrophomonas maltophilia, Rahnella aquatilis and Acinetobacter calcoaceticus (syn. Micrococcus calcoaceticus ). Listeria monocytogenes was not detected at any of the Southern Alberta farms sampled. Conclusion: These results indicate that a wide array of microorganisms are capable of causing food spoilage and some have not yet been documented or characterized.
PDF Abstract XML References Citation
Received: March 10, 2016;
Accepted: June 20, 2016;
Published: December 15, 2016
How to cite this article
M.W. Harding, N. Butler, W. Dmytriw, S. Rajput, D.A. Burke and R.J. Howard, 2017. Characterization of Microorganisms from Fresh Produce in Alberta, Canada Reveals Novel Food-spoilage Fungi. Research Journal of Microbiology, 12: 20-32.
URL: https://scialert.net/abstract/?doi=jm.2017.20.32
URL: https://scialert.net/abstract/?doi=jm.2017.20.32
INTRODUCTION
Large diverse communities of microorganisms can contaminate fruits and vegetables at any point throughout the production system from farm to fork. The size and diversity of these microbial communities are influenced by the plant, from which the produce originated1 the operational production and handling systems for fresh produce, the storage conditions2 and the interplay between pathogenic and non-pathogenic microorganisms within the community1,3. The rate of food spoilage will be governed in part by respiration rates and autolytic processes within the plant tissues. External factors, such as environmental conditions, activities of spoilage microflora and the resulting will also affect the shelf life of fresh produce. Microflora on fresh produce are a major factor in spoilage, but can also impact gastrointestinal health of consumers4,5 and may present risks of food-borne human illness. Therefore, two major problems can be ascribed to microbial contamination of fruits and vegetables, namely post-harvest food spoilage and human infections caused by pathogenic bacteria, viruses or parasites2,6. Ingestion of food containing non-pathogenic microorganisms can also affect microflora of the human gut by impairing function leading to poor gastrointestinal health or result in immune reactions and development of allergies7. Thus, characterizing the microbial communities associated with fresh produce is important to understand food preservation methods and impacts for food safety, especially in cases, where produce are ingested without cooking or end-user processing to inactivate or eradicate them.
A number of studies have been done to characterize the microbial communities associated with fresh produce1-3,8-17. These studies have revealed that microorganisms from diverse taxa are present on surfaces of fruits and vegetables. For example, Leff and Fierer2 identified hundreds of families of bacteria from 5 different phyla (Actinobacteria, Bacteriodetes, Firmicutes, Protobacteria and TM7) on 7 different types of produce using barcoded pyrosequencing of the 16S rRNA gene. However, these studies did not characterize the food spoilage potential of each culturable isolate by fulfilling Koch’s postulates.
Another important element of microbial colonization of food surfaces is their tendency to grow within extracellular polymeric substances as part of structured, surface-associated communities called biofilms18-20. The microorganisms that cause spoilage are no exception and are capable of forming biofilms15. The significance of the extracellular matrix produced by microbes is that it may afford protected sites for food-borne pathogens, including bacteria, viruses and parasites that cause diseases in humans. The biofilm matrix is thought to aid in their attachment, colonization and survival21-24. Considering the importance of microbial biofilms in plant disease and food spoilage, characterizations of biofilms on the surfaces of plants, including fruits and vegetables are limited15,25,26. It is assumed that all spoilage microorganisms form robust biofilms on the surfaces of the plant tissues they colonize, but in most cases, this has not been documented. Furthermore, if biofilms are formed on surfaces of fruits and vegetables by non-pathogenic or food-spoilage microflora, it is not known to what extent these biofilms actually reduce food safety by harbouring food-borne pathogens.
Here, it is reported the collection, isolation and identification of food spoilage microorganisms from surfaces of fresh produce obtained from farmer’s markets in Southern Alberta. Koch’s postulates were used to determine, which of the isolates were capable of causing spoilage. Visualizations of aggregates or biofilms, formed by microorganisms on produce surfaces were obtained via scanning electron microscopy. Testing for the presence of Listeria monocytogenes27,28 on operational farm surfaces was also performed. This pathogen was selected because it is a commonly occurring food-borne pathogen on fresh produce. Additionally, another concurrent survey of fresh produce was underway that tested for the presence of the other major food-borne pathogens E. coli, Salmonella spp., E. coli O157:H7, Campylobacter spp. and Cryptosporidium spp.29.
MATERIALS AND METHODS
Sampling of fresh fruits and vegetables: Ten farmers markets in nine different municipalities (Cardston County, County of Newell, County of stettler, Cypress County, Lethbridge County, Municipal district of Taber, Red deer County, Special area No. 3, Starland County) in Southern Alberta were visited (Fig. 1). The type of produce obtained was dependent upon seasonality and availability, but focussed on the sampling of carrots, lettuce, onions, spinach, tomatoes and strawberries. Each of the 10 markets were visisted twice during the marketing season. Produce was collected in the same condition as vendors sold to all customers and immediately stored in separate containers to avoid cross-contamination of surface-associated microorganisms during handling and storage.
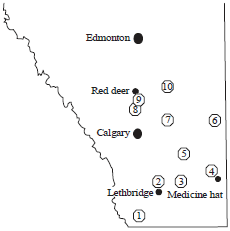 | |
| Fig. 1: | A map of the Southern half of Alberta marking locations of farms and farmer’s markets, where fresh produce samples were collected in 2007, 1: Cardston County, 2: Lethbridge County, 3: Municipal district of Taber, 4: Cypress County, 5: County of Newell, 6: Special area No. 3, 7: Starland County, 8-9: Red deer County and 10: County of Stettler. Environmental sampling collection sites were located at farms in 2: Lethbridge County, 3: M.D. of Taber, 5: County of Newell, 8-9: Red deer County and 10: County of stettler |
Samples were returned to the laboratory in coolers packed with ice and stored at 5°C for 1-6 days until isolations were performed. In total, 26 vegetables (Brussels sprouts, beans, beets, broccoli, cabbage, carrot, cauliflower, celery, corn, cucumber, garlic, green onion, leeks, lettuce, onion, parsnip, peas, peppers, potato, pumpkin, radish, spinach, squash, tomato, turnip and zucchini) and 7 fruits (black currant, cantaloupe, raspberry, sour cherry, saskatoon berry, strawberry and watermelon) were collected.
Environmental sampling of vegetable farms: Five farms were visited and environmental sampling performed (Fig. 1). Environmental sampling at the five farms was performed once. Samples at each site were collected from various zones (grading, packaging, storage and loading) on air-exposed agar petri dishes by removing the lids and exposing to the air for 20-25 min. Sampling sites were selected based on 3 criteria, (1) Site within the zone with the greatest amount of foot or vehicle traffic, (2) Site within the zone, where produce makes direct contact (or nearest surface to direct contact), (3) Site within the zone with abundant air ciruculation. All growth media used were Acumedia (Acumedia Manufacturers Inc., Lansing, MI) brand unless otherwise indicated. One Nutrient Agar (NA) and one Potato Dextrose Agar (PDA) dish were exposed in each farm zone sampled. After exposure, the lids were replaced and dishes were stored at room temperature (20-22°C) for at least 3 days or until sufficient microbial growth was present to allow sub-culturing of individual colonies. Each isolate was aseptically transferred to fresh agar until purified cultures were obtained. Each isolate was then transferred to agar slants for storage at 5°C.
Samples were also collected from hard surfaces using pre-hydrated sterile sponges (Qualicum Scientific Ltd., Nepean, ON). Surfaces of production, handling, grading and packaging equipment, as well as storage and transportation surfaces were sampled as follows, the seal on the sample bag was broken and the pre-hydrated, sterile sponge removed. An area of at least 1 m2 was firmly wiped and the sponge was immediately returned to the sample bag, labelled and stored in a cooler on ice or at 5°C, until microbial isolations were performed. Wiping was performed in duplicate. For recovery of microorganisms, the 1st set of sponges were covered in approximately 20 mL sterile phosphate-buffered saline and then aseptically hand massaged for 2 min after, which a sample of the liquid was streaked undiluted onto NA and PDA using a sterile loop. Inoculated agar plates were incubated at room temperature (20-22°C) for at least 3 days or until microbial growth was present and sub-culturing for isolation could be done. Each isolate was then transferred to agar slants for storage at 5°C.
The 2nd set of environmental sponges was tested for the presence of Listeria monocytogenes. Detection of L. monocytogenes in the environmental samples were performed as follows: To each sponge, 100 mL of demi-fraser broth was added and stomached for 30 sec. The homogenized samples were incubated at 30°C for 24 h. After incubation, the samples were mixed and 0.1 mL was transferred to 9.9 mL of MOPS-buffered Listeria enrichment broth. The inoculated broths were incubated at 35°C for 22 h. Positive, negative and sterility controls were included with each batch of samples tested. The enriched samples were tested using the BAX® System PCR assay (Du Pont Qualicon, Germany). Briefly, 5 μL of the enriched sample was transferred to lysis buffer, which was subsequently heated to 55°C for 60 min and then 95°C for 10 min. The lysate was then cooled and 50 μL transferred to a PCR reaction tube (BAX® system PCR assay for Listeria monocytogenes-D11000157). The inoculated reaction tubes were loaded in the BAX® system cycler/detector and the cycle run was performed as per the manufacturer’s recommendations.
Isolation of microorganisms from produce: All media used were Acumedia (Acumedia Manufacturers Inc., Lansing, MI) brand unless otherwise indicated. Humid chambers were prepared for each sample by placing non-sterile paper towels, 2-3 layers thick, moistened with tap water, in clean clear 8-10 lb plastic bags. Each sample were placed inside individual humid chambers, which were filled with air using a hand-held air pump and sealed with an elastic band. This was done to encourage or accelerate the growth of microorganisms on the samples. Humid chamber preparations were done inside a biosafety cabinet to prevent contamination or release of microorganisms. Moist chambers were incubated at room temperature (20-22°C) and maintained 85 to near-100% relative humidity. Produce samples were removed from humid chambers when spoilage was visible on the sample, ranging from 2-14 days incubation depending on the sample. Microorganisms visible on the decaying produce were aseptically transferred to agar plates. Suspected bacterial (non-filamentous) growth was plated on NA and suspected fungal growth (filamentous) was plated to PDA. Isolates were sub-cultured until pure and transferred to agar slants for storage at 5°C.
The bacterial isolates were grouped based on colony appearance and the fungal isolates were grouped based on colony appearance and spore morphology. One or more representative isolate(s) from each category were selected for DNA extraction, PCR amplification and sequencing.
Bacterial genomic DNA extraction and amplification: One milliliter suspensions from each of 40 fresh cultures of isolated colonies were made in sterile deionized water (DI water) and used for DNA extraction using Qiagen’s DNeasy Blood and Tissue Kit (Cat# 69504) following manufacturer’s protocols optimized for bacteria. Genomic DNA elution from the final spin column was eluted with 100 μL sterile DI water instead of manufacturer’s buffer to enhance sequence analysis. Quality and quantity of DNA were evaluated and adjusted by absorbance spectroscopy at 260 and 280 nm (NanoDrop Instruments).
Polymerase Chain Reaction (PCR) amplification of the 16S rDNA gene region was done using one of two primer sets: (1) P16S F/R primers P16SF, 5’-AGA GTT TGA TCC TGG CTC AG-3’, P16SR, 5’-GGT TAC CTT GTT ACG ACT T-3’30 and (2) 264F/1078R primers 264F, 5’-GAT GAT CAG CCA CAT TGG GAC-3’; 1078R, 5’-CCC AAC ATC TCA CGA CAC GAG-3’31. These primer sets amplified 1.5 kb (P 16S) and ~750 bp (264F/1078R) fragments by PCR done in a 50 μL volume containing 5 μL 10x reaction buffer, 2.5 μL MgCl2 (25 mM), 1 μL TAQ DNA polymerase recombinant (Cat# 10342020, Invitrogen), 35.5 μL sterile DI water, 1 μL dNTP’s (25 μM), 2 μL each primer (10 pmol) and 1 μL genomic DNA (1-50 ng μL–1). Conditions for PCR amplification were: 5 min at 95°C, followed by 35 cycles of 45 sec at 94°C, 45 sec at 55°C and 45 sec at 72°C and ending with 5 min at 72°C. After PCR amplification, 5 μL of product was separated in a 1% agarose gel in 1×tris-acetate-EDTA (TAE) buffer. Bands corresponding to each primer set were cut out and gel extracted using the Qiagen QIAquick Gel Extraction Kit (Ca# 28704). Final DNA elution was in 35 μL sterile DI water. Gel-extracted products were prepared for sequencing by aliquoting 11 μL of each product into two separate 0.5 mL PCR strip tubes and adding 1 μL of forward primer (3 pmol) to one tube and 1 μL of reverse primer (3 pmol) to the other. Products were sent to the University of Calgary, University Core DNA services for sequencing.
Fungal genomic DNA extraction and sequencing: The DNA extraction procedure described here is based on a method previously developed by Dr. Kim Kenward with modifications (K. Kenward, 2007, personal communication). Fresh cultures of fungal tissue and cells were prepared for DNA extraction by washing with sterile DI water to remove media and exposure to -20°C overnight to improve DNA recovery. Five milliliters of fungal extraction buffer (100 mM tris, 10 mM EDTA, 1% v/v SDS, pH 8.0) and 5% v/v of 20% PEG 8000 was added to each sample, vortexed for 20 sec and incubated at 65°C for 1 h. After incubation, each fungal sample were vortexed and transferred to a 15 mL phase lock gel light™ tube (Cat# 2302840). Extraction was performed in an equal volume of phenol : chloroform (1:1), gently inverted 30 times and centrifuged for 5 min at 1500×g. This step was repeated twice. Next, 1 volume of chloroform: isoamyl alcohol (24:1) was added to aqueous phase, gently inverted 30 times and centrifuged as before. The aqueous phase was decanted into new 15 mL centrifuge tubes with 0.1 volume of 3 M sodium acetate and 0.7 volume of cold 100% isopropanol added to each tube followed by mixing. Extract was left at room temperature for at least 5 min and then centrifuged at 14,000×g for 5 min to pellet DNA. After decanting supernatant, DNA pellet was rinsed with 70% ethanol and centrifuged at 14,000×g for 1 min. Ethanol supernatant was decanted and pellet was allowed to dry for 2 h. Up to 400 μL of 1xTE (10 mM tris-HCL, 10 mM EDTA, pH 8.0) at 65°C was used to resuspend each DNA pellet. Lastly, RNase A was added to each tube at a concentration of 0.1 μg/100 μL DNA. Quality and quantity of DNA were evaluated and adjusted by absorbance spectroscopy at 260 and 280 nm (NanoDrop Instruments).
The PCR amplification of a housekeeping gene region, β-tubulin was done using the primer set benA-T1 F (5’-ACC ATG CGT GAG ATT GTA AGT-3’) and benA-T2 R (5’- TAG TGA CCC TTG GCC CAG-3’). This primer set amplified a 600 bp fragment by PCR in a 60 μL volume containing 6 μL 10×reaction buffer, 3 μL MgCl2 (25 mM), 1.2 μL TAQ DNA polymerase recombinant (Cat# 10342020, Invitrogen), 41.1 μL sterile DI water, 1.2 μL dNTP’s (25 μM), 2.4 μL each primer (10 pmol) and 2.4 μL genomic DNA (1-50 ng μL–1). Conditions for PCR amplification were: 9 min at 95°C, followed by 35 cycles of 1 min at 95°C, 1 min at 55°C and 1 min at 72°C and ending with 7 min at 72°C. After PCR amplification, 5 μL of product were separated in a 1.2% agarose gel in 1x tris-acetate-EDTA (TAE) buffer. The ~600 bp band was cut out and gel extracted using the Qiagen QIAquick Gel Extraction Kit (Ca# 28704). Final DNA elution was in 35 μL sterile DI water. Gel-extracted product was prepared for sequencing by aliquoting 10 μL of product into two separate 0.5 mL PCR strip tubes and preparing aliquots of F/R primers (20 μM, enough volume for 5 μL per sample). Products were sent to McGill University (Genome Quebec Innovation Centre) for sequencing.
Sequence analysis: Forward and reverse nucleotide sequences were edited, assembled and aligned using Sequencher 4.8 software to obtain high-quality consensus sequences. Consensus sequence homologies were compared to those referenced in the NCBI database [http://blast.ncbi. nlm.nih.gov/Blast,32 by using nucleotide (nr/nt) BLASTN search (2.2.17, database of August, 2007) using default parameters.
Fulfilling of Koch’s postulates: All media used were Acumedia (Acumedia Manufacturers Inc., Lansing, MI) brand unless otherwise indicated. Fresh produce was purchased from local grocery stores and surface sterilized. The produce was heat-sensitive and therefore, needed to be surface sterilized without heat. Mixtures of hypochlorite and alcohol have been reported to be useful for disinfecting heat-sensitive objects33. A 1:1 solution of 1% bleach and 95% ethanol was prepared in a fume hood and the produce samples were immersed for 30 min and then rinsed 3 times in sterile distilled water. Each bacterial isolate were grown on NA and colonies transferred with a sterile loop to 3 mL of sterile distilled water and thoroughly suspended. Transfer and suspension of the bacterium continued until the turbidity visually matched a 0.5 McFarland standard. For fungal isolates, 0.5 cm2 diameter plugs were aseptically collected from the perimeter of an actively growing colony on PDA. The surface-sterilized produce was then wounded by cutting with a sterile scalpel and inoculated with either 250 μL of bacterial inoculum or one 0.5 cm2 fungal plug. Bacteria and fungi were inoculated to produce from which they were originally isolated. Inoculated tissues were placed separately into humid chambers as previously described. Wounding, inoculation and humid chamber set-up were done inside a biosafety cabinet to avoid contamination or release. Humid chambers were incubated at room temperature for 2-14 days depending on the rate of spoilage. When spoilage and/or microbial growth was present on samples in humid chambers, bacteria and fungi were aseptically excised from plant tissues and plated to NA or PDA. Identification of bacteria was done using colony morphology, cell morphology and Gram’s stain results. Fungi were identified using colony and spore morphologies.
Scanning electron microscopy: All samples were prepared for SEM using chemical fixation. Briefly, colonized plant tissues were excised with a scalpel, placed in a 7 mL scintillation vial containing primary fixative [3% glutaraldehyde (Electron microscopy sciences, USA)+1.6% paraformaldehyde (Electron microscopy sciences, USA) in 0.1 M Na cacodylate buffer pH = 7.5 (Electron microscopy sciences, USA)]. About 2-5 mL of primary fixative or enough to completely immerse all tissues was used. Vials were capped and incubated at room temperature in a fume hood for at least 2 h. Fixed tissues were rinsed 3 times (15 min each) in 0.1 M Na cacodylate buffer (Electron microscopy sciences, USA). After rinsing, 1-3 mL (or just enough to cover plant tissues) of 1% osmium tetroxide (Electron microscopy sciences, USA) were added to each vial. Samples in osmium tetroxide were incubated in a fume hood at room temperature for at least 1 h. Rinsing with buffer was repeated (as above) and samples were dehydrated through a graded ethanol (Fisher scientific, Canada) series with 10 min rinses each in ethanol at concentrations of 35, 50, 70, 85 and 95% and two rinses in absolute ethanol). If samples were stored overnight they were immersed in 35% EtOH and kept at 4°C prior to transfer through the dehydration series. Alternatively, samples were air-dried for 24 h in an un-capped vial in a fume hood. All samples were critical point dried, mounted onto aluminum stubs, sputter coated with gold and scanned using a Philips C-60 ESEM.
RESULTS
Bacteria and fungi isolated from produce and environmental sampling of farms: A total of 26 types of vegetables and 7 types of fruits were collected and tested (Table 1). Additionally, five farms were visited and environmental samples from surfaces and air were collected and tested as well.
| Table 1: | Summary of bacterial and fungal isolates collected in Southern Alberta from 33 types of locally-produced fresh fruit and vegetables and two types of environmental sampling |
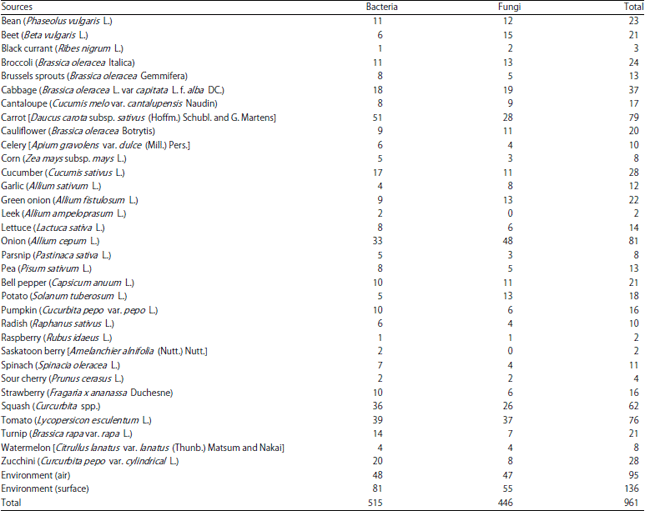 | |
Culturing and isolation of microorganisms from the produce and environment samples yielded a total of 515 bacterial isolates and 446 fungal isolates (Table 1). The greatest numberof isolates came from the environmental sampling of hard surfaces (n = 136) and from air (n = 95) followed by onion (n = 81), carrot (n = 79), tomato (n = 76), squash (n = 62) and cabbage (n = 37). Numbers of isolates collected from the remaining vegetable and fruit commodities are shown in Table 1.
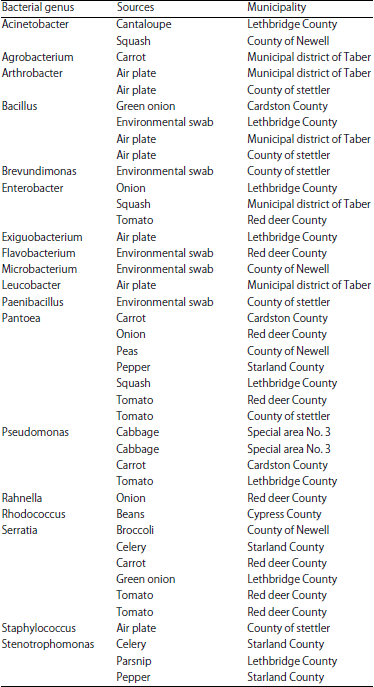
Of the 40 bacterial isolates for which the 16S rRNA region was sequenced, 18 different genera from 14 different sources were revealed (Table 2). The most commonly occurring genus was Pantoea with 7 isolates obtained from 6 sources. Other commonly occurring genera included Serratia, Bacillus and Pseudomonas. Most of the bacterial isolates identified were recovered from tomato. Of the 40 fungal isolates evaluated, the genetic sequence data (β-tubulin and/or EF-1α genes) for only 25 was of adequate quality to confirm identity, revealing 12 genera from 11 different sources (Table 3). Almost all the fungal isolates were from well-known genera and commonly found on plants and food with, such as Penicillium with 8 isolates from 6 sources and Fusarium with 5 isolates from 3 sources. Most of the fungi identified were recovered from onion and tomato followed by carrot and squash.
Fulfilling Koch’s postulates: The isolates of bacteria and fungi identified to species were tested to see if they could cause spoilage when inoculated onto surface-sterilized, wounded produce.
| Table 2: | Bacterial genera recovered from produce or environmental testing of farms from 9 counties in Southern Alberta |
 | |
This testing was done on the produce type from which the microorganisms were originally isolated. The species names for each isolate and their reactions on and recovery from surface-sterilized produce are shown in Table 4 and 5 for bacteria and fungi, respectively. Approximately 19% of the isolates were not recovered or did not cause spoilage, when inoculated onto surface-sterilized wounded produce. Approximately 81% of isolates were either capable of causing spoilage or were recovered in mixed cultures from symptomatic tissues. Approximately half the microorganisms from symptomatic, wound-inoculated tissues were recovered alone, indicating that they were capable of causing spoilage on wounded produce independently.
| Table 3: | Fungal genera recovered from produce or environmental testing of farms |
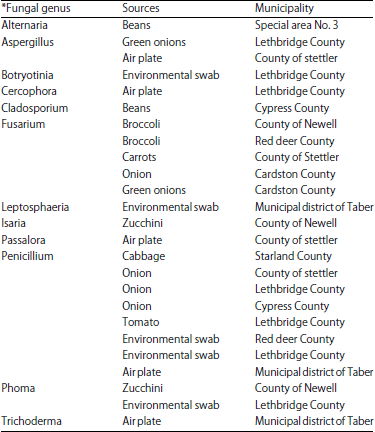 | |
| *Fifteen of the fungal isolates remained unidentified due to insufficient sequence information | |
The remaining isolates were recovered from the symptomatic tissues, but were found in combination with other commonly occurring spoilage fungi such as Fusarium, Penicillium and Rhizopus. These observations indicated that the majority of fungal and bacterial isolates collected from produce in this study were capable of functioning as opportunistic post-harvest spoilage microorganisms either alone or in combination with other spoilage microorganisms.
Scanning electron microscopy: Visualization of bean, beet and carrot tissues with symptoms of spoilage was done via Scanning Electron Microscopy (SEM) to see if there was evidence of microbial biofilms at or near sites of produce spoilage. The SEM images showed the remains of what appeared to be a matrix composed of abundant exopolysaccharide with single-celled and filamentous microorganisms embedded within. These SEM images revealed the presence of mixed-species biofilms associated with produce spoilage symptoms (Fig. 2).
| Table 4: | Bacterial specie’s identities and spoilage potential, when re-inoculated onto fresh produce |
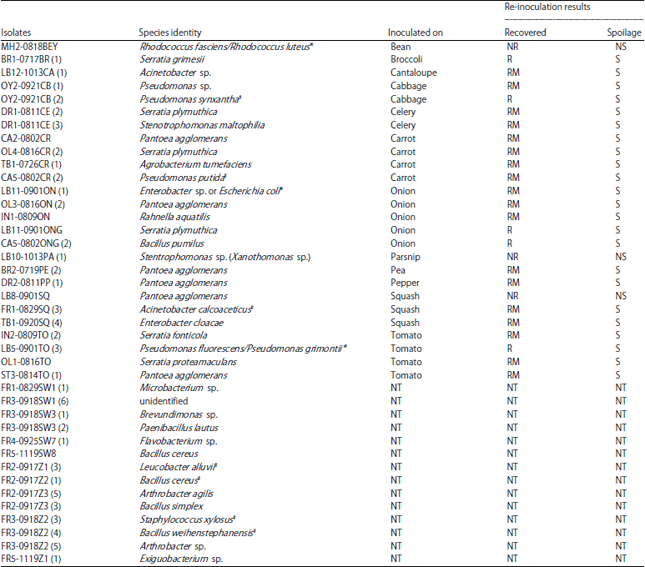 | |
| NR: Not recovered, R: Recovered, RM: Recovered in a mixed population, NS: No spoilage, S: Spoilage and NT: Not tested, *No resolution due to insufficient sequence information and #Closest species epithet | |
Food-borne human pathogens: All 19 environmental surface swabs from the five farms sampled were negative for the presence of Listeria monocytogenes (detection limit = 105 CFU mL–1 after enrichment), which indicated that this food-borne pathogen was not detectable at any of the farms sampled in Southern Alberta.
DISCUSSION
Bacteria and fungi causing spoilage of fresh fruits and vegetables have been characterized by many groups using a variety of techniques 1,2,34. While recognizing the limitations of the culture-based methods used to isolate microbes in this study, there remain some results worthy of discussion. Firstly, the evaluation of communities and populations of microbes on surfaces of fresh produce in Southern Alberta revealed that different sources and types of produce host diverse communities of microorganisms, a result that is consistent with those reported by others1. For example, there was no trend observed towards species that grew on only certain types of produce or at certain locations, but rather it appeared that the species compositions and community diversities were influenced by many factors, which could include location, crop rotation and historical farm operations and inputs (i.e., conventional versus organic), environmental and storage conditions and post-harvest interval.
 | |
| Fig. 2(a-d): | Scanning electron micrograph of vegetable surfaces with post-harvest spoilage, (a-b) Carrot surface at the site of post-harvest decay, rod-shaped and spherically-shaped microorganisms are seen encased in the remnants of an extracellular matrix and (c-d) Bean pod sample with post-harvest decay symptoms. A mixed species community of microorganisms encased in the remnants of an extracellular matrix is associated with the spoilage symptoms, scale bars = 5 µm |
Secondly, many isolates identified to species are commonly known to be associated with fresh produce and to cause spoilage. These included fungal genera, such as Penicillium, Fusarium Aspergillus, Alternaria, Cladosporium and Botrytinia and bacteria from the genera Pseudomonas, Pantoea and Serratia. However, there were also some species that are not generally considered to be food spoilage organisms that were present on produce and showed some ability to cause spoilage. These species included the fungus Isaria farinosus [(Holmsk.) A.H.S. Br. and G. Sm.] and the bacteria Stenotrophomonas maltophilia35, Rahnella aquatilis36 and Acinetobacter calcoaceticus (syn. Micrococcus calcoaceticus). Interestingly, these bacteria all share in common the ability to colonize a wide range of habitats and as having potential as a biological control agents to prevent plant disease or post-harvest spoilage37-39 and yet this study found it to cause spoilage. However, this is not the first time a microorganism has been shown to function as a biological control agent on one fruit and cause spoilage on another. For example, the yeast Pichia fermentans Lodder [anamorph: Candida lambica (Lindner and Genoud) Uden and H.R. Buckley ex S.A. Mey. and Ahearn (1983)] is demonstrated to be an effective biological control for prevention of Monolinia brown rot on apple and yet causes post-harvest spoilage on peach fruit40. Examples such as these indicate that the genetic and metabolic potential of individual microorganisms allows them to be both beneficial and harmful, depending on the host and the environment. Furthermore, our understanding of the microorganisms capable of spoilage and the conditions that are sufficient to produce spoilage is far from complete.
| Table 5: | Fungal specie’s identities and spoilage potential, when re-inoculated onto fresh produce |
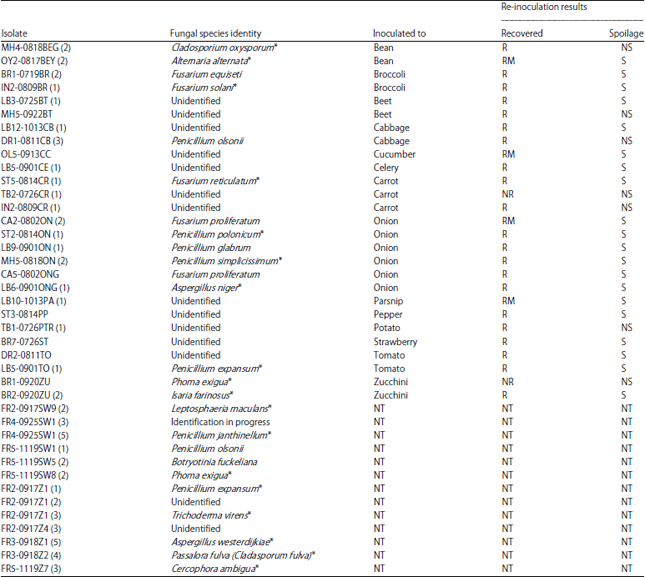 | |
| *Closest species epithet, NR: Not recovered, R: Recovered, RM: Recovered in a mixed population, NS: No spoilage, S: Spoilage and NT: Not tested | |
Another key finding in this report is the evidence of biofilms occurring with spoilage symptoms. It is no surprise to see microorganisms, growing as aggregates or biofilms causing spoilage as this has been documented by others Kumar and Anand41 and Korber et al.42, but this observation has serious implications on management of food spoilage microorganisms. It is well-known that microbial populations within a biofilm matrix are much more tolerant of physical and chemical treatments aimed at eradicating them when compared to solitary, planktonic populations43. Mixed species biofilms causing spoilage is even more troubling because they are even more resistant to disinfectants and sanitizers than mono-species biofilms24 and mixed species biofilms are linked to high-efficiency plasmid transfer, which can accelerate the acquisition and accumulation of genetic sequences that aid survival and minimize efficacy of management tools44. Alternatively, it is also reported that some mixed species biofilms can function in prevention and control of spoilage. For example, some will either inhibit pathogen colonization/growth or enhance their survival, while others can either facilitate internalization of pathogens into the host cells or inhibit the internalization of pathogens into the host cells. The practical relevance of the mixed species biofilms concept is widely accepted and discussed, but study in this area is in its infancy and conflicting results are likely due to differences in methodology, species, conditions, contact surfaces, niches and protocols, etc.24. More controlled studies from the biofilm perspective, using model systems are needed to more clearly draw the line between parasitism, antagonism and commensalism for biofilms containing mixed-species microbial communities.
Finally, this study did not detect Listeria monocytogenes contamination at any of the five farms sampled in Southern Alberta. While it is reassuring that L. monocytogenes was not found this small sampling of production surfaces does not allow any general conclusions regarding food-borne enteropathogenic bacteria on produce in Southern Alberta. A more thorough investigation of food-borne pathogens on fresh produce in Alberta was conducted by Bohaychuk et al.29. Their study looked for generic Escherichi coli, Salmonella spp., E. coli O157:H7 and Campylobacter spp. on lettuce, spinach, green onion, carrot, tomato and strawberry. Although their samples were collected at different locations of the province spatially (central and Northern Alberta), their collections were temporally concurrent with those presented in this report. Bohaychuk et al.29 used E. coli as an indicator of fecal contamination and found that it was detectable in 8.2% of all samples. However, no food-borne bacterial pathogens were detectable on any of the fresh produce sampled.
CONCLUSION
The results given in this report indicate that spoilage of fresh produce in Alberta is due to diverse, mixed-species communities of microorganisms capable of forming or becoming incorporated within, biofilms. Additionally, we see evidence that some microorganisms primarily growing commensally on plants or in soil as epiphytes and/or saprophytes, may also have the potential to opportunistically exploit harvested produce as a food source and cause spoilage.
ACKNOWLEDGMENTS
This study was supported by the Canadian Food Safety and Quality Program, Food Safety Initiative Funding, under Grant# 43885. The authors thank the staff of the Food Microbiology Laboratory of the Agri-Food Laboratories Branch, Alberta Agriculture and Rural Development, Edmonton, AB for Listeria monocytogenes screening of environmental sponges. We also thank DNA sequencing laboratories at University of Calgary (University Core DNA Services, Calgary, AB) and McGill University (Genome Quebec Innovation Centre, Montreal QC) for molecular analyses. Assistance with scanning electron microscopy from Nick Allan and Liz Middlemiss (Innovotech Inc., Edmonton, AB) is gratefully acknowledged.
REFERENCES
- Critzer, F.J. and M.P. Doyle, 2010. Microbial ecology of foodborne pathogens associated with produce. Curr. Opin. Biotechnol., 21: 125-130.
CrossRefDirect Link - Leff, J.W. and N. Fierer, 2013. Bacterial communities associated with the surfaces of fresh fruits and vegetables. PloS One, Vol. 8.
CrossRefDirect Link - Liao, C.H. and W.F. Fett, 2001. Analysis of native microflora and selection of strains antagonistic to human pathogens on fresh produce. J. Food Protect., 64: 1110-1115.
Direct Link - Round, J.L. and S.K. Mazmanian, 2009. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol., 9: 313-323.
CrossRefDirect Link - Lee, Y.K. and S.K. Mazmanian, 2010. Has the microbiota played a critical role in the evolution of the adaptive immune system? Science, 330: 1768-1773.
CrossRefDirect Link - Tournas, V.H., 2005. Spoilage of vegetable crops by bacteria and fungi and related health hazards. Crit. Rev. Microbiol., 31: 33-44.
CrossRefPubMedDirect Link - Hanski, I., L. von Hertzen, N. Fyhrquist, K. Koskinen and K. Torppa et al., 2012. Environmental biodiversity, human microbiota and allergy are interrelated. Proc. Natl. Acad. Sci. USA., 109: 8334-8339.
CrossRefDirect Link - King, A.D., J.A. Magnuson, T. Torok and N. Goodman, 1991. Microbial flora and storage quality of partially processed lettuce. J. Food Sci., 56: 459-461.
CrossRefDirect Link - Nguyen-The, C. and F. Carlin, 1994. The microbiology of minimally processed fresh fruits and vegetables. Crit. Rev. Food Sci. Nut., 34: 371-401.
CrossRefPubMedDirect Link - Beuchat, L.R., 1996. Pathogenic microorganisms associated with fresh produce. J. Food Protect., 59: 204-216.
Direct Link - Badosa, E., R. Trias, D. Pares, M. Pla and E. Montesinos, 2008. Microbiological quality of fresh fruit and vegetable products in Catalonia (Spain) using normalised plate-counting methods and real time polymerase chain reaction (QPCR). J. Sci. Food Agric., 88: 605-611.
CrossRefDirect Link - Ponce, A.G., M.V. Aguero, S.I. Roura, C.E. Del Valle and M.R. Moreira, 2008. Dynamics of indigenous microbial populations of butter head lettuce grown in mulch and on bare soil. J. Food Sci., 73: M257-M263.
CrossRefDirect Link - Oliveira, M., J. Usall, I. Vinas, M. Anguera, F. Gatius and M. Abadias, 2010. Microbiological quality of fresh lettuce from organic and conventional production. Food Microbiol., 27: 679-684.
CrossRefDirect Link - Rastogi, G., A. Sbodio, J.J. Tech, T.V. Suslow, G.L. Coaker and J.H. Leveau, 2012. Leaf microbiota in an agroecosystem: Spatiotemporal variation in bacterial community composition on field-grown lettuce. ISME J., 6: 1812-1822.
CrossRefDirect Link - Liu, N.T., A.M. Lefcourt, X. Nou, D.R. Shelton, G. Zhang and Y.M. Lo, 2013. Native microflora in fresh-cut produce processing plants and their potentials for biofilm formation. J. Food Protect., 76: 827-832.
CrossRefDirect Link - Shapaval, V., J. Schmitt, T. Moretro, H.P. Suso and I. Skaar et al., 2013. Characterization of food spoilage fungi by FTIR spectroscopy. J. Applied Microbiol., 114: 788-796.
CrossRefDirect Link - Liu, N.T., X. Nou, A.M. Lefcourt, D.R. Shelton and Y.M. Lo, 2014. Dual-species biofilm formation by Escherichia coli O157:H7 and environmental bacteria isolated from fresh-cut processing facilities. Int. J. Food Microbiol., 171: 15-20.
CrossRefDirect Link - Donlan, R.M., 2002. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis., 8: 881-890.
CrossRefPubMedDirect Link - Hall-Stoodley, L., J.W. Costerton and P. Stoodley, 2004. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol., 2: 95-108.
CrossRefDirect Link - Harding, M.W., L.L.R. Marques, R.J. Howard and M.E. Olson, 2009. Can filamentous fungi form biofilms? Trends Microbiol., 17: 475-480.
CrossRefDirect Link - Fett, W.F. and P.H. Cooke, 2003. Scanning electron microscopy of native biofilms on mung bean sprouts. Can. J. Microbiol., 49: 45-50.
CrossRefDirect Link - Heaton, J.C. and K. Jones, 2008. Microbial contamination of fruit and vegetables and the behaviour of enteropathogens in the phyllosphere: A review. J. Applied Microbiol., 104: 613-626.
CrossRefDirect Link - Berger, C.N., S.V. Sodha, R.K. Shaw, P.M. Griffin, D. Pink, P. Hand and G. Frankel, 2010. Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ. Microbiol., 12: 2385-2397.
CrossRefPubMedDirect Link - Jahid, I.K. and S.D. Ha, 2014. The paradox of mixed-species biofilms in the context of food safety. Comprehen. Rev. Food Sci. Food Saf., 13: 990-1011.
CrossRefDirect Link - Harding, M.W., L.L. Marques, R.J. Howard and M.E. Olson, 2010. Biofilm morphologies of plant pathogenic fungi. Am. J. Plant Sci. Biotechnol., 4: 43-47.
Direct Link - Pirie, J.H.H., 1940. Listeria: Change of name for a genus bacteria. Nature, 145: 264-264.
CrossRefDirect Link - Murray, E.G.D., R.A. Webb and M.B.R. Swann, 1926. A disease of rabbits characterised by a large mononuclear leucocytosis, caused by a hitherto undescribed bacillus Bacterium monocytogenes (n.sp.). J. Pathol. Bacteriol., 29: 407-439.
CrossRefDirect Link - Bohaychuk, V.M., R.W. Bradbury, R. Dimock, M. Fehr and G.E. Gensler et al., 2009. A microbiological survey of selected Alberta-grown fresh produce from farmers' markets in Alberta, Canada. J. Food Protect., 72: 415-420.
Direct Link - Jagoueix, S., M.J. Bove and M. Garnier, 1994. The phloem-limited bacterium of greening disease of citrus is a member of the α subdivision of the proteobacteria. Int. J. Syst. Evol. Microbiol., 44: 379-386.
CrossRefPubMedDirect Link - Kaneshiro, W.S., C.Y. Mizumoto and A.M. Alvarez, 2006. Differentiation of Clavibacter michiganensis subsp. michiganensis from seed-borne saprophytes using ELISA, Biolog and 16S rDNA sequencing. Eur. J. Plant Pathol., 116: 45-56.
CrossRefDirect Link - Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Coates, D. and J.E. Death, 1978. Sporicidal activity of mixtures of alcohol and hypochlorite. J. Clin. Pathol., 31: 148-152.
CrossRefDirect Link - Yashiro, E., R.N. Spear and P.S. McManus, 2011. Culture-dependent and culture-independent assessment of bacteria in the apple phyllosphere. J. Applied Microbiol., 110: 1284-1296.
CrossRefDirect Link - Palleroni, N.J. and J.F. Bradbury, 1993. Stenotrophomonas, a new bacterial genus for Xanthomonas maltophilia (Hugh 1980) Swings et al. 1983. Int. J. Syst. Evol. Microbiol., 43: 606-609.
CrossRefPubMedDirect Link - Izard, D., F. Gavini, P.A. Trinel and H. Leclere, 1979. [Rahnella aquatilis, a new member of the enterobacteriaceae]. Annales Microbiologie, 130: 163-177.
PubMedDirect Link - Calvo, J., V. Calvente, M.E. de Orellano, D. Benuzzi and M.I.S. de Tosetti, 2007. Biological control of postharvest spoilage caused by Penicillium expansum and Botrytis cinerea in apple by using the bacterium Rahnella aquatilis. Int. J. Food Microbiol., 113: 251-257.
CrossRefDirect Link - Zimmermann, G., 2008. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): Biology, ecology and use in biological control. Biocontrol Sci. Technol., 18: 865-901.
CrossRefDirect Link - Hayward, A.C., N. Fegan, M. Fegan and G.R. Stirling, 2010. Stenotrophomonas and lysobacter: Ubiquitous plant-associated γ-proteobacteria of developing significance in applied microbiology. J. Applied Microbiol., 108: 756-770.
CrossRefDirect Link - Giobbe, S., S. Marceddu, B. Scherm, G. Zara, V.L. Mazzarello, M. Budroni and Q. Migheli, 2007. The strange case of a biofilm-forming strain of Pichia fermentans, which controls Monilinia brown rot on apple but is pathogenic on peach fruit. FEMS Yeast Res., 7: 1389-1398.
CrossRefDirect Link - Kumar, C.G. and S.K. Anand, 1998. Significance of microbial biofilms in food industry: A review. Int. J. Food Microbiol., 42: 9-27.
CrossRefDirect Link - Simoes, L.C., M. Simoes and M.J. Vieira, 2010. Influence of the diversity of bacterial isolates from drinking water on resistance of biofilms to disinfection. Applied Environ. Microbiol., 76: 6673-6679.
CrossRefDirect Link - Van Meervenne, E., R. De Weirdt, E. Van Coillie, F. Devlieghere, L. Herman and N. Boon, 2014. Biofilm models for the food industry: Hot spots for plasmid transfer? Pathogens Dis., 70: 332-338.
CrossRefDirect Link